Bacillus subtilis 분리균주들의 독소생성 진균에 대한 항진균 활성
황지선 ・ 송홍규 * 강원대학교 생명과학과
Antifungal activity of Bacillus subtilis isolates against toxigenic fungi
Ji-Seon Hwang and Hong-Gyu Song *
Department of Biological Sciences, Kangwon National University, Chuncheon 24341, Republic of Korea
(Received February 13, 2020; Revised February 26, 2020; Accepted March 16, 2020)
*For correspondence. E-mail: [email protected];
Tel.: +82-33-250-8545; Fax: +82-33-259-5665
Toxigenic fungi can contaminate feed and grains and produce toxins which are toxic and carcinogenic to humans and animals at low concentration. This study aims to evaluate antifungal activity against toxigenic fungi by Bacillus subtilis AF11 and YR47 isolated from soil and their antifungal substances. These strains suppressed mycelial growth in the plate diffusion assay and showed the inhibition zones over 13.3 mm against 3 ochratoxigenic fugi and 4 fumonisinigenic fungi. AF11 and YR47 also inhibited sporulation of the target fungi by 77.7~
100.0 and 75.4~100.0%, respectively compared to those of the control. They removed 38~90% of the fungal spores (10
7spores/ml), and suppressed germination of the remaining spores by more than 94.9%. Ochratoxin A production decreased effec- tively when ochratoxigenic Aspergillus strains were cocultured with AF11 or YR47 in YES broth. It might be due to the antifungal substances including siderophore, β-1,3-glucanase, chitinase and protease produced by these strains. These results suggest that Bacillus subtilis AF11 and YR47 can be used to control growth of toxigenic fungi and their mycotoxin pro- duction.
Keywords: Bacillus subtilis, antifungal activity, antifungal substances, toxigenic fungi
진균은 식물의 생장 또는 저장 중에 발생하는데 이중 독 소를 생성하는 종류들은 특히 곡물이나 견과류 등을 오염
시켜 이를 섭취한 인간과 동물에게 큰 피해를 미치고 있다.
진균독소는 주로 Aspergillus, Penicillium 및 Fusarium과 같은 사상성 진균에 의해 생성되는데, 400개 이상의 진균 독소 중 주된 독소는 aflatoxins, ochratoxins, fumonisins, zearalenone 및 patulin 등이며(Richard, 2007), 이 중 ochra- toxin A와 fumonisin B1은 국제암연구소에 의해 인체발암 가능물질 2급으로 분류되었다(IARC, 1993). Ochratoxin A (OTA)는 사료곡물 및 커피 등에 오염 발생이 높으며 신장 독성, 기형유발, 면역 억제, 유전독성 및 신경독성을 나타내 며(Fazekas et al., 2002), Fumonisin B1 (FB1)은 곡물에서 오 염빈도가 가장 높고 독성이 강하다(Gelderblom et al., 2001).
우리나라에서도 저장 옥수수에서 Fusarium 속 독소가 다 량 검출되었으며(Kim et al., 2017), 대부분의 양돈사료가 fumonisins, deoxynivalenol 및 zearalenone 등의 진균독소 에 복합적으로 오염되었다고 보고된 바 있다(Chang, 2015).
이런 진균독소와 독소생성 진균으로 인한 공중 보건적 및 경제적인 피해 등으로 전 세계 국가들은 진균독소의 허용 기준을 정해놓고 있다(IARC, 1993).
진균독소 농도를 허용 기준 이하로 제어하기 위한 근본 적인 방법은 독소생성 진균의 생장을 억제하여 진균독소 를 분비하지 못하게 하는 것이다(Xiong et al., 2017). 그 중 항균성 미생물을 이용한 생물학적 제어는 물리화학적 제 어보다 환경 친화적이라는 장점이 있다(Wu et al., 2009).
