Research Article
An Annealing Control Primer (ACP) System Used for the Isolation and Identification of Copper-Induced Genes in Alfalfa Leaves
Ki-Won Lee, Sang-Hoon Lee, Ki-Yong Kim, Hee Chung Ji, Hyung Soo Park, Tae Young Hwang, Gi Jun Choi and Md. Atikur Rahman*
Grassland & Forages Division, National Institute of Animal Science, Rural Development Administration, Cheonan 31000, Republic of Korea
ABSTRACT
Copper (Cu) is a necessary microelement for plants. However, high concentrations of Cu are toxic to plants that change the regulation of several stress-induced proteins. In this study, an annealing control primer (ACP) based approach was used to identify differentially expressed Cu-induced genes in alfalfa leaves. Two-week-old alfalfa plants (Medicago sativa L.) were exposed to Cu for 6 h. Total RNAs were isolated from treated and control leaves followed by ACP-based PCR technique. Using GeneFishing ACPs, we obtained several genes those expression levels were induced by Cu. Finally, we identified several genes including UDP-glucuronic acid decarboxylase, transmembrane protein, small heat shock protein, C-type cytochrome biogenesis protein, mitochondrial 2-oxoglutarate, and trans-2,3-enoyl-CoA reductase in alfalfa leaves. These identified genes have putative functions in cellular processes such as cell wall structural rearrangements, transduction, stress tolerance, heme transport, carbon and nitrogen assimilation, and lipid biosynthesis. Response of Cu-induced genes and their identification in alfalfa would be useful for molecular breeding to improve alfalfa with tolerance to heavy metals.
(Key words : Copper, Gene, Toxicity, Alfalfa)
. INTRODUCTION
Heavy metal contamination is one of the most serious environmental thread faces by plants and humans. Precisely in case of copper (Cu), it occurs serious problems due to frequently use in industrial process and agricultural practice (Fernandez and Henriques, 1991). At the same time, presence of high concentration of Cu can be toxic to the plants (Chen et al., 2015). Plants are sensitive to Cu toxicity. It has been reported that the optimal level of Cu in plant tissues is 5-20 µg g
1(Fernandez and Henriques, 1991). However, the level of Cu in contaminated soil in South Asian countries is higher than acceptance level.
Excessive level of Cu in plant impairs a series of physiological and biochemical processes including leaf chlorosis, growth retardation, and metabolic disturbance. Cu interferences with enzyme and fatty acid metabolism, inhibits respiration and N2 fixation. High amount of Cu significantly reduces vegetative growth and induces the
indication of senescence in plants (Fernandez and Henriques, 1991).
Cu toxicity has most important implication for blocking of photosynthetic electron transport, and generating of free radicals. Reactive oxygen species (ROS) is reactive molecule that induced by several heavy metal stress including excess Cu toxicity (Boojar and Goodarzi et al., 2007). However, the high level of ROS in cell induces oxidative stress.
Plants develop a verity of defense mechanisms against oxidative damages by antioxidant enzymes / proteins. The gene or protein identification under multiple environmental stresses is the fundamental steps for understanding the molecular mechanisms in plants. Recently several abiotic stress induced proteins or gene identification techniques have been revealed in forage legume and grass plants including alfalfa (Rahman et al., 2016; Rahman et al., 2015;
Lee et al., 2015). An annealing control primer (ACP) technique is frequently used for identification of differentially expressed genes (DEGs) in plants and animals (Lee et al.,
* Corresponding author : Md. Atikur Rahman, Grassland & Forages Division, National Institute of Animal Science, Rural Development Administration, Cheonan 31000, Republic of Korea. Tel: +82-41-580-6767, Email: [email protected]
2015; Kim et al., 2010; Choi et al., 2007). In ACP-based technique, emphasizes on oligonuleotide primers which anneal with specificity to the desired template, whereas allowing only the target sequence to be amplified.
GeneFishing
TMDEG technique has been used for the detection of DEGs in multiple samples.
Alfalfa is one of the world’s leading forage legumes. It has been reported that alfalfa growth was inhibited by Cu toxicity (Ippolito et al., 2011). Therefore, it is imperative to find out Cu tolerance strategies along with promising genes / proteins by modern biotechnological approaches. Gene or protein identification in response to multiple stresses is the suitable way for understanding of molecular mechanism / to develop stress tolerant crops. To investigate the Cu-stress responsive genes in alfalfa leaves and their possible function in response to Cu stress, an ACP- based approach has been applied. The main focus of this study, to isolate and identify genes / proteins which are highly induced by Cu stress, that can provide new insight to develop heavy metal stress tolerant crop.
