Vol. 13, No. 3, September, 2006
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
<접수일:2006년 5월 9일, 심사통과일:2006년 6월 27일>
※통신저자:류 완 희
전주시 덕진구 금암동 산 2-20번지 전북대학교 의과대학 내과학교실
Tel:063) 250-1672, Fax:063) 254-1609, E-mail:[email protected] 이 논문은 한국과학재단의 해외 Post-doc. 연수지원에 의하여 연구되었음.
전신홍반루푸스 환자에서 IFN-α에 의한 CD4+ T 림프구의 ERK Phosphorylation의 감소
전북대학교 의과대학 내과학교실, 임상의학연구소
류 완 희
= Abstract =
IFN-α Downregulates ERK Phosphorylation of CD4+ T Lymphocytes in Systemic Lupus Erythematosus
Wan-Hee Yoo, M.D.
Department of Internal Medicine, Chonbuk National University Medical School, Research Institute of Clinical Medicine, Jeonju, Korea
Objective: CD4+ T cells from patients with systemic lupus erythematosus (SLE) display aberrant TCR signaling and IFN-α plays critical roles in the pathogenesis of SLE; however, the effects of IFN-α on disease-associated TCR signaling defects remain unknown. This study inves- tigated the ERK phosphorylation during TCR triggering and the effects of IFN-α on ERK signaling in CD4+ T cells.
Methods: CD4+ T lymphocytes were sorted from PBMC using magnetic beads in patients with SLE who met the 1982 revised ACR criteria for SLE and age-matched healthy controls.
The phosphorylation of ERK 1/2 was analyzed by flow cytometry and mean fluorescent intensity was measured to define the degree of phosphorylation of ERK. In some experiments, anti-CD3 stimulation was performed after preincubation with patient or control serum, diluted in tissue culture media, with or without addition of an anti-IFN-α antibody. The serum level of IFN-α was measured by ELISA.
Results: ERK-1/2 phosphorylation was decreased in CD4+ T cells of lupus patients than healthy controls and associated with disease activity. Pre-incubation of control CD4+ T cells with allogeneic lupus plasma decreased ERK-1/2 phosphorylation more than allogeneic control and RA plasma and this was reversed by anti-IFN-α Ab. Accordingly, ERK-1/2 phosphorylation
서 론
전신홍반루푸스(systemic lupus erythematosus, 루푸 스)는 chromatin, ribonucleoproteins 등 세포 내 성분 에 대한 자가항체의 생성과 T 림프구의 비정상적 반응을 특징으로 하는 전신성, 자가면역 질환이다 (1,2). 루푸스에서는 T 림프구 수용체(T cell receptor, TCR)의 매개에 의한 신호전달 체계의 이상, B 림프 구의 과대반응, apoptosis의 증가, 사이토카인 생성의 변화와 면역관용의 파괴 등 (3,4) 다양한 세포의 이 상에 의해서 발생하는 것으로 알려져 있다. 루푸스 쥐 모델과 루푸스 환자의 T 림프구에는 내인적 결 함으로 인하여 비정상적인 기능의 활성화를 갖고 있 다 (5,6). 즉, 루푸스 T 림프구는 TCR signaling (7)과, anergy avoidance의 이상 (8), CD40L (CD154)의 이상 발현 (9) 등이 in vitro 실험에서 밝혀졌다.
ERK pathway는 여러 가지 세포의 자극에 대한 세 포막으로부터 핵으로의 필수적인 신호전달체계이며 (10), 세포의 활성화의 조절에 기본적인 물질이다 (11). 루푸스에서는 여러 가지 TCR signaling의 결함 이 알려져 있으며, 활동성 루푸스 환자의 T 림프구 에서 ERK pathway의 결함이 병인에 중요한 역할을 할 수 있는 것으로 보고 있다. 정상대조군과 류마티 스 관절염 환자에 비해서 루푸스 환자의 T 림프구 는 정상적인 양의 ERK 단백질에도 불구하고 ERK- 1/2의 인산화가 감소해 있으며 (12), 이는 루푸스 T 림프구에서 DNA hypomethylation을 통하여 lymp- hocyte function-associate antigen-1 (LFA-1), CD70와 같 은 유전자의 발현을 증가시킨다 (13). 이러한 ERK pathway의 결함은 유전자 내인적 결함이나 유전자 발현의 조절 장애 등에 의해 발생하는 것으로 추측 하고 있지만 정확한 원인은 알려져 있지 않다.
최근에는 활동성 루푸스 환자의 혈청에 IFN-α가 증가해 있으며 (14,15), IFN-α을 치료제로 사용하는 경우에 자가항체의 생성과 루푸스와 유사한 증상이 초래되는 등 (16) IFN-α가 루푸스의 병인에 중요한 역할을 하는 것으로 알려졌다 (17). 또한, IFN-α가 루푸스의 면역체계에 미치는 영향에 대해서는 활동 성 루푸스 환자의 말초혈액의 단핵구 세포(PBMC)에 서 유전자 발현의 분석을 통해 규명되었다 (18).