이로 인해 생물학적 방제에 대한 관심이 높아지고 있는데,
다양한 미생물 중 Bacillus 속 미생물은 항세균 및 항진균 활성을 갖는 다양한 대사산물을 합성하여 일반적인 진균 및 독소생성 진균 억제에 대해 많은 연구가 수행되었다 (McKeen et al., 1986; Leifert et al., 1995; Zhang et al., 2008). 그러나 여러 진균이 동시에 오염되어 다른 종류의 독소를 생성할 수 있으므로 본 연구에서는 OTA와 FB1를 생산하는 진균들을 모두 억제하는 세균을 분리하고 이들 의 항진균 활성 및 항진균 물질을 조사하여 새로운 생물학 적 방제제로서의 가능성을 평가하였다.
재료 및 방법
세균 균주의 선별 및 동정
여러 지역에서 수집한 토양시료 5 g을 0.85% NaCl 용액 30 ml에 넣고 잘 혼합하고 10-4로 연속 희석하여 nutrient agar (NA, Difco Lab.)에 100 μl를 도말 후 배양하여 순수한 세균 집락을 분리하였다. 분리한 균주는 아래에 설명한 plate diffusion assay를 이용하여 다음의 독소생성 진균 모 두에 대해 생장 저해대 형성 여부를 조사하여 선별하였 다. Ochratoxin A (OTA)를 생성하는 Aspergillus alutaceus KCCM60421, A. awamori KCCM32316 및 A. fresenii KCCM60465은 한국미생물보존센터(Korean Culture Center of Microorganisms, KCCM)에서 분양받았고, fumonisin B1 (FB1)을 생산하는 Fusarium fugikuroi KACC46888과 48352, F. verticillioides KACC48356 및 F. proliferatum KACC48354는 국립농업과학원 한국농업미생물은행(Korean Agricultural Culture Collection, KACC)으로부터 분양 받 았다.
모든 대상 진균에 가장 높은 항진균 활성을 나타낸 세 균 균주 두 개를 최종 선발하여 API kit (50CH 및 20E, bioMérieux)를 이용한 생리생화학적 동정과 ㈜마크로젠 과 ㈜코스모진텍에 의뢰하여 염기서열을 분석하여 미국 National Center for Biotechnology Information의 등록균주 와 상동성을 비교하였고 균주의 염기서열은 GenBank에 등록하였다.
항진균 활성
Plate diffusion assay : 진균에 대한 생장 억제직경 조사를 위해 먼저 진균은 potato dextrose agar (PDA, Difco Lab)에
배양하였다. 선별 균주는 nutrient broth (NB, Difco Lab.)에 서 선배양 후 개체수를 보정하여(2 × 108 CFU/ml) NA에 100 μl를 도말하였다. 이들이 배지 위를 덮게 자라면 직경 10 mm cork borer로 뚫어내어 진균이 자라난 배지 disc는 새 PDA 중앙에 그리고 세균이 자라난 배지 disc는 주변부 에 올려 배양(30°C, 7일) 후 세균 주변에 생긴 진균 생장 저 해대의 직경을 측정하였다(Loqman et al., 2009).
진균 생장 및 포자형성 저해 : 선별 균주의 진균 생장과 포 자형성 억제능 조사는 먼저 선별 균주를 배양하고(NB, 48 h, 150 rpm, 37°C) 배양액을 원심분리(4°C, 2,800 × g, 40분) 하여 pellet을 회수 후 0.85% (w/v) NaCl로 2회 세척하였다.
세척한 세포는 분광광도계(Shimadzu UV-1700, Shimadzu Co.)로 OD600을 2.00으로 보정하여 세포 현탁액 1.5 ml와 액체상태의 PDA 13.5 ml를 섞어 petri dish에서 굳힌 뒤 중 앙에 paper disc (직경 5 mm)를 올리고 진균 포자 현탁액 (106 spores/ml)을 10 µl 분주하고 배양하였다(25°C, 10 days).