. MATERIALS AND METHODS 1. Plant material and copper treatment
Alfalfa seeds were planted in plastic pots containing commercial potting mix (Nursery Medium
®, Biomedia, Korea) and grown in growth chamber maintained at temperature of 25°C, 55~65% humidity with a light intensity of 400 mol m
2s
1and 14-h photoperiod. Two-week-old alfalfa seedlings were irrigated with cupric sulfate (CuSO
4.5H
2O, 500 µM) solutions up to 6 hours. After completing of treatment, plant leaves were harvested then quickly frozen in liquid nitrogen then stored at 80°C until further analysis.
2. Isolation of RNA, first strand cDNA synthesis and ACP-based GeneFishing
Total RNAs were isolated from treated and control leaf tissues of alfalfa using Plant RNeasy mini kit (Qiagen, USA), subsequently used it for the reverse-transcriptase- catalyzed first strand cDNA synthesis. The reverse transcription
reaction was performed according to Lee et al. (2009). In RT reaction was maintained up to 1.5 h at 42ºC in a final reaction volume of 20 µl, whereas the mixture containing 3 µg (7.5 µl) of the purified total RNA; 4 µl of 5 × reaction buffer (Promega, USA); 5 µl of dNTPs (for every 2 mM); 2 µl of 10 µM dT-ACP1 (5´-CTGTGAATGCTGCGACTACGA TIIIIIT(18)-3´); 0.5 µl of RNas in RNase Inhibitor (40 U/µl;
Promega); and 1 µl of moloney murine leukemia virus (MMLV) reverse transcriptase (200 U/µl; Promega). After synthesis, first-strand cDNAs were diluted by adding of 80 µl of purified water, finally prepared for GeneFishing
TMPCR.
Differentially expressed genes (DEGs) were screened by ACP-based PCR method using the GeneFishing
TMDEG kits (Seegene, Korea). DEGs were identified by an ACP-based PCR method as described by Lee et al. (2009). Briefly, second-strand cDNA synthesis was carried out at 50°C (mild stringency) during one cycle of first stage PCR. The final reaction volume of 49.5 l including 3~5 μ μ l (each 50 ng) of diluted first-strand DNA cDNA, 5 l of 10x PCR μ buffer plus Mg (Roche Applied Science, Germany), 5 µL of dNTP (each 2 mM), 1 µL of oligo(dT)15, oligo(dT)15 tail, or oligo(dT)15 ACP (10 µM), 1 µL of 10 µM arbitrary primer. The tube containing reaction mixture was kept at 94°C whereas 0.5 l of Taq DNA polymerase was added to μ the reaction mixture (5 U/ l, Roche Applied Science). The μ PCR steps for second-strand synthesis was maintained for one cycle at 94°C for 1 min, fol lowed by 50°C for 3 min, and 72°C for 1 min. After completion of second-strand DNA synthesis, 40 cycles were performed. Each cycle involved denaturation at 94°C for 40 sec, annealing at 65°C for 40 sec, extension at 72°C for 40 sec, and a final extension at 72°C to complete the reaction. The amplified PCR products were separated in 2% agarose gel and stained with RedSafe (Nucleic acid staining solution; iNtRON Biotechnology, Korea).
3. Gene cloning sequencing and bioinformatics analysis
The obtained DEGs were extracted from the gel by using
the GENCLEAN® II Kit (Q-BIO gene, USA) and directly
cloned into a TOPO TA cloning vector (Invitrogen, USA)
according to the manufacturer’s instructions. The cloned
plasmids were sequenced with an ABI PRISM 3100 Genetic Analyzer (Applied Biosystems, USA) using the M13 forward primer (5’-CGCCAGGGTTTTCCCAGTCACGA-3’) and M13 reverse primer (5’-AGCGGATAACAATTTCACACAGGA-3’) containing a total of 23 and 24 oligo- nucleotides, respectively. Sequencing data were used for bioinformatics analysis. We confirmed cDNA sequence in GenBank database through the BlastX program of NCBI (http://www.ncbi.nlm.nih.gov/BLAST/).
