치아 우식증에 따른 치수내 유전자 발현 변화에 관한 분석
오소희*∙김종수**
한림대학교 성심병원 소아치과*, 단국대학교 치과병원 소아치과**
치아 우식증은 범발성 질환이고 이에 대한 생체반응은 단순하지 않으며 질병 과정과 숙주의 활성 모두를 반영하는 복합적 인 반응이다. 이러한 반응을 이해하기 위해서는 질병의 세포학적, 분자학적인 면을 이해하는 것이 필수적이다.
이에 본 연구는 임상적으로 건강한 치아와 우식이 진행된 치아로부터 얻어진 치수 안의 유전자 발현을 규명하고 우식 병소 에서 일어나는 치유 및 재생에 관계되는 분자와 면역 세포들 사이의 분자 생화학적 상호작용을 규명하기 위해서 우식치아와 건전치아의 치수를 이용하여 cDNA 미세배열(microarray) 분석과 역전사효소 중합효소 연쇄반응 (RT-PCR) 분석, 그리고 면역화학염색법 (immunohistochemistry)을 시행하여 다음과 같은 결론을 얻었다.
1. cDNA 미세배열 분석 결과, 건전치아군인 대조군에서는 143개의 유전자가, 우식치아군인 실험군에서는 377개의 유전 자가 1.6배이상 발현되었다.
2. 역전사효소 중합효소 연쇄반응 분석에서 14개의 유전자를 선택하였고 cDNA 미세배열 분석결과와 동일한 결과를 확인 하였다.
3. TGF-β1의 면역조직화학적 관찰 결과, 건전치에 비해 우식치의 상아모세포와 치수에서 특히 강하게 발현되었다.
주요어: 치아 우식증, cDNA 미세배열 분석, 역전사효소 중합효소 연쇄반응 분석, 면역화학염색법 국문초록
Ⅰ. 서 론
치아 우식증은 범발성 질환으로 유전, 식이습관, 구강내 숙 주 환경등의 여러 요소가 관련되는 질병이다1). 치아 우식증의 치료는 감염된 병소를 제거하고 치수를 보호하고 인공적인 재 료로 이를 수복하는 여러 과정이 관여하고 있다. 치료의 성공은 환자의 전신적 조건, 수복 계획, 우식의 정도, 치수의 증상, 손 상의 정도, 치수 복조 약제 등에 따라 좌우된다2). 치수 생활력 보존은 현재의 수복 치과학에서 도전되는 과제이며 손상받은 상아질과 치수의 재생(regeneration)과 회복(repair)은 그 목 표가 되고 있다.
상아질-치수 복합체의 생활력 유지에서 중요한 점은 치아에 가해지는 해로운 자극과 손상에 계속적으로 반응하는 것이다.
그러나 반응의 양상과 정도는 상아질-치수 복합체에서 조직의 조건과 손상의 범위를 반영하고 있다. 치아 우식증은 한가지 반 응이 아니라 질병 과정과 숙주의 활성 모두를 반영하는 복합적
인 반응이고 이러한 반응을 이해하기 위해서는 질병의 세포학 적, 분자생물학적인 면을 이해하는 것이 필수적이다3,4).
1990년에 시작된 인간유전체사업(Human genome pro- ject)의 결과로 인간 유전체 서열 정보의 해독을 통하여 유전자 의 기능을 규명, 유전체 정보 등을 연구할 수 있는 토대가 마련 되었다. 그러나 인간의 복잡한 생명현상을 규명하기 위해서는 유전체의 서열정보만으로는 부족하며 단 간의 상호작용 및 유 전자 발현여부 등 추가적인 생물학적 정보들이 필요하게 되어 유전체의 기능을 밝히려는 기능 유전체학(functional ge- nomics)이 대두되었고 이의 중요한 도구가 cDNA microarray 이다. 기존의 유전공학 방법들의 대부분은 연구자가 동시에 많 은 수의 유전자에 대해 한꺼번에 실험할 수 없는 제약이 있었기 때문에“one-gene in the one-experiment"에 기반을 둔 연구 가 수행되었다. 그러나“one-gene in the one-experiment"의 방법으로는 유전자 전체의 발현을 보기에 어려운 점이 많았다5). cDNA microarray 방법은 세포의 발생, signal transduc-
교신저자 : 김 종 수
충남 천안시 신부동 산 7-1 / 단국대학교 치과병원 소아치과 / 041-550-1935 / [email protected] 원고접수일: 2010년 04월 02일 / 원고최종수정일: 2010년 07월 23일 / 원고채택일: 2010년 08월 02일
tion(신호 형질 도입), 질병의 진단과 특징, 치료 반응 등을 연 구하기 위해 보다 빠르고 효과적으로 동시에 몇 만개의 유전자 발현 분석을 할 수 있다6,7).
cDNA microarray 기술은 기능 유전체학(functional ge- nomics)을 발전시켰으며 오늘날 질병 진행 과정동안 발생하는 분자세포학적으로 일어나는 일들을 알게 해주는 믿을 만한 방 법이다8).
치아 우식증에 관한 이러한 분석은 재생 및 회복에 관계되는 분자와 면역 세포들 사이의 상호관계의 결과로 나타나는 세포 활성기전, 유전자 발현, 특정 세포의 관련정도 등을 알려준다9).
우식 병소 아래의 치수의 분자 세포학적 반응은 미래의 진단 과 치료의 향상에 많은 공헌을 할 것이다. 본 연구의 목적은 임 상적으로 건강한 치아와 우식 치아로부터 얻어진 치수 안의 유 전자 발현을 규명하고 우식 병소에서 일어나는 분자 생물학적 상호작용을 특징 짓기 위함이다.
Ⅱ. 연구재료 및 방법 1. 연구 재료
치아 우식이 없는 건전한 제3대구치 30개와 법랑질에서 상아 질까지 우식이 진행된 제3대구치 30개를 본 실험에 사용하였 다. 환자의 연령은 20세에서 30세 사이였고 실험을 위해 치아 를 사용한다는 환자의 동의를 받았다. 우식 병소의 정도는 임상 적 검사 및 방사선적 검사로 확인하여 법랑질에서 초기 및 중등 도의 상아질 우식이 진행된 1급에서 2급정도의 치아를 선택하 였다.
발치된 치아는 즉시 RNA stabilizing solution인 RNA Later(Sigma, UK)에 담가둔 후 diamond disc를 이용하여 치 아를 치축방향으로 절단하고, 치수만을 보존하여 조심스럽게 분리하여 즉시 조직표본 제작을 위해 -196℃의 액화 질소에 넣 어 급냉 하였다.
2. Total RNA 추출(extraction)
Total RNA는 Trizol reagent(Invitrogen Co., USA)를 이 용하여 추출하였으며 추출방법은 제조사에서 제공한 Trizol protocol을 사용하였다. RNA의 농도와 순도는 Biophotometer (Eppendorf Co., UK)를 사용하여 260 nm와 280 nm에서 흡광도를 측정하였고 포름알데히드가 포함된 아가로스 전기영 동을 사용하여 100 voltage에서 측정하였다.
본 연구에서 사용된 RNA 표본들은 260/280 nm ratio가 0.5/0.25이었고 28s/18s rRNA ratio가 1.94이었다.
3. cDNA 미세배열 분석(microarray analysis)
건전치와 우식치로부터 얻어진 치수 표본들 사이의 유전자 발현 분석을 위해 21,927개의 human sequence(Clontech
L., USA)를 이용하여 cDNA 미세배열기술(microarray)로 screening하였다. 모든 과정은 cDNA Microarray Assembline Kit(Agilent Technologies Inc., USA)를 이용하여 컴퓨터 제 어로 전과정 시행되었다.
