성장에 따른 백서 심근세포의 허혈 손상에 대한 Protein Kinase C의 보호 효과 차이
8
0
0
전체 글
(2) nary intervention, PCI)2)을 이용하여 막힌 혈관을 개. 아형은 허혈에 노출된 신생쥐 심근세포에서 세포질로부. 통하여 심근 관류를 재개함으로서 심근 조직의 손상을. 터 세포막으로 이동함이 관찰되었으며9) PKCδ 아형은. 최소화할 수 있다. 그러나 현실적으로는 환자의 시간 지. 재관류에 따른 손상으로부터 심근 보호 작용을 중재함. 연이나 동반된 질환, 또는 의료시설의 미비 등으로 혈류. 이 보고되었다.10) 이 중 허혈에 대한 심근 보호 작용과. 재개통이 불가능하거나 효과를 기대할 수 없는 경우가 흔. 의 연관성이 두드러지게 보고된 것은 PKCε 아형이다.. 3). 히 존재한다. 그러므로 허혈 자체를 제거하지 못하더라. PKCε 아형은 저산소증에서 세포막으로 이동할 뿐만 아. 도 허혈 자체에 대한 심근 조직을 보호할 필요성이 부각. 니라 선택적 억제제인 εV1-2 투여시 허혈에 대한 심근. 되었고 이에 따라 심근세포가 가지고 있는 자체 방어 기. 세포 보호 작용이 소실됨이 관찰되었다.11) 또한 guinea. 전을 밝혀내어 이를 이용하고자 하는 시도가 나타났다.. pig 모델에서 규칙적인 에탄올의 섭취에 의한 심장 보호. 허혈에 대한 세포의 고유 방어 기전은 1986년 Rei-. 작용이 심장에서의 PKCε 아형의 지속적인 활성화와. 4). mer 등 의 개 실험에서 허혈에 일단 노출된 심장은 이. 관련되어 있음이 보고되기도 하였다.12). 후의 허혈 상태에서 오히려 조직 손상이 줄어드는 것을. 한편 쥐 실험에서 심장 조직의 PKC 아형은 태생기,. 5). 관찰함으로써 그 존재가 처음 발견되었고 Murry 등. 신생기를 거쳐서 성체에 이르는 과정에서 변화가 일어나. 은 이를 허혈성 전처치(ischemic preconditioning)이. 는 것으로 알려져 있다. Clerk 등13)의 연구에 의하면 신. 라고 명명하였다. 허혈성 전처치의 기전을 규명코자 하. 생쥐 심근에서는 PKC α, βI, δ, ε, ζ 등의 아형이. 는 다수의 연구 결과 이를 중재하는 물질들이 밝혀졌는. 주를 이루다가 이후 이들 전체는 신속히 down-regul-. 데 그 중 protein kinase C(PKC)는 가장 중심적인 역. ation 되나 성체가 되면서 PKCε을 비롯한 일부 아형. 할을 하는 것으로 알려져 있다. 허혈성 전처치를 유도하. 은 심근세포당 함량이 신생쥐에 비해 약 10배 가량 증가. 는 물질들, 예를 들어 adenosine과 bradykinin 등은. 하는 것으로 관찰되었다. 국내에서도 박 등14)이 in situ. receptor와 연계된 G protein을 자극하여 PKC를 세포. hybridization을 이용하여 쥐 심근에서 발생, 발육에 따. 막으로 유인하여 활성화하고 활성화된 PKC는 ATP-. 른 PKC의 아형 변화를 보고하였다. 심근 보호 작용에. dependent potassium channel(KATP channel) 등 세. 서 중심 역할을 하는 PKC 아형의 함량이 성장에 따라. 포내 다른 물질에 작용하여 보호작용을 나타내는 것으. 크게 변하면 신생쥐와 성체쥐에서 허혈시 PKC 활성화에. 6). PKC는 일정하게 보존된(conserved). 따른 심근세포의 생존, 사멸 양식은 다르게 나타날 수 있. kinase domain (C3/C4) 외에 calcium binding dom-. 다. 그러나 이러한 차이가 허혈시 실제로 나타나는지를. ain(C2)과 cysteinerich, zinc finger-like motif(C1). 관찰한 연구는 아직 보고된 바 없다. 그러므로 본 연구. 