. RESULTS AND DISCUSSION
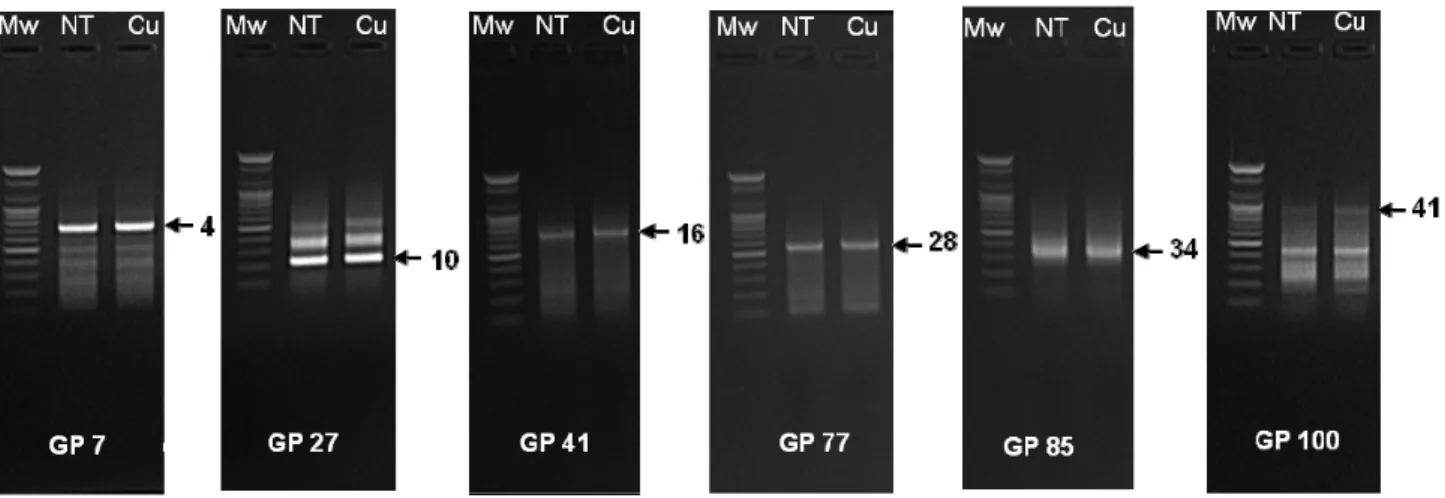
In order to identify Cu-responsive DEGs, ACP-based GeneFishing PCR approach was performed. By using different gene fishing primer we identified several Cu- induced genes including UDP-glucuronic acid decarboxylase (DEG 4), transmembrane protein (DEG 10), small heat shock protein (DEG 16), C-type cytochrome biogenesis protein (DEG 28), mitochondrial 2-oxoglutarate (DEG 34) and trans-2,3-enoyl-CoA reductase (DEG 41) in alfalfa leaf (Table 1, Fig. 1). According to their biological functions,
these identified genes are involved in several pathways and/
or processes including cell wall structural rearrangements, transduction, stress tolerance, heme transport, carbon and nitrogen assimilation, and lipid biosynthesis.
The components of plant cell wall like pectic polymers and hemicelluloses, which are predominantly derived from the precursor UDP-glucuronic acid (UDP-GlcA). UDP-GlcA produces from UDP- -D-glucose. Moreover, the UDP- α glucuronic acid decarboxylase (UGD) catalyzes the conversion of UDP-GlcA to UDP-xylose via a decarboxylation reaction, where UDP xylose is required for the synthesis of diverse plant cell-wall polysaccharides and xyloglucan (Pattathil et al., 2005). However, cell well component and rearrangement are important for stress tolerant mechanism whereas UGD involved during stress imposition. In this study, we found the induction of UDG (DEG 4) during Cu stress, indicating UDG involve in the alteration of cell wall components.
Previously it found to be down-regulated of UGD in barley response to salt stress (Mostek et al., 2015).
The signal transduction pathway triggers a series of molecular responses due to stress perception in plant cell.
Table 1. Copper (Cu)-stress induced differentially expressed genes in alfalfa leaves identified by ACP-based PCR approach. The DEGs sequences and their corresponding genes were searched and confirmed by BLASTP (http://www.ncbi.nlm.nih.gov/BLAST/).
No. Accession No. Name of gene (Blast search) Total score E-value Identity Length (bp)
DEG 4 XP_003591521.1 UDP-glucuronic acid decarboxylase
[Medicago truncatula] 347 3.00E-114 100% 800
DEG 10 XP_003613720.1 Transmembrane protein, putative
[Medicago truncatula] 44.3 5.00E-03 63% 380
DEG 16 KEH17633.1 Small heat shock protein
[Medicago truncatula] 340 4.00E-114 98% 908
DEG 28 KEH19586.1 C-type cytochrome biogenesis protein
[Medicago truncatula] 215 6.00E-65 99% 525
DEG 34 AFK44647.1 Mitochondrial 2-oxoglutarate/malate
carrier protein [Medicago truncatula] 166 5.00E-47 99% 444
DEG 41 XP_003617070.1 Trans-2,3-enoyl-CoA reductase
[Medicago truncatula] 322 8.00E-106 99% 809
Under Cu-stress, we have been found the transmembrane protein (DEG10) that was induced in alfalfa leaf. However, transmembrane proteins can activated response to high osmolarity that has been reported previously in yeast (Jan-Peter Nap, 2003), suggesting that it (DEP10) is a signal responsive protein as well as oxidative stress may induce during high concentration of Cu in alfalfa plants.