Total RNA에서 cDNA를 합성하기 위하여 total RNA를 대 조군과 실험군 모두 5 μg씩 취해서 random primer로 70℃에 서 5분간 denaturation시킨 후 10 mM dNTPs(Roche, Germany) 2 μl, RNase inhibitor 1 μl, AMV(Avian Myoblastosis Virus) reverse transcriptase (Promega, USA) 2 μl, 5xbuffer(Promega, USA) 8 μl를 42℃에서 1시 간 반응시켰다. Cy5와 Cy3(Amersham Pharmacia Biotech, UK)로 형광 레이블링 하였다. 마이크로어레이 슬라이드에 미 리 심어 놓은 DNA Probes 위에 labeled target cDNA를 넣고 cover glass를 덮고 50℃에서 16시간 동안 hybridization(혼 성화)하였다.
cDNA microarray는 GSI Lumonics SA 5000 scanner (GSI Lumonics, Watertown, MA, USA)를 이용하여 스캐닝 한 후 QUANTARRAY-program(GSI Lumonics)를 이용하 여 image analysis를 시행하였다. 양쪽 label이 하나의 혼합 영상으로 결합되어 Cy3:Cy5 강도 비율로 나타났고 건전치아 에 대한 우식치에서의 유전자의 다른 발현을 분석하기 위해서 Cy3:Cy5의 강도 비율이 QUANTARRAY-program으로부터 각 점에서 계산되었다. 우식치와 건전치 사이에서 유전자 발현 의 차이가 1.6배 이상 차이가 나는 값만을 선택하였다.
4. 역 전 사 효 소 중 합 효 소 연 쇄 반 응 분 석 (Reverse Transcriptase-Polymerase Chain Reaction (RT- PCR) Analysis)
RT-PCR 분석은, cDNA microarray 분석결과를 다른 방법 으로 다시 한번 확인하기 위하여 시행되었다. 선택한 유전자는 human genes tumor necrosis factor superfamily member 14 (TNF member 14), transforming growth factor alpha (TGF-α), major histocompatibility complex class I (MHC complex I), fibroblast growth factor receptor 3 (FGF- R3), bone-gamma-carboxyglutamate protein (osteocal- cin), transforming growth factor beta 1 (TGF-β1), en- dothelial cell growth factor 1 (EGF-1), Sp7 transcription factor, nestin, phospholipase A2 group IVA, CD69 anti- gen, pyruvate dehydrogenase complex, secreted phos- phoprotein 1, osteomodulin이다. Table 1은 semi-quanti- tative 와 real-time RT-PCR에 사용된 우식 치아에서 증가된 유전자 중 선택된 9개의 유전자의 primer design을 보여주고 Table 2는 우식 치아에서 감소된 유전자 중 선택된 5개의 유전 자의 primer design을 보여준다. 정량된 cDNA 0.1 μg에 10×
Reaction buffer(100 mM Tris-Cl, pH 8.3, 500 mM KCl, 0.01% gelatin), 25 mM MgCl2, 10 mM dNTP, 10 nm sense 및 antisense primer, 1 unit Taq DNA polymerase
및 증류수를 가하여 총량을 50 μl로 한 후, 95℃에서 10분 동안 반응한 후 다음 cycle부터는 95℃에서 1분 동안 반응하고 다시 60℃에서 20초, 72℃에서 20초 동안, 마지막 cycle에서는 72℃
에서 10분 동안 총 반응을 32회 시행하였다. mRNA level의 정확한 정량을 위해 human house keeping gene인 GAPDH 로 확인하였다. PCR반응이 종료된 검체들에서 각 산물을 10 μl 씩 취하여 15% 한천 겔(agarose gel)에 100 V에서 30분간 전 기영동하고 gel document system(Bio-Rad, USA)에서 분석 하였다.
5. 면역조직화학적 관찰(immunohistochemistry)
면역조직화학적 관찰을 위해서 avidin-biotin conjugate (ABC)법을 사용하여 면역 염색을 시행하였으며 일차 항체로는 흰쥐의 TGF-β1(Santa Cruze, USA) 항체를 사용하였다. 우 식치아와 건전치아의 치수만을 분리해 내어 5 μm의 절편을 제 작하였다. 조직절편의 파라핀을 xylen으로 제거하고 일련의 에
탄올과 증류수로 각 5분씩 재함수시켰다. 항체의 비특이적 반 응을 방지하기 위해 normal blocking serum(Dako Co., Denmark)으로 30분간 처리하였다. 여분의 혈청을 세척하고 절편에 TGF-β1에 대한 1차 항체(Santa Cruze, USA)를 희석 하여 적용시킨 후, 조직 절편이 건조되지 않도록 습도를 유지한 상태로 4℃에서 24시간동안 반응시켰다. 조직절편을 이차항체 인 anti-mouse IgG(Dako Co., Denmark)를 실온에서 15분 간 처리하였다. Streptavidine reagent(Dako Co., Denmark) 로 15분간 반응시켰고 필요한 경우 Mayer's hematoxylin으 로 3분간 대조 염색하여 Canada Balsam(Sigma, USA)으로 봉입하고 광학 현미경으로 관찰하였다. Negative controls은 primary antibody를 대신하여 PBS로 처리하였고 positive control은 oral squamous cell carcinoma를 사용하였다. 모든 염색은 2명의 조사자에 의해 독자적으로 표본을 알지 못하는 상태에서 관찰되었으며 labeling evaluation은 negative(-), weakly positive(+), positive(++), stongly positive(++) 로 평가되었다.
Table 2. Primer sequence of down-regulation genes
No Gene Name Sequence(5'-3') Product size(bp)
1Phospholipase A2(group IVA) AGG CAC CAG AGC CTT GAG TA 424
GGT GAG GTA CCC ACA CCA TC
2 CD69 antigen AGT CCC CAT TTC TCA ACA CG 477
CAT GCT GCT GAC CTC TGT GT
3 Pyruvate dehydrogenase complex TGC TGA CTG TGA CCT TGG AG 468
TTG GCA TTT CCC TCT TCA TC
4 Secreted phosphoprotein 1ACA GCC AGG ACT CCA TTG AC 422
GTG GGT TTC AGC ACT CTG GT
5 Osteomodulin AAT CAA TGC CTC CTG GTT TG 433
TTA GTG CTT CGT TGT TCA CCA Table 1. Primer sequence of up-regulation genes
No Gene Name Sequence(5'-3') Product size(bp)
1TNF member 14 AGT GTC TGC AGT GCC AAA TG 478
TCA CCT TCT GCC TCC TGT CT
2 TGF-α AAT CCA TCA GCA GGG ATC TG 404
CTT GAG GGG AAA GGA AGA CC
3 Major histocompatibility complex I AAT CCA TCA GCA GGG ATC TG 404
CTT GAG GGG AAA GGA AGA CC
4 FGF-R3 CAT GAG CTC CAA CAC ACC AC 497
ACA CCA GGT CCT TGA AGG TG
5 Bone-gamma-carboxyglutamate TGA GAG CCC TCA CAC TCC TC 450
(osteocalcin) TCT GGA GTT TAT TTG GGA GCA
6 TGF-β1GGG ACT ATC CAC CTG CAA GA 421
CAC GTG CTG CTC CAC TTT TA
7 EGF-1GCA CCT TGG ATA AGC TGG AG 461
GAC CAG GTC CCT TAA GTC TGG
8 Sp7 transcription factor ATG TCT TGC CCC AAG ATG TC 408
GCA GGC AGG TGA ACT TCT TC
9 Nestin GCC CTG ACC ACT CCA GTT TA 471
GGG ATT GCA ATG TCT GTG TG
Ⅲ. 연구 결과
1. cDNA 미세배열 분석(microarray analysis)
cDNA 미세배열 분석(microarray analysis)의 실험을 위하 여 추출된 RNA의 정량, 정성분석은 Fig. 1과 같다.