의 regulatory domain으로 구성되어 있으며 domain 구. 의 목표는 신생쥐와 성체쥐에서 분리된 심근세포를 대상. 성 여건에 따라 여러 개의 아형(isoform)이 존재하는 것. 으로 하여 PKC 활성화나 억제에 따른 심근 보호 효과. 로 밝혀졌다.. 7). 차이를 규명하고자 하였다.. 으로 나타났다.. 즉 PKC는 각 domain을 모두 갖춘 conventional. 재료 및 방법. PKC isoform(cPKC;α, βI, βII, γ)와 calcium binding domain이 결핍된 novel PKC isoform(nPKC; δ, ε, η/L, θ), 그리고 단수의 cysteinerich domain. 재 료. 만이 존재하는 atypical PKC isoform(aPKC;ζ, λ,. 성체쥐는 중량이 200~250 g 정도 되는 수컷 흰쥐. ι, μ)으로 분류되며 각 아형에서 효소의 활성화에 필. (Sprague-Dawley rat)를 사용하였으며 신생쥐는 같. 요한 calcium이나 diacylglycerol(DAG)에 대한 의존도. 은 종으로서 생후 1일에서 2일된 쥐를 택하였다.. 8). 는 서로 다르게 나타난다.. 이와 같이 PKC는 여러 아. 형으로 존재하고 또한 각각의 PKC 아형은 서로 다른 작. 방 법. 용을 가지고 있다고 알려져 있으므로 허혈에 대한 심근 보호 작용도 PKC 전체가 아닌 어떤 PKC 아형에 의해 특이적으로 중개될 것으로 생각된다. PKCα와 PKCε 690. 15). 성체쥐 심근세포의 분리 배양. 쥐를 dry ice가 담긴 통에 넣어 기절시킨 후 신속히 Korean Circulation J 2002; 32(8):689-696.
(3) 심장을 절제하여 Langendorff 기구에 매달고 대동맥. EDTA(Gibco)액에 넣고 4℃ shaker에서 밤새 소화시. 에 pH 7.4와 5% CO2/95% O2에서 평형시킨 Krebs-. 켰다. 다음날 trypsin 효소액은 버리고 10% fetal calf. Henseleit bicarbonate(KHB) buffer(118 mM NaCl,. serum(FCS;Gibco)가 함유된 DMEM액을 넣어 소화. 4.7 mM KCl, 1.75 mM CaCl2, 4.0 mM MgSO4, 0.5. 과정을 중화시킨 후 DMEM 상층액을 제거하였다. 여기. mM EDTA, 25 mM NaHCO3, 11.0 mM glucose, Si-. 에 collagenase 효소액(34 mg collagenase II와 DM-. gma, USA)를 관류시켜 혈액을 제거하였다. 5분 정도. EM 혼합용액)을 더하고 37℃ shaking bath에 3분 정. 지난 뒤 관류액을 Ca++ free KHB 완충액으로 바꾸고. 도 두었다가 상층부에 있는 소화액만 conical tube에. 심장 박동이 멈출 때까지 관류시켰다. 이어 0.05% co-. 모았다. 이 같은 과정을 3차례 정도 반복하여 모은 상. llagenase II(Biochemical, Worthington, UK)와 0.03%. 층액을 750 rpm에서 5분간 원심한 후 pellet은 10%. ++. free KHB 혼. FCS/DMEM액으로 재부유시켰다. 다음 이를 culture. 합액을 심근이 창백하고 긴장도가 떨어질 때까지 관류. flask에 옮기고 37도에서 75분간 배양하고 이후 부유. 시켜 조직을 소화(digestion)시켰다. 다음에 심장을 분. 액을 다시 새 culture flask에 옮기고 다시 75분간 배. 리하여 심근 조직만을 떼내어 trypsin(type IX)/DNase. 양하여 상층액인 심근세포와 접착된 비심근세포를 분리. ++. (Biomedical, Worthington, UK)이 추가 배합된 Ca. 