IFN-α는 TCR signaling의 일부를 사용하여 작용을 하며, ERK의 활성화에 영향을 주지만 (19,20), 루푸 스 CD4+ T 림프구에서 TCR를 통한 ERK에 영향을 주어 병인에 관련되는지에 대해서는 정확히 알려져 있 지 않다.
본 연구자는 INF-α가 루푸스 CD4+ T 림프구에 서 TCR/CD3를 통한 ERK의 인산화의 이상에 중요 한 역할을 할 수 있는 것으로 가정하였다. 본 연구 에서는 루푸스 CD4+ T 림프구에는 anti-CD3 항체를 이용한 TCR을 통한 활성화에 의한 ERK-1/2의 인산 화가 감소해 있는 것을 알았다. 또한, 루푸스 환자의 혈청이 정상 대조군의 CD4+ T 림프구에서 anti-CD3 항체에 의한 ERK-1/2의 인산화를 감소시켰으며, 이 는 IFN-α에 대한 항체(anti-IFN-α)에 의해 억제되었 다. Recombinant IFN-α도 ERK-1/2의 인산화를 감소 시켰다. 이러한 결과는 IFN-α가 직접 CD4+ T 림프 구에 TCR을 통한 세포 내의 신호전달체계에 영향을 주어 루푸스의 병인에 중요한 역할을 할 수 있음을 시사해 준다.
대상 및 방법 1. 대상 환자 및 대조군
미국 류마티스학회의 개정된 분류 기준 (21)을 만 족하는 루푸스 환자(10명, 평균 연령, 47.1±3.1세, was decreased in control CD4+ T cells pre-incubation with lupus plasma with high IFN-α levels more than lupus plasma with non-detectable IFN-α levels. Recombinant IFN-α inhibited TCR-mediated ERK-1/2 phosphorylation dose-dependently.
Conclusion: These results suggest that IFN-α stimulation in vivo may underlie the aberrant TCR-mediated MAPK signaling in lupus CD4+ T cells and associated with disease patho- genesis.
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: CD4+ T cells, ERK signal, IFN-α, Systemic lupus erythematosus
19∼64세, 여자: 9명, 남자: 1명)와 정상 대조군(10명, 평균 연령 45.3±2.6세, 21∼63세, 여자: 9명, 남자: 1 명)을 대상으로 하였다. 정상 대조군과 루푸스 환자 및 미국 류마티스학회의 분류 기준 (22)을 만족하는 류마티스관절염 환자의 혈청을 얻어 사용하였다. 루 푸스의 활동성은 SLEDAI (systemic lupus erythemato- sus disease activity index) (23)를 이용하여 평가하였 으며, 5 이상은 루푸스의 활성도가 높은 군으로 정 하였고, 5 이하는 활성도가 낮은 것으로 하였다. 고 용량의 스테로이드(prednisolone>30 mg/day)나 면역 억제제를 투여중인 환자는 제외하였으며, 모든 대상 에게 서면 동의서를 받은 후 연구를 시행하였다.
2. 항체 및 시약
Human CD3에 대한 단클론 항체와 형광물질을 부 착한 항체로 anti-human CD4-APC, phospho-ERK-1/2 (T202/Y204)-Alexa Fluor488을 사용했으며(BD Phar- mingen, San Diego, CA), tyrosine/threonine dual pho- sphorylated forms of ERK-1/2 (phospho-ERK-1/2)을 인 식하는 p-ERK-1/2에 대한 단클론 항체와 donkey anti- goat, bovine anti-mouse, goat anti-rabbit HRP는 Santa Cruz Biotechnology (Santa Cruz, California, USA)에서 구입해 사용하였다. Recombinant IFN-α와 human IFN- α에 대한 neutralizing mouse mAbs 및 isotype control 항체는 R&D Systems에서 구입하였고, PMA, iono- mycin는 Sigma-Aldrich (St. Louis, MO)에서 구해 사 용하였다.
3. 세포 및 세포 배양
루푸스 환자와 정상 대조군에서 헤파린을 처리한 신선한 말초혈액을 얻어 Ficoll-Hypaque (Amersham Pharmacia Biotech, Uppsala, Sweden)을 이용한 den- sity gradient centrifugation를 통해 PBMC를 분리하였 다. CD4+ T 림프구는 human CD4+ T 림프구 sor- ting kit (Miltenyi Biotec, Auburn, CA)를 이용하여 PBMC로부터 분리하였으며, 순도는 APC-conjugated anti-CD4 항체를 이용하여 유세포분석기로 분석하였 을 때 >96%였다. CD4+ T 림프구는 필요할 경우 에 10% FCS를 함유하는 RPMI 1640 (Gibco BRL, Grand Island, NY)에서 배양하였다.