합성된 cDNA를 이용한 microarray 분석 결과, 대조군과 실 험군 사이에서 1.6배 이상 발현량이 차이가 나는 유전자는 총 520개로 사용된 chip에 있는 21927개 유전자 중에서 2.37%
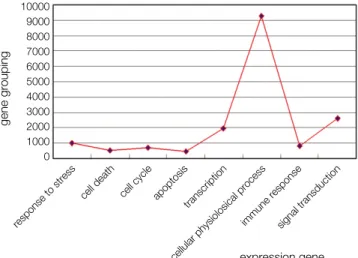
이었다. 건전치아군인 대조군에서는 143개의 유전자가, 우식치 아군인 실험군에서는 377개의 유전자가 1.6배이상 발현되었 다. Fig. 2는 microarray 결과를 M-A plot으로 표준화시켜 정 리한 것이다. Fig. 3은 사용된 모든 21927개 유전자들의 생물 학적 과정과 분자학적 기능에 관계된 gene grouping을 보여준 다. Fig. 4는 1.6배 이상 발현된 유전자들의 fold change를 그 래프로 보여준다.
2. 유전자 분석
Fig. 5, 6은 각 실험 결과를 NCBI의 Gene grouping tool을 이용하여 보다 더 기능적으로 관계가 되는 유전자끼리 분류하 였다.
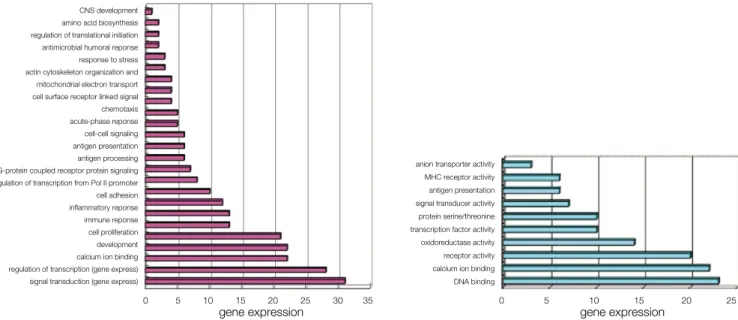
Fig. 5는 우식치아에서 발현이 증가된 유전자들의 생물학적 과정과 분자학적 기능적 분류로 생물학적 과정에서는 건전치아 에서 발현된 유전자와는 달리 면역반응과 염증반응, 항원작용, 항원제시 (antigen presentation)에 관계되었다.
분자학적 기능에서는 건전치아에서 발현된 유전자와는 달리 항원제시, MHC class I & II receptor activity에 관계되었다.
Fig. 6은 건전치아에서 발현이 증가된 유전자들의 생물학적 과정과 분자생물학적 기능적 분류로 생물학적 과정에서는 신호 전달, 전사의 조절, 대사, 세포 증식, 세포 유착등에 관계되는 유전자가 많았으며 분자생물학적 기능에서는 수용체 활성, 막 전위 수용체 활성, GTP 결합등에 관계되는 유전자가 많았다.
L1 (kb)
5.7
4.0
2.0
1.0 0.5 0.2
28S rRNA
18S rRNA
28S rRNA 18S rRNA
No Sample Ribosomal RNAratio(28s/18s)
L Ladder -
1Sample A 1.94
Fig. 1. RNA quality control report.
Fig. 2. M-A plot.
M -6 -4 -2 0 2 4 6
2 4 6 8 10 12 14 A
10000 9000 8000 7000 6000 5000 4000 3000 2000 1000 0
gene grouping
response to stress
cell death cell cycle apoptosis transcription
cellular physiolosical process
immune responsesignal transduction
expression gene Fig. 3. Gene grouping of used total genes.
250
200
150
100
50
0
frequency of fold change
'-2.5'~ '-2.0'~ '-1.6'~ -1.6'~ '2.0'~ '2.5'~ '3.0'~ '3.5'~
fold change range
Fig. 4. Histogram presenting frequency of fold change of the 1.6-fold or greater differentially expessed genes.
우식치아에서 발현이 증가된 선택된 유전자 중 nestin, Sp7 transcription factor, EGF-1, TGF-β1, bone-gamma-car- boxyglutamate protein(osteocalcin), FGF-R3, MHC com- plex I, TNF member 14, TGF-α는 RT-PCR 분석으로 다시 한번 확인하였다. 우식치아에서 발현이 감소된 선택된 유전자 중 phospholipase A2 group IVA, CD69 antigen, pyruvate dehydrogenase complex, secreted phosphoprotein 1, os- teomodulin은 RT-PCR 분석으로 다시 한번 확인하였다.
3. 유전자 발현 분석
cDNA microarray 결과를 확인하기 위해서 선택된 서로 다른 유전자로 역전사효소 중합효소 연쇄 반응 분석을 시행하였다.
유전자를 선택한 기준은 유전자 발현이 건전치의 치수 조직
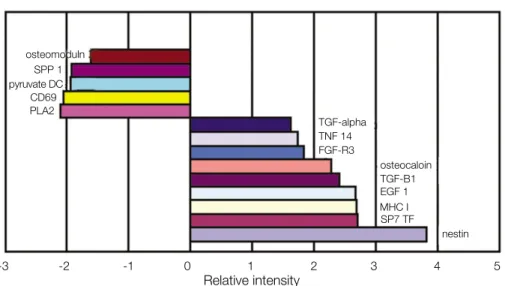
과 우식치의 치수 조직에서 cDNA microarray 분석법을 통해 보고된 적이 없고 발현 강도의 cut-off value가 1.6배 이상인 유전자를 선택하였다. Table 3은 선택된 유전자들의 발현 조직 및 기능을 분류하였고 Fig. 7은 선택된 유전자들의 상대적인 발 현 강도를 fold change를 기준으로 보여주었다.
RT-PCR 분석에서 tumor necrosis factor(ligand) super- family member 14, TGF-α, MHC class I, FGF-R3, bone- gamma-carboxyglutamate(osteocalcin), TGF-β1, EGF-1, Sp7 transcription factor, nestin이 우식 치아에서 증가되었 고 특히 MHC class I, TGF-β1, Sp7 transcription factor, nestin은 뚜렷한 증가를 보였다. 또한 phospholipase A2 group IVA, CD69 antigen, pyruvate dehydrogenase com- plex, secreted phosphoprotein 1, osteomodulin은 우식치아 에서 감소되었다(Fig. 8).