PDA에 세균 세포를 첨가하지 않은 대조구의 진균 생장 반 경과 처리구의 진균 생장 반경을 비교하여 진균 생장 저해 율을 나타내었다. 한편 진균이 자라난 PDA 표면에 0.05%
Tween 80 용액 15 ml을 반복적으로 살포하면 진균 포자가 서서히 Tween 80 용액에 현탁되므로 전량 현탁될 때까지 포자 회수를 반복한 후 대조구와 처리구의 포자수를 비교 하였다(Veras et al., 2016).
진균 포자 제거 및 발아 저해: 진균의 포자 제거와 발아 저 해에 대한 선별 균주 배양액의 효과는 Veras 등(2016)의 방 법을 수정해서 사용하였다. 1.5 ml microcentrifuge tube에 15% yeast extract sucrose (15% YES; yeast extract 20 g, sucrose 150 g, KH2PO4 1 g, MgSO4 0.5 g, DW 1 L, pH 6.2
± 0.2) 800 μl, 진균 포자 현탁액(107 spores/ml) 100 μl와 세 균 균주 현탁액(OD600 1.00) 100 μl를 첨가하고 배양 후 (30°C, 24~48시간) 광학현미경으로 포자수와 발아 정도를 관찰하였다. 대조구에는 세균 균주 현탁액 대신 0.85%
NaCl 100 μl를 첨가하였다. 대조구의 포자수와 비교하여 포자 제거율을 조사하였고 발아관의 길이가 포자의 지름 과 같거나 길 때 발아한 것으로 간주하였으며 각 실험구 당 무작위로 포자 100개를 관찰하여 저해율을 계산하였다.
액체배지에서 독소생성 진균 생장 및 진균독소 농도 변화 : 대상 진균 중 OTA를 생성하는 Aspergillus 3종에 대해 액체 배지에서 진균 건조중량과 독소농도를 조사하였다. 100 ml flask에 15% YES 배지 45 ml, 진균의 포자 현탁액(107
spores/ml) 500 µl와 OD600을 1.00으로 보정한 세균 균주 배 양액 5 ml를 첨가하여 배양하였다(30°C, 200 rpm, 7 days).
대조구는 세균 배양액 대신 15% YES 배지 5 ml를 첨가한 것을 이용하였다. 진균의 생장은 진균 균사체를 여과지 (Whatman No. 1)로 걸러내어 중량 변화가 없을 때까지 fume hood에서 실온 건조 후 무게를 측정하였고, 진균독소 는 균사체를 걸러낸 배양액에 100 ml의 chloroform을 넣어 2회 진탕하여(300 stroke/min, 20분) 추출하였다. 유기상은 flat bottom flask에 모아 회전 감압 증발기로 55°C에서 증 발시켰고 2 ml의 70% methanol로 재용해 후 시린지 필터 (Hyundai micro, 0.2 μm, hydrophobic)로 여과하여 형광검 출기(FLD; Agilent Technologies, 1260 Infinity, 333 nm excitation, 460 nm emission)가 부착된 high performance liquid chromatograph (Younglin, YL9100 HPLC System) 로 C18 column (150 × 4.6 mm; Waters Co.)을 이용하여 분 석하였다. 이동상은 water/acetonitrile/acetic acid (99:99:2, v/v), 유속은 1 ml/min, 주입량은 20 μl, column oven 온도 는 30°C로 설정하여 15분 동안 분석하였다(Xiong et al., 2017).
항진균 물질
Siderophore : Siderophore 정량 분석은 먼저 세균 균주 선 배양액을 2 × 108 CFU/ml로 보정하여 90 ml NB에 10% 접 종하여 배양액을 준비하였다(150 rpm, 37°C). 24시간마다 세균 균주 배양액(15 ml)을 원심분리(4°C, 2,800 × g, 40분) 하여 상등액 10 ml를 회수하고 pH 2.0으로 보정 후 동량의 ethyl acetate를 첨가하여 진탕하였다. 유기상 5 ml를 회수 하여 Hathway 용액을 동량 첨가 후 반응시켰다(30분, 암조 건). 그 후 분광광도계(Shimadzu UV-1700, Shimadzu Co.) 로 OD700을 측정하였고 siderophore 농도는 dihydroxy benzoic acid로 작성한 표준곡선으로 정량하였다(Nagarajkumar et al., 2004).