Anti-CD3 항체를 이용하여 TCR triggering을 통한
ERK의 인산화를 조사하기 위하여, 분리한 5×106 PBMC를 plate-bound anti-CD3 항체(10μg/mL)가 있는 48-well tissue culture plate에 넣어 37oC에서 10, 20, 30, 60분 동안 자극한 후에 유세포분석기로 분석하 였다. 혈청과의 배양 실험을 위하여 정상 대조군에서 분리한 CD4+ T 림프구를 L-glutamine (2 mm, Gibco BRL), penicillin (0.01 U/mL, Gibco BRL), strep- tomycin (0.01 g/mL, Gibco BRL)을 함유하는 complete RPMI 1640을 정상대조군, 루푸스 또는 류마티스 관 절염 환자의 혈청과 50%씩 혼합한 배양액에 5% CO2, 37oC에서 2일 동안 배양하였다. CD4+ T 림프구를 모아 complete RPMI 1640 medium으로 세척한 후에 plate-bound anti-CD3 항체(10μg/mL)로 48-well tissue culture plates에서 15분 동안 자극하였다. IFN-α, anti-IFN-α (10μg/mL) 또는 isotype control 항체 (10 μg/mL)의 효과를 보기 위하여 혈청과 배양하는 동안 첨가하였으며, recombinant IFN-α (500, 1,000, or 2,000 IU/mL)는 자극하기 2일 전에 처리하였다.
4. 유세포분석법
CD4+ T 림프구의 순도는 anti-CD4-PE와 isotype control 항체를 사용하여 유세포 분석법으로 조사하 였으며, 검사를 위하여 CD4+ T 림프구를 0.5%
BSA를 함유하는 wash buffer로 세척한 후에 해당하 는 항체와 isotype control 항체를 처리한 후에 검사 하였다. ERK-1/2의 인산화를 분석하기 위하여 2%
formaldehyde로 고정한 후에 90% methanol으로 per- meabilize한 후에 p-ERK-1/2-Alexa Fluor488 항체로 1 시간 동안 처리한 후 FACSan with CellQuest soft- ware (BD Bioscience)로 조사하고 Flow Jo software (Tree Star, Ashland, OR)를 이용하여 분석하였다.
ERK-1/2의 인산화 정도는 MFI (mean fluorescence in- tensity)를 측정하였다.
5. ELISA of IFN-α
Paired IFN-α capture와 detection 항체를 R&D Sys- tems에서 구해 혈청의 IFN-α를 측정하기 위하여 sandwich ELISA를 이용하였다.
6. 통계처리
모든 실험 결과는 평균±SEM으로 표기하였으며,
analysis of variance (ANOVA)를 이용하여 통계처리 하였다[SPSS 12.1 (SPSS, Chicago, IL)]. p values<
0.05를 유의한 차이가 있는 것으로 하였다.
결 과
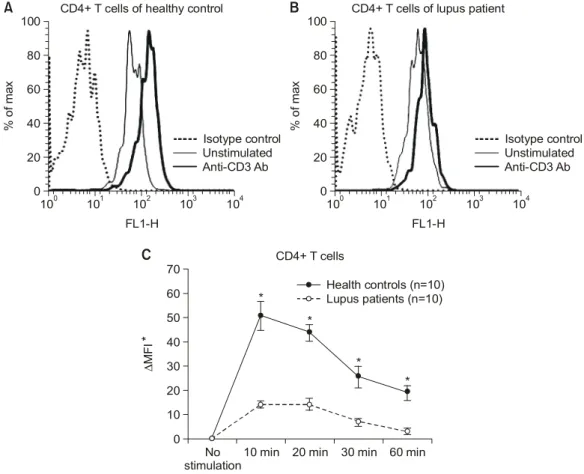
1. 루푸스 CD4+ T 림프구에서 anti-CD3 항체에 의한 ERK-1/2 인산화의 감소
CD4+ T 림프구의 TCR/CD3 complex를 통한 ERK- 1/2의 인산화를 조사하기 위하여 정상인과 루푸스
환자의 말초 혈액에서 PBMC를 분리한 후에 plate- coated anti-CD3 Ab (10 g/mL)를 이용하여 10, 20, 30, 60분 동안 자극하였다. 각 세포에서 유세포분석 기를 이용하여 자극 전후의 MFI의 차이를 구하였다.
anti-CD3항체에 의한 ERK-1/2의 인산화는 정상인에 비해 루푸스 CD4+ T 림프구에서 10, 20, 30, 60분 자극 후에 모두 유의하게 감소하였다(p<0.01 by Anova test) (그림 1). ERK-1/2는 두 군에서 모두 자 극 후 10∼20분 사이에 가장 많이 활성화되었으며, 이는 ERK-1/2 활성화의 변화가 kinetics의 이상에 의
Fig. 1. Decreased ERK-1/2 phosphorylation in lupus CD4+ T cells. ERK-1/2 phosphorylation was measured in CD4+
T cells from ten patients with SLE and same number of healthy controls before and after 10, 20, 30 and 60 min with plate-coated anti-CD3 Ab (10μg/mL) by flow cytometry. Representative experiments showing the histograms of the expression of p-ERK-1/2 in CD4+ T cells in healthy control (A) and lupus patients (B) before and after stimulation with anti-CD3 Ab. The difference of MFI (ΔMFI) of p- ERK-1/2 expression between MFI of before and after stimulation with anti-CD3 Ab in CD4+ T cells (C). Results are expressed as mean difference of MFI of p-ERK-1/2 between the MFI before and after stimulation with anti-CD3 Ab in CD4+ T cells (mean±
SEM). *p<0.01 compared with the results of lupus CD4+ T cells.