CNS development amino acid biosynthesis regulation of translational initiation antimicrobial humoral reponse response to stress actin cytoskeleton organization and mitochondrial electron transport cell surface receptor linked signal chemotaxis acute-phase reponse cell-cell signaling antigen presentation antigen processing G-protein coupled receptor protein signaling regulation of transcription from Pol II promoter cell adhesion inflammatory reponse immune reponse cell proliferation development calcium ion binding regulation of transcription (gene express) signal transduction (gene express)
0 5 10 15 20 25 30 35
gene expression
anion transporter activity MHC receptor activity antigen presentation signal transducer activity protein serine/threonine transcription factor activity oxidoreductase activity receptor activity calcium ion binding DNA binding
0 5 10 15 20 25
gene expression
Fig. 5. Biologic process (a) and molecular function (b) identified by the 377 genes more abundantly expressed in carious pulpal tissue.
protein kinse cascade protein transport heparin binding heparin bindingprotein biosynthesis ossification icosanoid metabolism cell surface receptor linked signal transduction growth factor activity cell proliferation metabolism regulation of transcription (gene expression) signal transduction (gene expression)
0 5 10 15 20
gene expression
GTPase activity structural molecule activity lathosterol oxidase activity transcription coactivator activity catalytic activity transferase activity phospholipid binding GTP binding transmembrane receptor activity receptor activity
0 5 10 15 20 25
gene expression Fig. 6. Biologic process (a) and molecular function (b) identified by the 143 genes more abundantly expressed in healthy pulpal tissue.
4. 면역조직화학적 관찰 결과
RT-PCR 분석 결과를 다시 한번 확인하기 위해서 우식치의 치수에서 발현이 증가된 TGF-β1을 면역조직화학적 관찰 하여 발현의 증가를 확인하였다. TGF-β1은 건전치에 비해 우식치의 조상아세포와 치수에서 특히 강하게 면역화학 염색이 발현되었 다(Table 4)(Fig. 9).
Table 3. The target and effects of selected genes in carious pulpal tissue Blood
Nerve immune growth cell phtsiologic gene
metabolism
vessel reponse &develop process expression
A.gene with up-regulation
Nestin o o o
SP7 TF o o
EGF 1 o o o o o
TGF-β1 o o o o o
osteocalcin o o
FGF-R3 o o o
TNF 14 o o o o
MHC I o o o
TGF-α o o o o
B.gene with down-regulation
PLA2 o o o
CD69 o o o o
pyruvate DC o o o
SPP 1 o o o
Osteomodulin o o o
Sp7 TF:Sp7 transcription factor, EGF 1:endothelial cell growth factor 1, TGF-β1:transforming growth factor beta 1, FGF-R3:fibroblast growth factor recep- tor 3, TNF 4:human genes tumor necrosis factor superfamily member 14, MHC I:major histocompatibility complex class I, TGF-α:transforming growth factor alpha, PLA2:phospholipase A2 group IVA, CD69:CD69 antigen, pyruvate DC:pyruvate dehydrogenase complex, SPP 1:secreted phosphoprotein 1.
Fig. 7. The differential expresson of selected genes in healthy and carious pulpal tissue.
differential expression of genes
osteomoduln SPP 1 pyruvate DC CD69 PLA2
TGF-alpha TNF 14 FGF-R3
osteocaloin TGF-B1 EGF 1 MHC I SP7 TF
nestin
Table 4. Labelling score of TGF-β1
samples healthy carious
ododtoblasts of pulp ++ +++
Dental pulp + ++
predentin + +
negative(-), weakly positive(+), positive(++), strongly positive(+++)
-3 -2 -1 0 1 2 3 4 5 Relative intensity
Fig. 8. The RT-PCR analysis was done selected genes.
(i) Up-regulaton genes: nestin, Sp7 transcription factor (Sp7 TF), transforming growth factor beta 1 (TGF-β1), endothelial cell growth factor 1 (EGF-1), osteocalcin, fibroblast growth factor receptor 3 (FGF-R3), major histocompatibility complex class I (MHC class I), transforming growth factor alpha (TGF-α), human genes tumor necrosis factor superfamily member 14 (TNF member 14).
(ii) Down-regulaton genes: Phospholipase A2 (PLA2) group IVA , CD69 antigen, pyruvate dehydrogenase complex (Pyruvate DC), secreted phosphoprotein 1 (SPP 1), osteomodulin.
Fig. 9. Immunohistochemistry with TGF-β1.
Sections stained for TGF-β1. Pulp of healthy teeth(a, c) and pulp of carious teeth(b, d). positive reactions are repre- sented by the brown staining. The dark dots present in this figure are the nuclei of the cells stained by Mayer`s haematoxylin : immunohistochem- istry, original magnification ×100(a, b), ×400(c), ×600(d).
(a)
(c)
(b)
(d)
H C H C H C H C H C
H C H C H C H C
H C H C H C H C H C
nestin Sp7 TF EGF-1 TGF-β1
FGF-R3 MHC classI TGF-α TNF-14
osteocalcin
PLA2 CD69 antigen pyruvate DC SPP 1 osteomodulin
Ⅳ. 총괄 및 고안
치아 우식증은 여전히 중요한 치과 질환이고 치아 우식증 하 방의 치수에서는 복합적인 여러 반응이 일어나게 되며 질병으 로부터 직접적 혹은 간접적으로 조직의 손상이 야기되어 여러 가지 방어 반응이 질병으로부터 일어난 손상과 치유를 위한 수 복과 재생을 목적으로 나타난다. 상아모세포는 고도의 분화과 정을 거치는 세포이며 치수의 바깥쪽에 위치하여 상아질을 만 든다. 상아모세포들은 일차, 이차, 삼차 상아질 생성에 관여하 여 수복 상아질(reparative dentin)이나 골양 상아질(osteo- dentin)을 생성하고 치수 손상후 수복 재생 과정(reparative process)에 관여한다10).
치아 우식증의 치료는 감염된 병소를 제거하고 치수를 보호 한다. 인공적인 재료로 이를 수복하는 여러 과정이 관여하고 있 으며 미래의 치료는 우식 질환동안 발생하는 분자 생물학적 이 해에 기초한 유전자(gene) 치료, 줄기 세포(stem cell) 치료, 성장 인자(growth factor) 치료 등의 생물학적 접근이 기대된 다. 유전자 치료는 치아의 치유 과정을 생물학적으로 조절하는 미래의 치료 방향이며 줄기 세포 치료 또한 생활 치수 치료에서 미래에 가장 주목받는 치료가 될 것이다. 치아의 손상을 치유할 수 있는 적합한 줄기 세포의 보존은 치아 손상의 문제를 해결할 수 있는 중요한 방법 중의 하나가 될 것이다. 조직의 치유과정 을 매개하는 성장인자의 사용도 상아질 치수 복합체의 치유를 유도하는 데에 사용될 수 있을 것이다5). 본 연구에서 cDNA microarray를 이용한 광범위한 유전자 분석은 이러한 앞으로 의 치료 방향을 제시하는 데에 도움을 줄 것으로 사료된다.
치수 조직은 대부분 결합 조직으로 구성되어 있으나 혈관과 신경섬유를 포함하는 이종(heterogenous) 세포들, 그리고 질 환의 진행과정에서 중추적 역할을 담당하는 세포들 사이의 상 호작용으로 인하여 끊임없이 변화하는 세포들로 구성되어 있 다. 이러한 실험군의 한계에도 불구하고 우리는 microarray 실 험 결과 우식치아와 건전치아의 치수 조직에서 여러 유전자들 의 다른 발현을 볼 수 있었다. 또한 그 중 아직까지 우식치아의 치수에서의 발현이 알려지지 않은 것들도 있었다.
대사 경로(pathway)나 기전과 관련된 의미있는 생물학적 정 보로 유전자 발현을 해석하는 일은 미래의 유전자 분야에서 도 전되는 과제이다. 본 연구에서 유전자의 기능적 분류는 치아 우 식을 조절하는 분자생물학적 기능을 규명하고 있으며 앞으로의 연구에 중요한 기초가 될 것이다.