Small heat shock proteins (sHsps) are belonging to the chaperone family which is induced ubiquitously in prokaryotic and eukaryotic system response to heat stress (Sun et al., 2002). Though it is well known sHsps are involved in stabilizing and preventing aggregation of proteins under stress subsequently participate in abiotic stress tolerance (Wang et al., 2004), but mechanisms of cell protection by sHsps are poorly documented. sHsps are largely elucidated as molecular chaperone activity in vitro (Lee et al., 1997). However, we found sHsp was induced response to Cu-stress indicating a correlation of stress tolerance in alfalfa plant. C-type cytochrome biogenesis protein (DEG 28), involved in heme biogesis pathway (Lezhneva et al., 2008). Heme has significant regulatory function in cell biology, as a co-factor heme plays pivotal role in fundamental biological processes including oxidative metabolism, oxygen storage and transport, and signal transduction. However, specific role of C-type cytochrome biogenesis protein during heavy metal stresses are mostly unknown.
The protein 2-oxoglutarate (2OG) belongs to a sub-family of oxidoreductases, proteins involved in plant growth and development. Previously it has been documented as salt responsive protein in model plant Brachypodium distachyon (Kim et al., 2012). However, 2OG can be consumed in glutamine synthetase-glutamate synthase cycle during carbon- deficit conditions response to salinity (Hodges et al., 2003).
In our study we, found mitochondrial 2-oxoglutarate (2OG ; DEG34) to be induced indicating Cu-toxicity not only involved in growth retardation of plant but also an indication of carbon deficiency during stress.
Lipids are important components of biological membrane regulate the physiological functions of the membrane. It plays as signaling molecules that can sense the extracellular condition. Lipid-mediated signaling induces in response to multiple stresses including salinity, extreme temperature, and drought. To mitigate particular stresses, accumulation of unique lipids even in trace quantities play essential under stressed conditions in plants (Okazaki et al., 2014).
Unfortunately metal ions interfere with the biogenesis of several classes of lipids. However, under Cu-stress trans- 2,3-enoyl-CoA reductase (DEG 41) was found to be induced that involved in lipid synthesis. This result suggests that several alterations may be occurred during Cu-toxicity in alfalfa plants. This enzyme would be considered as lipid-mediated signaling or defense molecule against Cu stress that can contribute to plant survival.
Fig. 1. Agarose gel image shows the bands of differentially expressed genes (DEGs) in alfalfa leaves. Arrows
indicating copper (Cu) induced DEGs. Mw, molecular weight size marker; NT, non-treatment; Cu, copper
treatment; GP, gene fishing primer.
. CONCLUSION
Annealing control primer based PCR technique is very simple and convenient approach that was applied for the identification of differentially express genes in alfalfa leaves response to Cu-stress. We have identified several genes including UDP-glucuronic acid decarboxylase, transmembrane protein, small heat shock protein, C-type cytochrome biogenesis protein, mitochondrial 2-oxoglutarate and trans- 2,3-enoyl-CoA reductase in alfalfa leaf. These identified genes are involved in several pathways and/or processes including cell wall structural rearrangements, transduction, stress tolerance, heme transport, carbon and nitrogen assimilation, and lipid biosynthesis. Understanding of the mechanisms of Cu-stress tolerance and Cu-responsive genes in alfalfa would be useful for molecular breeding of heavy metal tolerant crop production.
. ACKNOWLEDGEMENTS
This work was supported by the “Cooperative Research Program for Agriculture Science & Technology Development (Project No. PJ00859904)” and Postdoctoral Fellowship Program of 2016, National Institute of Animal Science, Rural Development Administration, Republic of Korea.
. REFERENCES
Boojar, M.M.A. and Goodarzi, F. 2007. The copper tolerance strategies and the role of antioxidative enzymes in three plant species grown on copper mine. Chemosphere. 67:2138-2147.
Chen, J., Shafi, M., Li, S., Wang, Y., Wu, J., Ye, Z., Peng, D., Yan, W. and Liu, D. 2015. Copper induced oxidative stresses, antioxidant responses and phytoremediation potential of Moso bamboo (Phyllostachys pubescens). Scientific Reports. 5:13554.