% of max
100 101 102 103 104 FL1-H
0 20 40 60 80
100 CD4+ T cells of healthy control
A
CD4+ T cells of lupus patient% of max
100 101 102 103 104 FL1-H
0 20 40 60 80 100
Isotype control Unstimulated Anti-CD3 Ab
B
*
CD4+ T cells
No
stimulation10 min 20 min 30 min 60 min 0
10 20 30 40 50 60 70
∆MFI*
Duration of stimulation with anti-CD3 Ab (10 g/mL)µ Health controls (n=10) Lupus patients (n=10)
*
*
*
C
Isotype control Unstimulated Anti-CD3 Ab
해서 발생하지 않았음을 의미한다. 이는 lupus-prone Fas-intact MRL/Mp+Fas-lpr (MRL/+Fas-lpr) mice에서 CD4+ T 림프구에 결함이 있다는 보고를 뒷받침해 준다 (5,6,24).
ERK1/2 인산화의 감소는 이환 기간, 치료 방법, ACR criteria 수, 신장 침범 여부, 항체의 존재 형태, 보체 등의 임상 소견과는 무관하였으나(data not shown), SLEDAI가 높은 군에서 낮은 군에 비해 더 컸다(p<0.01 by ANOVA test) (그림 2). 이는 ERK 활성화의 감소가 루푸스의 활성도와 관련이 있다는 기존의 보고와 일치한다 (12).
2. 루푸스 혈청에 의한 정상 CD4+ T 림프구의 ERK 인산화의 변화
루푸스 환자에서 CD4+ T 림프구의 ERK-1/2 활 성화의 결함은 알려져 있지만, 정확한 원인은 잘 알 려져 있지 않다. 루푸스 환자의 혈청에는 정상인에
비해 자가항체, 면역복합체, IFN-α와 같은 사이토카 인 등이 증가해 있으며 (15,16,25), 혈청에 의해 세포 내 전사인자의 발현이 변할 수 있다 (26). 혈청인자 가 TCR을 통한 ERK-1/2의 인상화에 미치는 영향을 보기 위하여 정상인의 CD4+ T 림프구를 RPMI 1640와 혼합한 환자의 혈청과 2일 동안 배양한 후 anti-CD3 항체(10μg/mL)로 15분간 자극하고 ERK- 1/2의 인산화를 유세포분석기로 분석하였다. 정상인 이나 류마티스 관절염 환자의 혈청과 배양한 경우 보다 루푸스 환자의 혈청과 배양한 CD4+ T 림프구 에서 ERK-1/2의 인산화가 유의하게 감소하였다(그림 3A). 이는 혈청 내 인자가 루푸스 환자의 CD4+ T 림프구의 ERK pathway의 결함의 원인이 될 수 있음 을 시사한다.
IFN-α는 TCR signaling의 일부를 사용하여 작용 하며 (27), ERK signal에 억제 효과를 갖고 있어 (19,20) IFN-α가 루푸스 CD4+ T 림프구의 ERK sig- nal 결함과 관련이 있을 것으로 가정하였다. 루푸스 혈청에 의한 ERK pathway에 미치는 영향이 IFN-α 에 의한 것인지를 알아보기 위하여 anti-IFN-α 또는 isotype control 항체를 2일간 처리한 후에 동일한 실 험을 하였으며, anti-IFN-α 항체는 루푸스 혈청에 의 한 ERK-1/2 인산화의 감소를 억제하였다(그림 3B).
루푸스 혈청에 의한 ERK-1/2의 인산화에 미치는 효 과를 규명하기 위하여 IFN-α가 높은 루푸스 혈청과 낮은 혈청을 이용하여 위와 같은 실험을 하였으며, IFN-α가 높은 루푸스 혈청이 ERK-1/2의 인산화를 보다 더 감소시키는 것을 알 수 있었다(그림 3C).
이는 IFN-α가 루푸스 CD4+ T 림프구의 TCR을 통 한 ERK signaling의 결함을 일으켜 병인에 관련될 수 있음을 시사한다.