저자는 cDNA microarray 결과를 한번 더 확인하기 위해서 역전사 효소 중합효소 연쇄반응 분석을 시행하였다. RT-PCR 분석에서 증가 혹은 감소된 유전자 중 명백하게 큰 차이를 보이 는 것도 있었으나 그 차이가 작은 정도를 보이는 것도 있었던 것은 우리가 실험에 사용한 치아들은 법랑질에서 상아질까지 우식이 진행된 1급에서 2급의 병소, 즉 병소의 크기가 작은 치 아만을 사용하여서 상대적으로 손상에 영향을 받지 않은 치수 조직이 많았기 때문인 것으로 사료된다. 또한 본 실험 결과에서 는 상대적으로 염증 과정 및 면역 체계에 관여하는 세포나 유전
자가 적었다. 앞으로의 실험에서는 우식이 깊게 진행된 치아에 서의 유전자 발현을 분석해 보는 것도 의미있는 실험이 될 것으 로 사료된다.
우식치아에서 정상보다 증가되었거나 감소된 유전자들은 여 러 가지 분자생물학적 기능적 분류에 속하고 있다.
우식치아에서 증가된 유전자 중 nestin은 중간 섬유로서 신 경계와 근육의 발생에 주로 관여한다11). 치아 발생의 상아질 형 성 과정 동안 nestin은 조상아세포와 교두 부위의 치수 섬유모 세포에서 발견되었다12). Imad A 등13)은 Nestin이 우식 병소 주의의 상아모세포 돌기에서 발현되었다고 보고하였는데 이는 치수가 우식 과정에 대한 방어기전으로 상아세관이 점진적으로 석회화되고 상아모세포의 생활력을 유지하는 기전에서 nestin 이 수복 상아질의 완성에 중요한 역할을 담당하고 있는 것으로 볼 수 있으며 본 실험에서도 이를 확인할 수 있었다. nestin은 우식에 대한 수복 과정에 관여하고 있으며 치아 손상 후 상아모 세포의 상아질 분비 활성에 관여하는 것으로 볼 수 있다.
우식 병소와 같은 병적 과정에서 상아모세포의 활성으로 수 복 상아질이 분비되는데 이러한 자극은 여러 가지 성장 인자들 이 상아질로부터 유리되어 신호 전달 물질로 작용하기 때문이 다14). 우식 탈회 과정동안 상아 기질로부터 유리된 성장인자들 은 수복 상아질을 만드는 세포 분비 활성을 증가시키는 신호 전 달에 매우 중요한 역할을 한다15).
이러한 성장인자 중 대표적인 것으로 transforming growth factor-beta(TGF-β)를 들 수 있으며 본 실험에서도 TGF-β1 이 우식 병소에서 증가되었고 cDNA microarray를 통해 분자 생물학적으로도 증명하였다.
TGF-β1은 초기 뇌상기의 치성 상피에서 발견된 이후 상아모 세포, 분화중인 치수세포등에 영향을 주어 일차 상아질과 이차 상아질 형성을 유도한다고 알려져 있다16). Sloan AJ 등17)은 TGF-β1을 노출되지 않은 치수의 조상모세포에 적용하여 수복 상아질을 형성하는 신호전달 능력을 보여 주었고 McLachlan JL 등18)은 RT-PCR을 통해 TGF-β1이 우식치아의 상아모세포 에서 증가됨을 보여주었다. 이러한 발견들은 어떻게 자연적인 재생 과정을 치아 보존에 사용할 수 있을지 뿐만 아니라 치아의 재생 및 회복에 있어 생물학적으로 작용하여 치료할 수 있는 가 능성을 제공한다.
Fibroblast growth factor receptor 3(FGF-R3)는 네개의 다른 membrane spanning tyrosine kinases로 구성되었으며 fibroblast growth factors에 높은 친화력을 지닌 수용체로 작 용하고 골격의 발생에 필수적 역할을 한다19). Nakajima 등은 골절의 치유시 FGF-R3가 연골 형성을 조절하고 연골내 골화 를 증진시켜 골절의 치유에 기여한다고 보고하였다20). 아직까지 FGF-R3가 치아에서 연구된 적은 없었으나 FGFs는 치배 형성 과정 중 개시기, 형성기, 분화기에 관여함이 밝혀졌다21).
여러 연구에서 TGF-β와 FGFs가 notch signalling을 조절 함이 밝혀 졌고21), notch signalling은 우식 병소에 의해 유도 되어 조직 치유가 분자생물학적으로 일어나는 초기에 증가한 다. 수복 상아질을 형성하는 데에 있어서 TGF-β의 효과는 손상
받은 치수에서 notch signalling pathway의 발현을 조절한다22). 본 연구에서 우식치아에서 증가된 FGF-R3 역시 FGFs와 비슷 한 역할을 할 것으로 사료되나 이에 대한 더 많은 연구가 있어 야 할 것이다.
Endotherial cell growth factor-1(ECGF-1)도 우식치아에 서 증가하였으며 angiogenic growth factor로서 잘 알려져 있 다. Angiogenesis는 모든 조직에서 상처 치유과정에 주요한 역 할을 담당하고 있고 pulpal angiogenesis는 치수의 성공적인 치유를 위한 필요조건이다23). 치수의 치유과정에서 subodon- toblastic capillary plexus의 재혈관화가 삼차 상아질(수복 상 아질)의 반응에 필요하다24). Begue-Kirn C 등25)은 angoigenic growth factors는 상아질 회복에 관계되는 생리적인 과정과 odontobalst-like cell의 새로운 세대로의 분화와 관련된 신호 전달에 중요한 역할을 한다고 하였으며 본 실험에서도 ECGF- 1은 그러한 역할을 한다고 사료된다.
sp7 transcription gene은 osterix gene으로도 알려져 있으 며 조골세포와 골형성에 관여한다. Nakashima 등26)은 osterix gene이 결여된 흰쥐에서 골 생성이 일어나지 않음을 보고하였 다. sp7 transcription gene은 osteocalcin, osteonection, osteopontin, bone sialoprotein과 같은 수많은 중요한 조골세 포 유전자의 발현을 조절한다. sp7 transcription gene은 bone morphogenic protein-2(BMP-2)로 자극된 murine pluripotent myoblast C2C12 세포로부터 유도되는 것이 처 음 증명되었고 동시에 골수 기질 세포와 연골세포에서도 증가 되었다. 또한 sp7 transcription gene의 발현의 조절은 골치유 에 유용함이 보고되었으나27)아직까지 sp7 transcription gene 이 치아 연관세포에서 발견된 적은 없었다.
우식치아에서 증가된 유전자인 osteocalcin은 K+-depen- dent이고 비콜라겐성 상아질과 골 기질을 형성하는 단백질이 다. 조골세포와 상아모세포에 의해 합성되며 그러한 세포들의 생활력 및 분화와 조골 능력을 볼 수 있는 잘 알려진 표식인자 이다28). Owen 등29)은 osteocalcin이 골 형성과 조골세포의 분 화에 중요한 역할을 하며 조골세포의 가장 마지막 분화단계에 서 발현한다고 보고하였다. Osteocalcin과 치아 우식과의 관계 는 아직 밝혀진 것이 없으나 Hirata 등30)은 형태적 변형을 동반 한 치아 삭제 후 수복 상아질 형성 초기 단계에 발현되는 것을 관찰하였으며 특히 RT-PCR 분석에서 치아 삭제 후 2-3일 안 에 발현되었다고 보고하였다. 본 연구에서는 건전치아에 비해 우식치아에서 osteocalcin이 더 많이 발현되는 것을 관찰하였 고 이는 osteocalcin의 발현이 수복 상아질의 형성과 관계가 있 으며 외부 자극원에 대해 치수의 생활력을 유지하기 위해 방어 하는 작용을 도와주는 것으로 사료된다.