β-1,3-Glucanase : 먼저 세균 균주 선배양액을 2 × 108 CFU/ml로 보정하여 45 ml NB에 10% 접종하여 배양액을 준비하였다(150 rpm, 37°C). 세균 균주 배양액을 24시간 주 기로 2 ml씩 회수하여 원심분리(4°C, 5,000 × g, 20분) 후 상 등액 900 µl와 10% laminarin 100 µl를 반응시켰다(40°C, 10분). 반응액 100 µl에 dinitrosalysilic acid (DNS) 용액 200 µl을 첨가하여 3분간 100°C에서 반응 후 5분간 냉각시 켜 반응을 종료시켰다. 그 후 OD550을 측정하고, glucose로
작성한 표준곡선으로 β-1,3-glucanase를 정량하였다. 효소 의 활성은 40°C에서 1분간 1 mmol의 glucose를 생산하는 효소의 양을 1 unit으로 정의하였다(Marcello et al., 2010).
Chitinase : Chitinase 활성을 조사하기 위해 세균 균주를 basal salt broth [K2HPO4 0.7 g, KH2PO4 0.3 g, MgSO4 0.5 g, FeSO4 0.01 g, ZnSO4 0.001 g, MnSO4 0.001 g, (NH4)2SO4
0.25 g, yeast extract 1 g, 증류수 1 L, 0.1% (w/v) colloidal chitin]에서 선배양하고 2 × 108 CFU/ml로 보정하여 45 ml basal salt broth에 접종 후 배양하였다(150 rpm, 37°C). 24 시간마다 3 ml를 채취하여 원심분리(4°C, 2,800 × g, 40분) 후 상등액을 회수하였다. 상등액 500 μl와 1% chitin이 첨가 된 50 mM sodium phosphate buffer (pH 7) 500 μl를 반응 (37°C, 1시간) 후 DNS 용액과 혼합시켰다. 시료의 흡광도 는 575 nm에서 측정하고 N-acetyl-D-glucosamine (NAG) 로 표준곡선을 작성하여 정량하였다. 효소활성 1 U는 60분 동안 1 ml 당 1 μmol의 NAG를 생성하는 효소의 양으로 정 의하였다(Nagpure and Gupta, 2013).
Protease : Protease 활성 조사는 세균 균주를 NB 배지에 선 배양하고 2 × 108 CFU/ml로 보정하여 45 ml NB 배지에 10% 첨가한 뒤 배양(150 rpm, 37°C) 24시간 간격으로 1 m 씩 회수하여 원심분리(4°C, 5,000 × g, 20분) 후 상등액 500 μl에 1% casein이 함유된 100 mM Tris-HCl (pH 8.0) 완충 액을 동량 첨가하여 반응시켰다(60°C, 15분). 반응액에 20%
trichloroacetic acid를 500 μl 혼합하여 반응을 종료시켰고 원심분리(4°C, 12,000 × g, 20분) 후 상등액의 흡광도를 280 nm에서 측정하고 표준물질로 tyrosine을 이용하여 효소 생 성을 정량하였다(Ghorbel et al., 2003).
통계분석
모든 실험은 3회 반복하였으며 분석에 이용한 프로그램 은 SPSS v. 24.0 (IBM SASS Statistics 24)였고 대조구와 처 리구 사이 유의한 차이는 Independent two-sample t-test를 이용하여 분석하였다. 모든 실험에서 p < 0.05일 때 통계적 으로 유의한 차이가 있다고 보았다.