하였다. 분리된 심근세포는 0.1% penicillin/streptomy-. free KHB 혼합 효소액에서 가위로 잘게 자르고 이를. cin이 함유된 10% FCS/DMEM 배지에서 5% CO2/95%. 모아 20분간 37℃ shaking water bath에서 소화시켰. O2의 조건하에 배양하였다.. hyaluonidase(Sigma)가 함유된 Ca. 다. 소화액을 모아서 세척 완충액(wash buffer) 50 ml Dulbecco’s modified Eagle’s media(DMEM;Gi++. bco, USA)와 50 ml Ca. 심근세포의 허혈 배양. free KHB 혼합액을 혼합한. 저산소 공급(hypoxia)은 95% N2 /5% CO2 gas가. 후 500×g에서 3분간 원심하고 상층액은 버리고 남은. 담긴 탱크로부터 관을 통하여 incubator내에서 심근세. 부분을 세척 완충액으로 재부유시켰다. 이를 bovine se-. 포를 배양중인 용기에 연결하여 저산소 기체를 공급하. rum albumin(Sigma)을 함유한 DMEM 용액 위에 붓. 였다. 용기 입구는 고무마개로 막고 N2 gas가 출입할. 고 심근세포들이 가라앉을 때까지 약 13분 정도 기다. 수 있는 입구만을 남겨 놓았다. 저산소 공급과 함께 배양. 렸다가 상층액을 버리고 DMEM 용액으로 부유시켰다.. 배지 대신 Ca++ free Henseleit buffer(6.9 g/L NaCl,. 평판배양은 미리 laminin(Collaborative Biomedical. 350 mg/L KCl, 180 mg/L CaCl2, 144 mg/L MgSO4,. Product, USA)을 넣어서 배양해 둔 평판에 DMEM액. 162 mg/L KH2PO4, 2.1 g/L NaHCO3, 2.16 g/L dex-. 을 10ml 정도 붓고 세포 부유액을 분배하였다. 분리된. trose;Sigma)를 배양액으로 교체하였으며 허혈군은. 성체 심근세포는 Volz 등16)에 의해 고안된 배양 배지. 이중 dextrose 대신에 deoxydextrose를 동량 사용하. (2 mg/mL albumin, 2 mmol/L L-carnitine, 5 mmol/. 여 영양원을 박탈하였다. 이 배양액을 사용하기 전에 95%. L creatine, 5 mmol/L taurine, 1.3 mmol/L L-gluta-. N2/5% CO2 gas를 20분간 공급하여 배지내의 남아 있. mine, 0.1 μmol/L insulin, 0.1 mmol/L triiodothyron-. 는 산소를 미리 제거하였다.. ine, 2.5 mmol/L pyruvate, 0.1% penicillin/streptomycin;Sigma)와 5% CO2 /95% O2의 조건에서 배양 되었다.. 18). 심근세포의 생존/사멸 평가:Trypan blue staining. Trypan blue(Sigma)와 배지를 각각 동량으로 혼합 한 후 실온에서 약 2분간 두었다가 이를 광학 현미경으 17). 신생쥐 심근세포의 분리 배양. 로 관찰하여 trypan blue에 짙게 염색되는 세포는 사망. 생후 1일 정도 되는 신생쥐의 흉곽을 절개한 후 신속. 한 것으로 간주하고 trypan을 배척하여 염색되지 않는. 히 절제한 심장을 Hank’ s buffered salt solution(HB-. 세포는 살아있는 세포로 간주하여 단위 면적당의 생존한. SS;Gibco)에 담구고 가위로 잘게 자른 후 이를 다시. 세포의 수를 관측하였다. 성체쥐의 경우 수축하는 세포. HBSS로 세척하였다. 그 후 조직을 건져내어 trypsin/. (beating cell) 외에 정지한 세포(quiescent cell)라도. 691.