우식치아에서 증가된 유전자인 major histocompatibility complex는 두 개의 항원군인 class I과 class II로 분류되며 class II는 세포막 결합 glycoprotein인 HLA-DP와 -DQ와 - DR로 구성된다. 대부분의 면역 반응은 class II 유전자를 가진 세포에 의해 T 세포에서 항원의 발현으로 시작한다31). 치수가 손상에 반응하여 방어 기전을 일으킬 때 염증 반응도 시작하게
되며 우식이 점점 진행됨에 따라 염증 반응은 심해지고 미생물 이 치수로 파급되는 급성 상태가 되어 심한 경우 치수 괴사나 치근 병소로 발전한다. Kamal 등32)은 항원제시(antigen pre- sentation) 세포와 대식 세포 관련 항원제시 세포가 초기 우식 증에서 초기 치수 반응 동안 관찰되었음을 보고하였다.
MHC class I과 class II는 치수에 분포되어 있고 우식 병소 아래에서 발견되며 외부의 자극에 대해서 면역 반응을 개시하 는 항원 제시 세포이다33,50). 면역조직화학적 연구에서 건전치아 도 MHC class I과 class II 세포들이 관찰되었으나 치수염이 진행되었을 때에는 이 세포들의 수가 증가함이 보고되었다34). 치수에서 MHC class II는 초기 방어 반응에 참여하고 노출된 상아세관을 통해 오는 외부 자극에 대한 생물학적 감지장치로 작용한다35).
우식치아에서 증가된 유전자인 tumor necrosis factor(lig- and) superfamily, member 14(TNF superfamily, member14) 는 면역체계의 생체 항상성의 유지에 중요하게 관여하고 이 체 계의 생물학적인 기능은 염증작용과 숙주 방어 작용에서 보호 효과가 있으며 감염에 대한 숙주 반응을 구성하는 세포 반응을 개시하고 참여한다36). 또한 TNF는 중요한 염증 매개체이며 성 장을 자극하고 분자의 결합을 발현시키며 활성화시키고 cy- tokine의 생성을 유도하여 면역반응을 시작하는 중심에 절대적 역할을 한다37). Cytokine과 chemokine은 세균 감염에 대한 숙 주 반응으로 연구되고 있고 대다수의 면역 세포에 의해 표현되 며 목표가 되는 세포에 생물학적 효과를 가하여 면역체계 반응 을 매개하는 중요한 역할을 한다. TNF-α, IL-1β, IL-6, IL-8 과 같은 cytokine은 주로 염증 전 반응과 국소적 및 전신적 염 증반응의 양쪽을 매개한다38). McLachlan JL 등39)은 10개의 건전 치아와 10개의 우식 치아의 치수에 대한 비교연구에서 TNF-α, IL-1β, IL-6, IL-8가 우식 치아에서 더 많이 발현된다 고 보고하였고 본 연구에서는 TNF superfamily, member 14 가 우식치아에서 더 많이 발현되었다.
우식치아에서 증가된 유전자인 transforming growth fac- tor, alpha(TGF-α)40)는 epidermal growth factor와 같은 성 장 인자 군이며 같은 세포 표면 수용체이다41). Twardzik42)는 TGF-α가 흰쥐의 배아에서 7일째 가장 많이 발현되고 이후 점 점 줄어든다고 보고 하였고 Wilcox과 Derynck43)는 흰쥐의 배 아에서 첫 번째와 두 번째 branchial arch에서 발견되었다고 보고하였다. Kronmiller 등44)은 TGF-α는 하악의 발생시 치판 의 형성에서 치아형성을 개시하는 중요한 역할을 하고 있다고 보고하였다.
우식치아에서 감소된 유전자인 phospholipase A2(PLA2), group IVA는 lysophospholipid와 유리지방산을 만들기 위해 phospholipids의 sn-2 위치에서 에스테르 결합의 가수분해를 촉매하는 효소이다45). PLA2는 중요한 지질 매개체의 생성에 관여하고 여러 생리적인 그리고 병적인 환경에서 phospho- lipids의 항상성에 관여한다46). 또한 PLA2의 활성은 arachi- donic acid를 유리하는데 작용하고 그 후 prostaglandin을 생 성하게 되는데 prostaglandin은 조직 손상이나 염증 결과, 통
각 과민(hyperalgesia)의 발생 에 중요한 역할을 한다47). 우식치아에서 감소된 유전자인 CD69 antigen(p60, early T-cell activation antigen)는 CD69 antigen은 염증의 음성 조절자(negative modulator)로서 이는 TGF-β의 합성을 통해 조절하고 그로 인해 여러 가지 염증 전 산물을 감소시킨다고 하 였고 CD69는 면역 세포 활성과 직접적으로 또는 간접적으로 염증전 효과에 관여한다고 보고하였다48).
우식치아에서 감소된 유전자인 secreted phosphoprotein 1 (osteopontin, bone sialoprotein 1)은 골에서 주요한 비콜라 겐성 단백질(major noncollagenous protein)인 bone sialo- protein이며49) bone sialoprotein은 골, 광화된 연조직, 상아 질, 백악질을 포함한 광화된 조직에서 높게 발현된다50). 최근의 연구에서 법랑모세포에서도 발현이 보고되었고51) Garcia 등은 bone sialoprotein 1이 맹출 중인 치아뿐만 아니라 치배의 상 아모세포에서 면역화학염색시 약하게 염색된 것을 관찰하였고 이것은 상아질 기질의 생성과 광화에서 세포 전달과 관련이 있 음을 보고하였다52). 본 연구에서 건전치아에 비해 우식치아에서 감소된 것도 상아질의 광화와 관련이 있다고 사료된다.
우식치아에서 감소된 유전자인 pyruvate dehydrogenase complex, component X는 acetyl-CoA를 형성하는 pyruvate 의 비가역적인 산화형탈탄산화를 촉매하는 미토콘드리아 내 다 효소 복합체이며 신경의 발생에 관여하며 부족시 신경학적 이 상을 초래할 수 있다53).
우식치아에서 감소된 유전자인 osteomodulin에 대하여 O'Reganx 등54)은 osteomodulin이 치배단계에서는 일차 상아 질 분비와 관련된 치수의 끝부분에 배열되어 있는 상아모세포 에서 발현이 관찰되었고 초기 치관 형성시에 상아모세포에서 발견되고 새로 형성된 mantle dentin에서 많이 나타났으며 상 아질 기질 주위의 성숙한 상아모세포에서 높은 발현으로 보아 치아의 발생과 상아질의 광화에 중요한 역할을 담당한다고 보 고하였다. 본 연구에서 건전치아에 비해 우식치아에서 감소된 것도 secreted phosphoprotein 1과 같이 상아질의 광화와 관 련이 있다고 사료된다.