3. CD4+ T 림프구에서 anti-CD3 항체의 자극에 의한 ERK 인산화가 IFN-α에 의해 감소
마지막으로 IFN-α가 TCR/CD3 complex에 의한 ERK의 인산화에 미치는 영향을 알아보기 위해서 recombinant IFN-α (1,000 IU/mL)를 정상인의 CD4+
T 림프구와 2일 동안 배양 후 plate-coated anti-CD3 항체(10μg/mL)로 15분간 자극한 후에 유세포분석기 로 ERK-1/2의 인산화를 조사하였으며, IFN-α이 CD4+
T 림프구의 ERK-1/2 인산화를 억제하고, IFN-α의 Fig. 2. Association of altered ERK-1/2 phosphorylation
with disease activity of SLE (SLEDAI) in lupus CD4+ T cells. The degree of ERK-1/2 phos- phorylation was compared between lupus patients with high disease activity (SLEDAI≥5) and pa- tients with low disease activity (SLEDAI<5) in CD4+ T cells after stimulation with plate-coated anti-CD3 Ab (10μg/mL) for 10, 20, 30 and 60 min. Results are expressed as mean difference of MFI of p-ERK-1/2 between the MFI before and after stimulation with anti-CD3 Ab in CD4+ T cells (mean±SEM). *p<0.01 compared with the results of lupus CD4+ T cells with SLEDAI≥5.
용량에 따라 ERK-1/2의 인산화가 감소함을 관찰할 수 있었다(그림 4).
고 찰
본 연구에서는 정상 대조군에 비해 루푸스 환자의
CD4+ T 림프구에서 anti-CD3 항체에 의한 ERK-1/2 의 인산화가 감소해 있는 것을 보여주고 있다. 루푸 스 CD4+ T 림프구에서 ERK-1/2 인산화가 감소한 원인으로는 ERK-1/2의 catalytic activity의 내인적 결 함과 관련이 없는 것으로 제시되고 있지만, 정확한 원인은 잘 알려져 있지 않다. 본 연구에서는 TCR Fig. 3. The effects of serum factors on the ERK-1/2 phosphorylation of CD4+ T cells. CD4+ T cells from healthy controls were pre-incubated in complete RPMI 1640 containing 50% sterile plasma from control, lupus or RA patients for 2 d and washed with complete RPMI and activated using plate-coated anti-CD3 Ab (10μg/mL) for 15 min and analyzed for ERK-1/2 phosphorylation by flow cytometry. Representative experiments showing the difference of MFI of p-ERK-1/2 between before and after TCR triggering with anti-CD3 Ab in control CD4+
T cells pre-incubated in different plasma (A). Neutralizing anti-IFN-α Ab reverse the effects of lupus soluble factors on the ERK-1/2 phosphorylation in CD4+ T cells, but not in isotype control Ab. MAPK inhibitor, U0126 was used as a positive control (B). Comparison of the effects of lupus plasma with high IFN-α level (>100 ng/mL) and lupus plasma with un-detectable IFN-α level on the ERK-1/2 phosphorylation by TCR triggering (C). Results are expressed as mean difference of MFI of p-ERK-1/2 between the MFI before and after stimulation with anti-CD3 Ab in CD4+ T cells (mean±SEM). ΔMFI: Difference in the MIF of p-ERK-1/2 between stimulated and un-stimulated CD4+ T cells with anti-CD3 Ab (10μg/mL), #U 0126: MAPK kinase inhibitor,
Ⓛanti-IFN-α: anti-IFN-α Ab (10μg/mL), *p-value: 0.01 compared with self and allogeneic normal plasma.
signaling에 영향을 줄 수 있는 것으로 알려진 (19,27) IFN-α가 TCR/CD3 complex에 의한 ERK-1/2의 활성 화를 억제하고 이를 통하여 루푸스의 병인에 중요한 역할을 할 수 있음을 시사해 준다.
IFN-α는 apoptotic cell fragments, DNA, RNA 등에 대한 자가항체의 영향으로 생성될 수 있으며 (28), dendritic cells, T, B 림프구 및 natural killer cells과 단핵구 세포 등에 영향을 주어 autoreactive T 림프구 를 갖는 T-cell repertoire를 특징으로 하는 면역체계 의 이상을 일으키는 것으로 알려져 있다 (14,29).
IFN-α는 항원제시세포를 통하여 T 세포를 자극하여 hyper-responsive T 림프구의 생성을 증가시킬 수 있 다 (30). IFN-α는 T 림프구의 생존을 증가시키거나 (31), 사이토카인의 생성에 영향을 주어 T 림프구의 면역조절에 영향을 주지만 (32), 직접 CD4+ T 림프 구의 세포 내 신호전달체계에 영향을 주어 루푸스의 병인에 관련이 있는지에 대해서는 잘 알려져 있지 않다.