결과 및 토의
세균 균주의 선별 및 동정
다양한 출처의 토양시료에서 분리한 1,000여 개의 세균
Table 1. Growth inhibition of AF11 and YR47 strains against toxigenic fungi by plate diffusion assay for 7 days at 30°C
Target organism Inhibition zone (mm)
AF11 YR47
A. alutaceus KCCM60421 27.0 ± 2.3 28.0 ± 0.0 A. awamori KCCM32316 20.0 ± 4.2 13.3 ± 2.6 A. fresenii KCCM60465 40.0 ± 0.6 25.7 ± 0.7 F. fugkuroi KACC46888 32.7 ± 1.5 19.0 ± 1.0 F. fugkuroi KACC48352 29.0 ± 2.0 19.3 ± 0.7 F. verticillioides KACC48356 33.0 ± 0.6 19.3 ± 1.2 F. proliferatum KACC48354 32.3 ± 0.7 19.0 ± 0.6
Table 2. Effect of strains AF11 and YR47 on colony radius (CR) and sporulation inhibition (SI) of toxigenic fungi during after incubation (10 days, 25°C)
Target organisms Control AF11 YR47
CR (mm) CR (mm) SI (%) CR (mm) SI (%)
A. alutaceus KCCM60421 40.7 ± 1.5 0.0 ± 0.0*** 99.8 ± 0.2* 8.7 ± 0.6*** 95.4 ± 1.2*
A. awamori KCCM32316 27.0 ± 0.0 11.7 ± 0.6*** 100.0 ± 0.0*** 0.0 ± 0.0*** 100.0 ± 0.0***
A. fresenii KCCM60465 39.0 ± 1.0 6.3 ± 0.6*** 100.0 ± 0.0* 4.3 ± 0.6*** 100.0 ± 0.0*
F. fugikuroi KACC46888 43.3 ± 2.1 13.7 ± 0.6*** 97.6 ± 0.5* 20.7 ± 0.6*** 98.9 ± 0.3*
F. fugikuroi KACC48352 40.7 ± 1.5 18.9 ± 0.6*** 78.9 ± 1.3* 14.3 ± 0.6*** 87.6 ± 1.0*
F. verticillioides KACC48356 42.0 ± 1.0 9.0 ± 1.7*** 95.3 ± 1.7** 8.3 ± 1.2*** 96.0 ± 1.1**
F. proliferatum KACC48354 41.7 ± 0.6 19.7 ± 0.6*** 77.7 ± 1.3*** 20.7 ±1.2*** 75.4 ± 2.8***
* p<0.05; ** p<0.01; *** p<0.001.
균주 중 AF11과 YR47 균주가 독소생성 진균 7종에 대한 항진균 활성이 높게 나타났다. 두 선별 균주에 대한 API kit 를 이용한 생리생화학적 검사에서 전형적인 Bacillus 종의 특성을 나타내었으며(결과 미제시), 16S rDNA 염기서열 분석 결과 AF11은 Bacillus subtilis DSM 10과 100%의 상 동성을 나타냈고 aflatoxin B1을 생산하는 Aspergillus flavus 에 대한 항진균 활성이 보고되었다(Lee, 2018). YR47의 염 기서열은 미국 NCBI database 등록 균주와 비교 시 Bacillus subtilis Y22의 염기서열과 100%의 상동성을 나타내었다.
YR47 균주의 염기서열은 MN559293으로 GenBank에 등 록하였고 생물자원센터(KCTC)에 KCTC18663P로 기탁 하였다.
항진균 활성
Plate diffusion assay : 독소생성 진균에 대한 plate diffusion assay에서 AF11과 YR47은 대상 진균 모두에 13.3 mm 이 상의 억제 직경을 나타냈다(Table 1). 특히 AF11은 A. fresenii KCCM60465에 40.0 mm의 억제환을 나타냈고 YR47은
A. alutaceus KCCM60421를 28.0 mm 억제하였는데 이는 Lactobacillus sp. 균주가 OTA 생성 A. ochraceus의 생장을 억 제하지 못한 결과(Gourama, 1997)와 대조적이다. Juodeikiene 등(2018)의 보고에서 Lactobacillus sakei KTU05-6는 Fusarium avenaceum M8에 대한 억제직경이 11.8 mm였는데 본 균 주들은 19.0 mm 이상의 억제직경을 나타냈으며, L. brevis NS는 Fusarium sp.를 억제하지 못했지만 AF11과 YR47 균 주는 Fusarium spp.에 대한 높은 항진균 활성도 나타냈다.