현재 치수 치료와 수복 치료시 이러한 유전자와 성장인자에 기초한 약제는 없으나 앞으로 이러한 유전자들의 더 많은 발현 양상에 대한 연구와 세포에서의 발현 연구, 임상적 사용을 위한 연구를 통하여 생리적인 수복 재생 과정에 기초한 치관 부위 병 소 수복을 대신하고 근관 치료에 사용될 수 있는 약제가 소개될 수 있을 것이다.
cDNA microarray를 이용하여 한번에 많은 양의 유전자 발 현을 분석하는 방법으로 우식의 정도, 남아있는 법랑질 및 상아 질의 두께, 환자의 나이, 세균 감염등의 여러 중요한 요소들을 고려하여 초기 단계부터 우식 질환에 관계된 분자 생물학적 회 로를 종합적으로 이해하고 목표가 되는 치료방법에 보다 쉽게 접근할 수 있게 되었다. 본 연구는 전 세계에서 가장 널리 퍼져 있는 만성질환 중 하나인 치아 우식 병소에서 일어나는 재생 회 복적인 과정과 염증 반응, 방어 반응들 사이의 복잡한 상호작용 을 분자 생물학적으로 규명하였다. 덧붙여 본 연구의 새로운 발
견은 염증 반응을 억제하고 치아의 생활력을 유지하며 필수적 으로 방어 반응과 재생 회복 반응이 일어나는 데에 목표를 두는 새로운 치료 방법의 연구에 기초가 될 수 있을 것이다.
Ⅴ. 결 론
치아 우식증은 범발성 질환이고 이에 대한 생체반응은 단순 하지 않으며 질병 과정과 숙주의 활성 모두를 반영하는 복합적 인 반응이다. 이러한 반응을 이해하기 위해서는 질병의 세포학 적, 분자학적인 면을 이해하는 것이 필수적이다.
이에 본 연구는 임상적으로 건강한 치아와 우식이 진행된 치 아로부터 얻어진 치수 안의 유전자 발현을 규명하고 우식 병소 에서 일어나는 치유 및 재생에 관계되는 분자와 면역 세포들 사 이의 분자 생화학적 상호작용을 규명하기 위해서 우식치아와 건전치아의 치수를 이용하여 cDNA 미세배열(microarray) 분 석과 역전사효소 중합효소 연쇄반응(RT-PCR) 분석, 그리고 면역화학염색법(immunohisto-chemistry)을 시행하여 다음과 같은 결론을 얻었다.
1. cDNA 미세배열 분석 결과, 건전치아군인 대조군에서는 143개의 유전자가, 우식치아군인 실험군에서는 377개의 유전자가 1.6배이상 발현되었다.
2. 역전사효소 중합효소 연쇄반응 분석에서 14개의 유전자를 선택하였고 cDNA 미세배열 분석결과와 동일한 결과를 확 인하였다.
3. TGF-β1의 면역조직화학적 관찰 결과, 건전치에 비해 우 식치의 상아모세포와 치수에서 특히 강하게 발현되었다.
참고문헌
1. Abiko Y : Passive immunization against dental caries and periodontal disease: development of recombinant and human monoclonal antibodies. Crit Rev Oral Biol Med, 11:140-158, 2000.
2. Smith AJ, Murray PE, Lumley PJ : Preserving the vital pulp in operative dentistry 1. A biological approach. Dent Update, 29:64-69, 2002.
3. Smith AJ, Murray PE, Lumley PJ : Preserving the vital pulp in operative dentistry: 4. Factors influenc- ing successful pulp capping. Dent Update, 29:225- 230, 2002.
4. Smith AJ : Pulpal responses to caries and dental repair. Caries Res, 36:223-232, 2002.
5. Lockhart DJ, Winzele REA : Genomics, gene expres- sion and DNA arrays. Nature, 405:827-836, 2000.
6. Brown PO, Botstein D : Exploring the new world of the genome with DNA microarrays. Nat Genet, 21:33-37, 1999.
7. Debouck C, Goodfellow PN : DNA microarrays in
drug discovery and development. Nat Genet, 21:48- 50; 1999.
8. Paakkonen V, Ohlmeier S, Bergmann U, et al. : Analysis of gene and protein expression in healthy and carious tooth pulp with cDNA microarray and two-dimensional gel electrophoresis. Eur J Oral Sci, 113:369-379, 2005.
9. McLachlan JL, Smith AJ, Bujalska IJ, et al. : Gene expression profiling of pulpal tissue reveals the mol- ecular complexity of dental caries. Biochim Biophys Acta. 25:1741:271-281, 2005.
10. Lesot H, Begue-Kirn C, Kubler MD, et al. : Experimental induction of odontoblast differentiation and stimulation during reparative processes. Cell Mater, 3:211-217, 1993.
11. Lendahl U, Zimmerman LB, McKay RDG : CNS stem cells express a new class of intermediate fila- ment protein. Cell, 60: 585-595, 1990.
12. About I, Maquin D, Lendahl U, et al. : Nestin expression in embryonic and adult human teeth under normal and pathological conditions. Am J Pathol, 157:287-295, 2000.
13. Imad A, Dominique LM, Urban L, et al. : Nestin Expression in Embryonic and Adult Human Teeth under Normal and Pathological Conditions.
American J Patho, 157:1, 2000.
14. Tziafas D, Smith AJ, Lesot H : Designing new treatment strategies in vital pulp therapy. Dent, 28:77-92, 2000.
15. Bj�rndal L, Darvann T : A light microscopic study of odontoblastic and non-odontoblastic cells involved in tertiary dentinogenesis in welldefined cavitated cari- ous lesions. Caries Res, 33:50-60, 1999.
16. Melin M, Joffre-Romeas A, Farges JC, et al. : Effects of TGF-β1 on dental pulp cells in cultured human tooth slices. J Dent Res, 79:1689-1696, 2000.
17. Sloan AJ, Smith AJ : Stimulation of the dentine- pulp complex of rat incisor teeth by transforming growth factorbeta isoforms 1-3 in vitro. Arch Oral Biol, 44:149-156, 1999.
18. McLachlan JL, Smith AJ, Sloan AJ : Cooper PR.Gene expression analysis in cells of the dentine- pulp complex in ealthy and carious teeth. Arch Oral Biol, 48:273-283, 2003.
19. Peters K, Werner S, Chen G, et al. : Two FGF receptor genes are differentially expressed in epithe- lial and mesenchymal tissues during limb formation and organogenesis in the mouse. Development,
114:233-243, 1992.
20. Nakajima A, Shimizu S, Moriya H, et al. : Expression of fibroblast growth factor receptor-3 (FGFR3), signal transducer and activator of tran- scription-1, and cyclin-dependent kinase inhibitor p21 during endochondral ossification: differential role of FGFR3 in skeletal development and fracture repair. Endocrinology, 144:4659-4668, 2003.
21. Mitsiadis TA, Henrique D, Thesleff I, et al. : Mouse Serrate-1(Jagged-1): Expression in the developing tooth is regulated by epithelial-mesenchymal inter- actions and fibroblast growth factor-4. Development, 124:1473-1483, 1997.
22. Mitsiadis TA, Romeas A, Lendahl U, et al. : Notch2 protein distribution in human teeth under normal and pathological conditions. Exp Cell Res, 282:101- 109, 2003.
23. Baume, LJ : The biology of pulp and dentine. A his- toric, terminologic-taxonomic, histologic-biochemical, embryonic and clinical survey. Monogr Oral Sci, 8:210-220, 1980.
24. Schroder, U : Effects of calcium hydroxide-contain- ing pulp-capping agents on pulp cell migration, pro- liferation and differentiation. J Dent Res, 64:541- 548, 1985.
25. Begue-Kirn C, Smith AJ, Loriot M, et al. : Comparative analysis of TGFbs, BMPs, IGFs, Msxs, fibronectin, osteonectin and bone sialoprotein gene expressions during normal and in vitro induced odontoblast differentiation. Int J Dev Biol, 38:405- 420, 1994.
26. Nakashima K, Zhou X, Kunkel G, et al. : The novel zinc finger-containing transcription factor osterix is required for osteoblast differentiation and bone for- mation. Cell, 108:17-29, 2002.