비록 루푸스 CD4+ T 림프구의 ERK-1/2 인산화 의 결함이 병인에 중요한 역할을 하는 것에 대해서 는 잘 알려져 있지만, 이에 대한 정확한 원인은 밝 혀지지 않았다. 루푸스 환자의 혈청은 정상 대조군 과 다르게 자가항체, 면역복합체, IFN-α, IL-6 등의
사이토카인 등을 함유하고 있으며 이는 다양한 세포 에 영향을 줄 수 있다 (33). 따라서 루푸스 혈청이 TCR/CD3 complex를 통한 ERK signaling의 결함과 관련이 있을 것으로 가정하였으며, 이를 규명하기 위하여 정상 대조군의 CD4+ T 림프구에서 anti- CD3 항체에 의한 ERK-1/2의 인산화에 루푸스 혈청 이 미치는 영향을 조사하였다. 본 연구에서 류마티 스 관절염 환자나 정상 대조군의 혈청에 비해 루푸 스 혈청이 ERK-1/2의 인산화를 억제하는 것을 관찰 하였으며, 이는 루푸스 혈청에 존재하는 혈청인자가 루푸스의 ERK pathway의 결함과 관련이 있음을 시 사한다.
IFN-α는 TCR의 세포내 신호전달체계인 Lck, ZAP70를 통해T 림프구의 증식을 억제하며 (27), T 림프구와 T 세포주에서도 IFN-α가 ERK의 활성화를 억제하여 성장 억제를 일으키는 것으로 알려져 있다 (19,20). 이러한 결과에 따라 본 연구자는 IFN-α가 루푸스 CD4+ T 림프구의 ERK-1/2의 결함과 관련 이 있을 것으로 가정하였다. 본 연구에서는 루푸스 혈청에 의한 효과가 anti-IFN-α 항체에 의해서 억제 되고, 특히 IFN-α를 보다 많이 함유하는 루푸스 혈 청이 낮은 혈청에 비해 더욱 ERK의 활성화를 억제 하는 것을 알 수 있었다. 또한, recombinant IFN-α Fig. 4. The effects of recombinant IFN-α on the ERK-1/2 phosphorylation of CD4+ T cells. Control CD4+ T cells were pre-incubated with IFN-α (1,000 IU/mL) for 2 d and washed two times with complete RPMI 1640 and activated using plate-coated anti-CD3 Ab (10μg/mL) for 15 min and analyzed for ERK-1/2 phosphorylation by flow cytometry (A). The effects of IFN-α on ERK activation were also evaluated with increasing dosage of IFN-α (500, 1,000, OR 2,000 IU/mL) (B). Results are expressed as mean difference of MFI of p-ERK-1/2 between the MFI before and after stimulation with anti-CD3 Ab with or without IFN-α in CD4+ T cells (mean±
SEM). *p-value: 0.05 compared with the ΔMFI of CD4+ T cells stimulated with anti-CD3 without IFN-α.
∆MFI
0 500 1,000 2,000
* 0
20 40 60 80
% of max
100 101 102 103 104 FL1-H
0 20 40 60 80
A
100No stimulation INF- and aCD3α aCD3
Isotype control
B
*
가 정상 CD4+ T 림프구의 ERK-1/2 인산화를 감소 시키는 것을 알 수 있었다. 자가면역 질환의 발생에 사이토카인 생성의 이상이 중요한 역할을 하며, 루 푸스와 류마티스 관절염에서 IFN-α 생성의 비정상 적인 조절이 있는 것은 (34) 본 연구 결과를 뒷받침 해준다. 그렇지만 다른 사이토카인이나 자가 항체와 같은 혈청인자의 영향에 대해서는 추가적인 연구가 필요하다.
본 연구에서는 루푸스의 활성도가 CD4+ T 림프 구의 ERK-1/2의 결함과 관련이 있을 것으로 가정하 였으며, SLEDAI score가 높은 환자에서 낮은 환자에 비해 TCR/CD3 complex를 통한 ERK-1/2의 인산화가 유의하게 감소한 것을 알 수 있었다. 루푸스 혈청이 ERK-1/2의 활성화에 미치는 영향이 혈청 IFN-α 농 도와 관련이 있으며, 실제 IFN-α가 루푸스의 활성도 와 관련이 있다는 보고 (18)는 본 연구 결과를 지지 해 준다. 그렇지만, 본 연구는 적은 수의 환자를 대 상으로 하였고 루푸스 활성도가 높지 않은 환자를 대상으로 하였다는 제한이 있지만, ERK-1/2의 활성 화가 루푸스의 활성도와 관련이 있다는 기존의 보고 (12)와 일치한다.
DNA methylation은 chromatin의 구조와 유전자의 발현에 필수적이며, DNMT에 의한 CG pairs의 de- oxycytosine (dC) bases를 methylation하는 것은 유전 자의 발현을 억제하는 기전이 된다 (35). DNMT의 발현은 MAPK signaling pathway와 activator protein 1 (AP-1)에 의해서 조절되며 (36-38), 루푸스에서는 MAPK signaling의 결함이 DNMT의 발현의 감소의 원인이 되며 (12), 이는 LFA-1, CD70, perforin과 여 러 가지 cytokines과 같은 hypomethylation에 민감한 유전자의 과다발현을 일으켜 병인에 관련되는 것으로 알려져 있다. 따라서, 향후에는 IFN-α에 의한 ERK- 1/2 인산화의 감소가 DNMT의 발현에 미치는 영향 과 hypomethylation에 민감한 유전자의 발현에 영향을 주어 CD4+ T 림프구의 autoreactivity의 원인이 될 수 있는지에 대해서는 연구가 필요할 것으로 보인다.