진균 생장 및 포자형성 저해 : AF11과 YR47 균주는 모든 대상 진균에 대해 생장 직경과 포자형성을 유의하게 저해 하였다. AF11은 A. alutaceus KCCM60421을, YR47은 A.
awamori KCCM32316의 생장을 완전히 제어하였고, 두 균 주는 A. awamori KCCM32316와 A. fresenii KCCM60465 의 포자형성에 대해 100% 저해율을 나타냈다(Table 2). Shi 등(2014)이 보고한 Bacillus subtilis CW14는 OTA 생성 A.
ochraceus 3.4412와 A. carbonarius의 생장을 28.8~44.4%
저해하였는데, 같은 종인 AF11과 YR47은 이보다 더 높은 진균 생장 저해를 나타냈다. Lactobacillus plantarum E98 균주는 Fusarium avenaceum D141의 생장을 32% 제어한 반면(Laitila et al., 2002), 본 균주들은 이보다 더 우수하게 Fusarium spp.의 생장을 억제하였다.
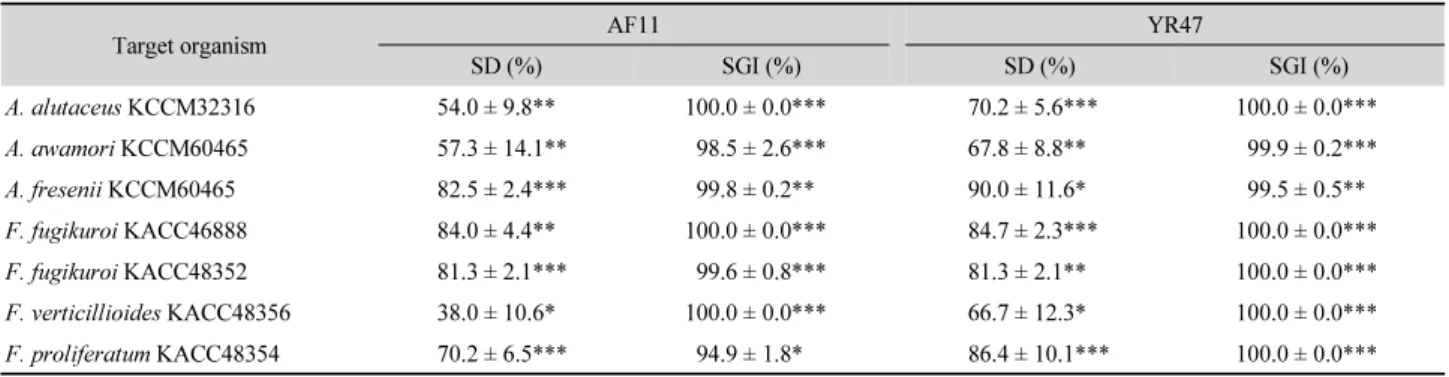
진균 포자 제거 및 발아 저해 : 대조구와 균주 처리구의 진 균 포자수 및 포자 발아 정도를 조사하였을 때 대조구에서 는 대부분의 포자가 상당히 발아하였지만 AF11과 YR47 처리구에서는 상당수의 포자가 제거되어 대조구에 비해 유의하게 잔류 포자수가 적었으며, 남아있는 포자 중 발아 한 것이 거의 없었다(Fig. 1). AF11은 대상 진균의 포자를 38.0% 이상 제거하였으며 포자 발아 또한 94.9% 이상 억제 하였다. YR47은 66.7% 이상으로 포자를 제거하였으며 포
(A)
(B)
(C)
Fig. 1. Effect of strains AF11 and YR47 on spore degradation and germination of A. fresenii KCCM60465 after 24 h incubation at 30 in 15% YES medium [control (A), AF11 (B), and YR47 (C)].