Choi, Y.W., Kim, Y.W., Bae, S.M., Kwak, S.Y., Chun, H.J., Tong, S.Y., Lee, H.N., Shin, J.C., Kim, K.T., Kim, Y.J. and Ahn, W.S.
2007. Identification of differentially expressed genes using annealing control primer-based genefishing in human squamous cell cervical carcinoma. Clinical Oncology. 19:308-318.
Fernandes, J.C. and Henriques, F.S. 1991. Biochemical, physiological, and structural effects of excess copper in plants. The Botanical Review. 57:246-273.
Hodges, M., Flesch, V., Gálvez, S. and Bismuth, E. 2003. Higher
plant NADP dependent isocitrate dehydrogenases, ammonium assimilation and NADPH production. Plant Physiology and Biochemistry. 41:577-585.
Ippolito, J.A., Ducey T.F. and Tarkalson, D.D. 2011. Interactive effects of copper on alfalfa growth, soil copper, and soil bacteria. World Journal of Agricultural Sciences. 3(2):138-148.
Jan-Peter Nap, 2003. Genomics for biosafety and plant biotechnology.
Proceeding of the NATO Advanced Research Workshop, 15-19 October, Bansko, Bulgaria.
Kim, D.Y., Hong, M.J., Jang, J.H. and Seo, Y.W. 2012. cDNA-AFLP analysis reveals differential gene expression in response to salt stress in Brachypodium distachyon. Genes & Genomics. 34:
475-484.
Kim, Y.-S., Do.H.J., Bae, S., Bae, D.-H. and Ahn, S.W. 2010.
Identification of differentially expressed genes using an annealing control primer system in stage III serous ovarian carcinoma.
BMC Cancer. 10:1-14.
Lee, G.J., Roseman, A.M., Saibil, H.R. and Vierling, E. 1997. A small heat shock protein stably binds heat-denatured model substrates and can maintain a substrate in a folding-competent state. The EMBO Journal. 16:659-671.
Lee, S.-H., Lee, K.-W., Kim, K.-Y., Choi, G.J., Yoon, S.H., Ji, H.C., Seo, S., Lim, Y.C. and Ahsan, N. 2009. Identification of salt-stress induced differentially expressed genes in barley leaves using the annealingcontrol-primer-based GeneFishing technique.
African Journal of Biotechnology. 8(7):1326-1331.
Lee, S.-H., Choi, G.J., Kim, K.-Y., Ji, H.C., Park, H.S., Lee, D.-G.
and Lee, K.-W. 2015. Identification of temperature stress-induced differentially expressed genes of tall fescue leaves. Research Journal of Biotechnology. 10(3):99-101.
Lezhneva, L., Kuras, R., Ephritikhine, G. and de Vitry, C. 2008. A novel pathway of cytochrome c biogenesis is involved in the assembly of the cytochrome b(6)f complex in Arabidopsis chloroplasts. The Journal of Biological Chemistry. 283:24608- 24616.
Mostek, A., Börner, A., Badowiec, A. and Weidner, S. 2015.
Alterations in root proteome of salt-sensitive and tolerant barley lines under salt stress conditions. Journal of Plant Physiology.
174:166-176.
Okazaki, Y. and Saito, K. 2014. Roles of lipids as signaling molecules and mitigators during stress response in plants. The Plant Journal. 79:584-596.
Pattathil, S., Harper, A.D. and Bar-Peled, M. 2005. Biosynthesis of UDP-xylose: characterization of membrane-bound AtUxs2. Planta.
221(4): 538-548.
Rahman, M.A., Alam, I., Kim, Y.-G., Ahn, N.-Y., Heo, S.-H., Lee,
D.-G., Liu, G. and Lee, B.-H. 2015. Screening for salt-responsive proteins in two contrasting alfalfa cultivars using a comparative proteome approach. Plant Physiology and Biochemistry. 89:112- 122.
Rahman, M.A., Kim, Y.-G, Alam, I., Liu, G., Lee, H., Lee, J.J. and Lee B.-H. 2016. Proteome analysis of alfalfa roots in response to water deficit stress. Journal of Integrative Agriculture. 15:
1275-1285.
Sun, W., Van Montagu, M. and Verbruggen, N. 2002. Small heat shock proteins and stress tolerance in plants. Biochimica et Biophysica Acta. 1577:1-9.
Wang, W., Vinocur, B., Shoseyov, O. and Altman, A. 2004. Role of plant heat-shock proteins and molecular chaperones in the abiotic stress response. Trends Plant Science. 9(5):244-252.
(Received August 15, 2016 / Revised September 5, 2016 / Accepted September 5, 2016)