결 론
본 연구에서는 루푸스 CD4+ T 림프구는 질환의 활성도와 비례하여 ERK의 결함이 있었으며, 루푸스
혈청은 IFN-α와 관련되어 이러한 세포내의 신호전 달체계의 결함과 관련이 있었다. Recombinant IFN-α 는 CD4+ T 림프구에서 TCR/CD3 complex를 통한 ERK signaling의 이상을 초래하였다. 이러한 결과는 IFN-α가 루푸스 CD4+ T 림프구의 ERK signaling에 직접적으로 결함을 야기하여 병인에 중요한 역할을 할 수 있을 것으로 보인다. 그렇지만 IFN-α에 의한 CD4+ T 림프구의 ERK의 활성화에 미치는 영향이 루푸스의 병인에 어떻게 영향을 줄 수 있는가에 대 한 구체적인 기전을 알기 위한 연구가 필요할 것으 로 생각된다.
REFERENCES
1) Hardin JA. The lupus autoantigens and the path- ogenesis of systemic lupus erythematosus. Arthritis Rheum 1986;29:457-60.
2) Morrow JNL, Watts R, Isenberg D. Systemic lupus erythematosus. Autoimmune rheumatic disease. Oxford, United Kingdom, Oxford University Press, 1999.
3) Dayal AK, Kammer GM. The T cell enigma in lupus.
Arthritis Rheum 1996;39:23-33.
4) Tsokos GC, Liossis SN. Immune cell signaling defects in lupus: activation, anergy and death. Immunol Today 1999;20:119-24.
5) Zielinski CE, Jacob SN, Bouzahzah F, Ehrlich BE, Craft J. Naive CD4+ T cells from lupus-prone Fas- intact MRL mice display TCR-mediated hyperproli- feration due to intrinsic threshold defects in acti- vation. J Immunol 2005;174:5100-9.
6) Vratsanos GS, Jung S, Park YM, Craft J. CD4(+) T cells from lupus-prone mice are hyperresponsive to T cell receptor engagement with low and high affinity peptide antigens: a model to explain spontaneous T cell activation in lupus. J Exp Med 2001;193:329-37.
7) Tsokos GC, Nambiar MP, Tenbrock K, Juang YT.
Rewiring the T-cell: signaling defects and novel prospects for the treatment of SLE. Trends Immunol 2003;24:259-63.
8) Yi Y, McNerney M, Datta SK. Regulatory defects in Cbl and mitogen-activated protein kinase (extra- cellular signal-related kinase) pathways cause persis- tent hyperexpression of CD40 ligand in human lupus T cells. J Immunol 2000;165:6627-34.
9) Desai-Mehta A, Lu L, Ramsey-Goldman R, Datta SK.
Hyperexpression of CD40 ligand by B and T cells in
human lupus and its role in pathogenic autoantibody production. J Clin Invest 1996;97:2063-73.
10) Treisman R. Regulation of transcription by MAP kinase cascades. Curr Opin Cell Biol 1996;8:205-15.
11) Seger R, Krebs EG. The MAPK signaling cascade.
FASEB J 1995;9:726-35.
12) Deng C, Kaplan MJ, Yang J, Ray D, Zhang Z, McCune WJ, et al. Decreased ras-mitogen-activated protein kinase signaling may cause DNA hypo- methylation in T lymphocytes from lupus patients.
Arthritis Rheum 2001;44:397-407.
13) Kammer GM, Tsokos GC. Lupus: molecular and cellular pathogenesis. p. 231-560, Totowa, NJ, The Humana Press, 1999.
14) Hooks JJ, Moutsopoulos HM, Geis SA, Stahl NI, Decker JL, Notkins AL. Immune interferon in the circulation of patients with autoimmune disease. N Engl J Med 1979;301:5-8.
15) Preble OT, Black RJ, Friedman RM, Klippel JH, Vilcek J. Systemic lupus erythematosus: presence in human serum of an unusual acid-labile leukocyte interferon. Science 1982;216:429-31.
16) Crow MK, Kirou KA, Wohlgemuth J. Microarray analysis of interferon-regulated genes in SLE. Auto- immunity 2003;36:481-90.
17) Ronnblom LE, Alm GV, Oberg KE. Possible induction of systemic lupus erythematosus by inter- feron-alpha treatment in a patient with a malignant carcinoid tumour. J Intern Med 1990;227:207-10.
18) Bennett L, Palucka AK, Arce E, Cantrell V, Borvak J, Banchereau J, et al. Interferon and granulopoiesis signatures in systemic lupus erythematosus blood. J Exp Med 2003;197:711-23.