Table 3. Effects of strains AF11 and YR47 on spore degradation (SD) and spore germination (SGI) of toxigenic fungi after 24 h incubation at 30°C in 15%
YES broth
Target organism AF11 YR47
SD (%) SGI (%) SD (%) SGI (%)
A. alutaceus KCCM32316 54.0 ± 9.8** 100.0 ± 0.0*** 70.2 ± 5.6*** 100.0 ± 0.0***
A. awamori KCCM60465 57.3 ± 14.1** 98.5 ± 2.6*** 67.8 ± 8.8** 99.9 ± 0.2***
A. fresenii KCCM60465 82.5 ± 2.4*** 99.8 ± 0.2** 90.0 ± 11.6* 99.5 ± 0.5**
F. fugikuroi KACC46888 84.0 ± 4.4** 100.0 ± 0.0*** 84.7 ± 2.3*** 100.0 ± 0.0***
F. fugikuroi KACC48352 81.3 ± 2.1*** 99.6 ± 0.8*** 81.3 ± 2.1** 100.0 ± 0.0***
F. verticillioides KACC48356 38.0 ± 10.6* 100.0 ± 0.0*** 66.7 ± 12.3* 100.0 ± 0.0***
F. proliferatum KACC48354 70.2 ± 6.5*** 94.9 ± 1.8* 86.4 ± 10.1*** 100.0 ± 0.0***
* p<0.05; ** p<0.01; *** p<0.001
자 발아도 최대 100% 제어하였다(Table 3). 이 포자 발아 저 해는 Gourama (1997)가 보고한 Lactobacillus sp. A2와 A3 의 독소생성 진균 5종의 포자발아 저해능(84~85%)보다 높 았다. 진균 포자 제거는 세균 균주가 chitinase 혹은 β-1,3- glucanase 등과 같은 진균 세포벽 분해효소를 생산하여 제 거하는 것으로 판단된다. 많은 Bacillus 종이 진균 질병에 대한 생물학적 제어제로 잘 알려져 있으며(Ongena and Jacques, 2008), 이들이 생산하는 일부 물질이 진균의 포자 발아를 억제하는 것으로 보고되었다(Leelasuphakul et al., 2008).
액체배지에서 독소생성 진균 생장 및 독소농도 변화 : YES 배지에서 OTA 생성 진균 3종과 세균 균주를 동시 접종 후 7일 배양 시, A. alutaceus KCCM60421, A. awamori KCCM 32316과 A. fresenii KCCM60465의 대조구 건조중량은 각각 1.2, 2.3 및 2.1 g이었고 균주 처리구는 건조중량을 86% 이상 감축시켰다(Fig. 2). 그 중 A. alutaceus KCCM 60421 진균의 YR47 처리구와 A. fresenii KCCM60465 진균의
AF11 및 YR47 처리구는 진균의 생장을 완전히 저해하였 다. 진균이 생산한 OTA 농도는 2.5 μg/L에서 1.9 mg/L으로 다양했지만 균주 처리구는 독소농도를 모두 54% 이상 감축 시켰고 그 중 A. alutaceus KCCM60421가 배양 7일차에 생 산한 1.9 mg/L의 OTA를 YR47이 99.9% 감소시켜 1 μg/L의 OTA가 검출되었다(Fig. 3). 이는 AF11과 YR47 균주가 진균 의 생장을 효율적으로 억제하기 때문에 독소생산 자체를 저해하는 것으로 판단되며 이 균주들이 진균독소 제거능 이 있기 때문에(Lee, 2018) 잔류 독소를 더욱 저감시킨 것 으로 추정된다.
항진균 물질
균주가 생산하는 물질 중 항진균 활성이 있다고 알려진 siderohore, chitinase, β-1,3-glucanase 및 protease의 생성 을 조사하였다. Siderophore는 환경에 존재하는 미량원소 인 철 이온을 독점하여 가용성을 감소시켜 항균 활성을 나 타낸다고 알려져 있는데(Husen, 2003), YR47은 배양 96시
(A) (B) (C)
Fig. 2. Growth inhibition of ochratoxigenic A. alutaceus KCCM60421 (A), A. awamori KCCM32316 (B), and A. fresenii KCCM60465 (C) by simultaneous inoculation of strains AF11 and YR47 in 15% YES medium (7 days, 30°C, 200 rpm). ** p<0.01; *** p<0.001.