27. Zhang X, Schwarz EM, Young DA, et al. : Cyclooxy- genase -2 regulates mesenchymal cell differentiation into the osteoblast lineage and is critically involved in bone repair. J Clin Invest, 109:1405-1415, 2002.
28. Camarda AJ, Butler WT, Finkelman RD, et al. : Immuno-cytochemical localization of gamma-car- boxyglutamic acid-containing proteins (osteocalcin) in rat bone and dentin. Calcified Tissue International 40:349-355, 1987.
29. Owen TA, Bortell R, Yocum SA, et al. : Coordinate occupancy of AP-1 sites in the vitamin D responsive and CCAAT box elements by Fos-Jun in the osteocalcin gene: model for phenotype suppression of transcription.
Proc Nat Acad Sci USA, 87:9990-9994, 1990.
30. Hirata M, Yamaza T, Mei YF, et al. : Expression of osteocalcin and Jun D in the early period during reactionary dentin formation after tooth preparation in rat molars. Cell tissue Res 319:455-465, 2005.
31. Yoshiba N, Yoshiba K, Nakamura H, et al. : Immunohistochemical localization of HLA-DR-posi- tive cells in unerupted and erupted normal and cari- ous human teeth. J Dent Res, 75:1585-1589, 1996.
32. Kamal AMM, Okiji T, Kawashima N, et al : Defense responses of dentin/pulp complex to experimentally- induced caries in rat molars: An immunohistochemi- cal study on kinetics of pulpal 1a antigen -express- ing cells and macrophages. J Endod, 23:115-120, 1997.
33. Ohshima H, Sato O, Kawahara I, et al. : Responses of immunocompetent cells to cavity preparation in rat molars: an immunohistochemical study using OX6-monoclonal antibody. Connect Tissue Res, 32:303-311, 1995.
34. Bergenholtz G, Nagaoka S, Jontell M : Class II anti- gen expressing cells in experimentally induced pul- pitis. Int Endod J, 24:8, 1991.
35. Ohshima H, Sato O, Kawahara, et al. : Responses of immunocompetent cells to cavity preparation in rat molars: an immunohistochemical study using OX6-monoclonal antibody. Connect Tissue Res, 32:303-311, 1995.
36. Dinarello, CA : Biologic basis for interleukin-1 in disease. Blood, 87:2095-2147, 1996.
37. Thomas H, Klaus P : The intriguing biology of the tumour necrosis factor/tumour necrosis factor recep- tor superfamily: players, rules and the games.
Immunol, 115:1-20. 2005.
38. Nathan C : Points of control in inflammation.
Nature, 420: 846-852, 2002
39. McLachlan JL, Sloan AJ, Smith AJ, et al : S100 and cytokine expression in caries Infect Immun, 72:4102-4108, Jul 2004.
40. DeLarco JE, Todaro GJ : Growth factors from murine sarcoma virus transformed cells. Proc natn Acad Sci USA, 75:4001-4005,1978.
41. Partanen AM, Ekblom P, Thesleff I : Epidermal growth factor inhibits morphogenesis and cell differ- entiation in cultured mouse embryonic teeth. Dev Biol, 111:84-94, Sep 1985.
42. Twardzik DR : Differential expression of transform- ing growth factor-alpha during prenatal development of the mouse. Cancer Res, 45:5413-5416, 1985.
43. Wilcox JN, Derynck R : Developmental expression of transforming growth factors alpha and beta in mouse fetus. Molec Cell Biol, 8: 3415-3422, 1988.
44. Kronmiller JE, Upholt WB, Kollare J : Effects of retinol on the temporal expression of transforming growth factor-a mRNA in the embryonic mouse mandible. Archs Oral Biol, 38:185-188, 1993.
45. Wang Y, Kowalski PE, Thalmann I, et al. : Otoconin- 90, the mammalian otoconial matrix protein, con- tains two domains of homology to secretory phospho- lipase A2. Proc Natl Acad, 95:15345-15350, 1998.
46. Diaz BL, Arm JP : Phospholipase A2, Prostaglandins, Leukot. Essent Fat Acids, 69:87-97, 2003.
47. Kujubu DA, Fletcher BS, Varnum BC, Lim RW, et al. : TIS10, a phorbol ester tumor promoter- inducible mRNA from Swiss 3T3 cells, encodes a novel prostaglandin synthase/cyclooxygenase homo- logue. J Biol Chem, 266:12866-12872, 1991.
48. Sancho D, Gomez M, Viedma F, Esplugues E, Gordon-Alonso M, Garcia-Lopez MA, de la Fuente H, Martinez-A C, Lauzurica P, Sanchez-Madrid F.CD69 downregulates autoimmune reactivity through active transforming growth factor-beta pro- duction in collagen-induced arthritis. J Clin Invest, 112:872-882, 2003.
49. Buchaille ML, Couble H, Magloire, et al. : Expression of the Small Leucine-Rich Proteoglycan Osteoadherin/ Osteomodulin in Human Dental Pulp and Developing Rat Teeth R. Bone, 27:265-270, 2000.
50. Oldberg A, Franze NA, Heinegard D : The primary structure of a cell-binding bone sialoprotein. J Biol Chem, 263:19430-19432, 1988.
51. ChenJ, Sasaguri K, Sodek J, et al. : Enamel epithe- lium expresses bone sialoprotein (BSP).
EuropeanJournal of Oral Sciences, 106: 331-336, 1998.
52. Garcia1 JMQ, Martins MD, Jaeger RG, et al. : immunolocalization of bone extracellular matrix pro- teins(type I collagen, osteonectin and bone sialopro- tein) in human dental pulp and cultured pulp cells.
International Endodontic Journal, 36:404-410, 2003.
53. Mark TJ, Saleh M, Susannah LH : Inactivation of the Murine Pyruvate Dehydrogenase (Pdha1) Gene and Its Effect on Early Embryonic Development. Mol Gen and Met, 74:293-302, 2001.
54. O'Regan AW, Nau GJ, Chupp GL, et al : Osteopontin (Eta-1) in cell-mediated immunity:
teaching an old dog new tricks. Immunol Today, 21:475-478, 2000.
Key words: Caries, cDNA microarray analysis, RT-PCR analysis, Immunohistochemistry
GENE EXPRESSION ANALYSIS OF THE DENTAL PULP IN HEALTHY AND CARIES TEETH
So-Hee Oh*, Jong-Soo Kim**
*Hallym Sacred Heart Hospital, **Department of Pediatric Dentistry, School of Dentistry, Dankook University
Deep caries may induce pulpitis and the pulpal tissue interacts with microbial invasion. The immune response to protect the pulpal tissue can be mediated by cellular signal molecules produced by the pulpal cells. The un- derstanding of these processes is important to find future therapeutic method for the diseased pulp.
The pulp tissue from sound teeth was set as control group (n=30) and the pulp tissue from decayed teeth was set as test group (n=30). Total RNA was extracted from the pulp of each group and it was used for cDNA microarray and reverse transcriptase-polymerase chain reaction(RT-PCR). The expression of TGF-β1 was stud- ied by immunohistochemistry.
The results were as follows:
1. cDNA microarray analysis identified 520 genes with 6-fold or greater difference in expression level with 143 genes more abundant in health and 377 genes more abundant in disease.
2. The RT-PCR analysis was done for randomly selected 14 genes and the results supported the result of cDNA microarray assay.
3. TGF-β1 was highly expressed in the carious pulp and it was found in odontoblast by immunohistochem- istry.
In conclusion, many cytokines were found to be significantly changed their expression in the diseased pulp(/M/>1.6).
Abstract