19) Romerio F, Zella D. MEK and ERK inhibitors enhance the anti-proliferative effect of interferon- alpha2b. FASEB J 2002;16:1680-2.
20) Caraglia M, Tagliaferri P, Marra M, Giuberti G, Budillon A, Gennaro ED, et al. EGF activates an inducible survival response via the RAS->Erk-1/2 pathway to counteract interferon-alpha-mediated ap- optosis in epidermoid cancer cells. Cell Death Differ 2003;10:218-29.
21) Tan EM, Cohen AS, Fries JF, Masi AT, McShane DJ, Rothfield NF, et al. The 1982 revised criteria for the classification of systemic lupus erythematosus. Arthri- tis Rheum 1982; 25:1271-7.
22) Arnett FC, Edworthy SM, Bloch DA, McShane DJ, Fries JF, Cooper NS, et al. The American Rheu- matism Association 1987 revised criteria for the clas-
sification of rheumatoid arthritis. Arthritis Rheum 1988;31:315-24.
23) Hawker G, Gabriel S, Bombardier C, Goldsmith C, Caron D, Gladman D. A reliability study of SLEDAI:
a disease activity index for systemic lupus ery- thematosus. J Rheumatol 1993;20:657-60.
24) Bouzahzah F, Jung S, Craft J. CD4+ T cells from lupus-prone mice avoid antigen-specific tolerance induction in vivo. J Immunol 2003;170:741-8.
25) Bengtsson AA, Sturfelt G, Truedsson L, Blomberg J, Alm G, Vallin H, et al. Activation of type I interferon system in systemic lupus erythematosus correlates with disease activity but not with antiretroviral anti- bodies. Lupus 2000;9:664-71.
26) Juang YT, Wang Y, Solomou EE, Li Y, Mawrin C, Tenbrock K, et al. Systemic lupus erythematosus se- rum IgG increases CREM binding to the IL-2 pro- moter and suppresses IL-2 production through CaMKIV. J Clin Invest 2005;115:996-1005.
27) Petricoin EF 3rd, Ito S, Williams BL, Audet S, Stancato LF, Gamero A, et al. Antiproliferative action of interferon-alpha requires components of T-cell- receptor signalling. Nature 1997;390:629-32.
28) Vallin H, Perers A, Alm GV, Ronnblom L. Anti- double-stranded DNA antibodies and immunostimul- atory plasmid DNA in combination mimic the endog- enous IFN-α inducer in systemic lupus erythema- tosus. J Immunol 1999;163:6306-13.
29) Stewart TA. Neutralizing interferon alpha as a ther- apeutic approach to autoimmune diseases. Cytokine Growth Factor Rev 2003;14:139-54.
30) Dalod M, Hamilton T, Salomon R, Salazar-Mather TP, Henry SC, Hamilton JD, et al. Dendritic cell responses to early murine cytomegalovirus infection:
subset functional specialization and differential reg- ulation by interferon alpha/beta. J Exp Med 2003;
197:885-98.
31) Matikainen S, Sareneva T, Ronni T, Lehtonen A, Koskinen PJ, Julkunen I. Interferon-alpha activates multiple STAT proteins and upregulates prolifera- tion-associated IL-2Ralpha, c-myc, and pim-1 genes in human T cells. Blood 1999;93:1980-91.
32) Eriksen KW, Sommer VH, Woetmann A, Rasmussen AB, Brender C, Svejgaard A, et al. Bi-phasic effect of interferon (IFN)-α: IFN-alpha up- and down-re- gulates interleukin-4 signaling in human T cells. J Biol Chem 2004;279:169-76.
33) Linker-Israeli M, Wallace DJ, Prehn J, Michael D, Honda M, Taylor KD, et al. Association of IL-6 gene
alleles with systemic lupus erythematosus (SLE) and with elevated IL-6 expression. Genes Immun 1999;
1:45-52.
34) Palucka AK, Blanck JP, Bennett L, Pascual V, Banchereau J. Cross-regulation of TNF and IFN-α in autoimmune diseases. Proc Natl Acad Sci USA 2005;102:3372-7.
35) Attwood JT, Yung RL, Richardson BC. DNA methy- lation and the regulation of gene transcription. Cell Mol Life Sci 2002;59:241-57.
36) Rouleau J, MacLeod AR, Szyf M. Regulation of the DNA methyltransferase by the Ras-AP-1 signaling pathway. J Biol Chem 1995;270:1595-601.
37) MacLeod AR, Rouleau J, Szyf M. Regulation of DNA methylation by the Ras signaling pathway. J Biol Chem 1995;270:11327-37.
38) Yang BC, Wang YS, Lin LC, Liu MF. Induction of apoptosis and cytokine gene expression in T-cell lines by sera of patients with systemic lupus erythema- tosus. Scand J Immunol 1997;45:96-102.