(A)
(B) (C)
Fig. 3. Inhibition of OTA production of ochratoxigenic A. alutaceus KCCM60421 (A), A. awamori KCCM32316 (B), and A. fresenii KCCM60465 (C) by simultaneous inoculation of strains AF11 and YR47 in 15% YES medium (7 days, 30°C, 200 rpm). ** p<0.01; *** p<0.001.
(A) (B)
(C) (D)
Fig. 4. Production of siderophore (A), β-1,3-glucanase (B), chitinase (C), and protease (D) by Bacillus subtilis AF11 (○) and YR47 (◇).
간에 siderophore를 50 μM로 가장 많이 생산하였다(Fig.
4A). β-1,3-Glucanase와 chitinase는 진균 세포벽 성분 중 glucan과 chitin을 분해하는 효소로서(Fridlender et al., 1993)
YR47은 β-1,3-glucanase와 chitinase를 각각 배양 72시간과 24시간에 65.8 U/ml와 1.5 U/ml의 최대 활성을 나타내어 (Fig. 4B and C) Bacillus subtilis AJF-1A5의 β-1,3-glucanase
활성 51 U/ml (Tang et al., 2004) 그리고 Rahmawati 등 (2016)이 분리한 31개 균주의 chitinase 최대 활성 0.46 U/ml보다 높은 결과를 나타내었다. Protease 또한 chitinase 및 glucanase와 함께 진균의 세포벽을 분해할 수 있는 효소 로서 진균 세포의 용해나 사멸을 유도한다(Tseng et al., 2008). AF11 균주는 배양 72시간에 265 mg/L, YR47 균주 는 48시간 배양 후 238 mg/L의 protease를 생산하였다(Fig.
4D). AF11 균주는 β-1,3-glucanase는 생성하지 못하였으 나 1.2 U/ml의 chitinase 활성을 나타내었으며 특히 배양 48 시간에 122 μM의 매우 높은 siderophore 생성능을 가진 것 으로 보고되었다(Lee, 2018). AF11과 YR47 균주는 이와 같은 항진균 물질을 분비하여 독소생성 진균의 생장, 포자 생성 및 포자 발아를 저해하고 이차 대사산물인 독소의 생 성을 제어한다고 판단된다.
적 요
독소생성 진균은 사료 및 곡물 등을 오염시키며 독소를 생산하는데 이는 저농도로도 인간과 동물에게 독성과 발 암성을 나타낸다. 본 연구에서는 토양에서 분리한 Bacillus subtilis AF11과 YR47 균주의 독소생성 진균에 대한 항진 균 활성과 이들의 항진균 물질을 조사하였다. 이 균주들 은 평판확산시험에서 균사 생장을 효과적으로 억제하여 3개의 오크라톡신 생성진균과 4개의 퓨모니신 생성진균 에 대해 13.3 mm 이상의 억제 직경을 나타내었다. AF11과 YR47은 또한 대상 진균의 포자형성을 대조군에 비해 각각 77.7~100.0 및 75.4~100.0% 저해하였다. 이들은 진균 포자 (107 spores/ml)를 38~90% 제거하였으며, 잔류 포자의 발아 를 94.9% 이상 억제하였다. YES 액체배지에서 오크라톡신 A (OTA) 생성 Aspergillus와 AF11 또는 YR47을 동시 접종 시 OTA 생성이 효과적으로 낮아졌다. 이는 이 균주들이 생 성하는 siderophore, β-1,3-glucanase, chitinase 및 protease 를 포함한 항진균 물질 때문인 것으로 판단된다. 이 결과는 Bacillus subtilis AF11과 YR47 균주가 독소생성 진균의 생장 및 진균독소 생성의 제어에 이용될 수 있는 것을 제시한다.
References