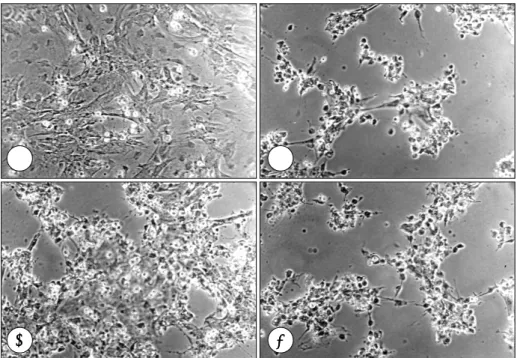
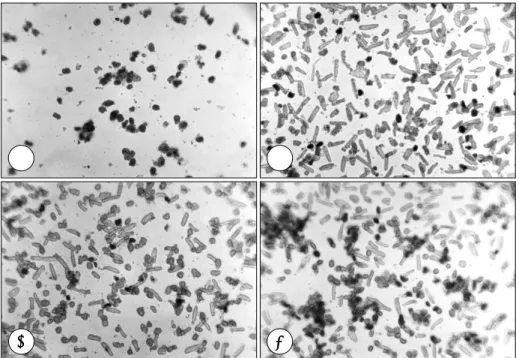
(4) 원래의 막대 형태(rod shape)를 유지하고 있으면 생존 세포로 간주하였다.. 통계학적 분석 모든 실험은 3회 이상 반복 시행하였다. 대조군과 허 혈군 사이의 생존 비교는 unpaired t-test로 하였으며 p 값이 0.05 미만을 유의한 것으로 하였다.. PKC 활성화 및 억제 19). PKC를 활성화하는 물질로서 phorbol ester 의 일종. 결. 인 12-o-tetradecanoylphorbol 13-acetate(TPA;. 과. Sigma)를 100 nM의 농도로서 배지에 첨가하였으며 PKC 의 억제제로는 staurosporin(Sigma)을 100 nM 농도 로 첨가하였다.. 허혈 상태에서의 성체쥐와 신생쥐 심근세포의 생존 차이 허혈 상태에서 trypan blue 염색으로 산출한 단위 면 적 당의 평균 생존율은 신생쥐에서는 6시간 경과 시 68.9. Fig. 1. Survival of myocyte under control (white bar) and ischemic condition (black bar) as determined by trypan blue exclusion. A:neonatal rat ventricular myocyte, B:adult rat ventricular myocyte. *:p<0.5 between control and ischemia group.. A. B. C. D. Fig. 2. Trypan blue staining of neonatal rat ventricular myocyte cyultured for 24 hours under various conditions. A:control, B:ischemia, C:ischemia+ staurosporin 100 mM, D:ischemia+ 12-o-tetradecanoylphorbol 13-acetate (TPA) 100 mM.. 692. Korean Circulation J 2002; 32(8):689-696.
(5) ±6.4%, 12시간 60.3±7.3%, 24시간에서 34.3±7.5%,. 즉 신생쥐의 경우 허혈 24시간에서 성체쥐의 경우는 허. 36시간 경과시 8.2±6.6%로 나타났으며(시간에 따른. 혈 6시간 경과 시점에 실험을 하였다. Fig. 2는 신생쥐. 유의성 p=0.004) 이에 비해 성체쥐에서는 허혈 3시간. 에서의 trypan blue 염색으로 허혈 24시간에 다수의 세. 경과 시에 63.1±8.3%, 6시간 경과 시 42.8±6.1%, 12. 포 사망을 보이고 있으며 생존 세포는 staurosporin 투. 시간 경과 시 10.1±5.8%로서(p=0.031) 분리된 성체. 여에 의해서 약간의 감소를 보이나 100 nM TPA를 투. 심장의 심근세포에서 허혈 손상에 의한 세포 사멸이 이. 여한 군의 세포 생존 비율은 큰 변화를 보이지 않았다. 반. 른 시간 내에 더 많이 발생하였다(Fig. 1).. 면 Fig. 3에서 성체쥐의 심근세포는 허혈 6시간에서 생. 신생쥐와 성체쥐에서 PKC 활성화에 따른 심근 보호 효과. urosporin에 의해서 생존 세포를 거의 찾아볼 수 없을. 의 차이. 정도로 세포 사망이 늘어났다. 그러나 PKC 활성제제인. 존 세포의 감소를 보이고 있으며 PKC 억제제인 sta-. 각각의 군에서 충분하게 세포의 사망이 나타나는 시간. 100 nM의 TPA를 투여한 군에서는 거의 비허혈 대조. A. B. C. D. Fig. 3. Trypan blue staining of adult rat ventricular myocyte cyultured for 6 hours under various conditions. A: control, B:ischemia, C:ischemia+ staurosporin 100 mM, D:ischemia+ 12-o-tetradecanoylphorbol 13-acetate (TPA) 100 mM.. Fig. 4. Survival of myocyte under control (white bar) and ischemic condition (black bar) under various conditions as determined by trypan blue exclusion. A:neonatal rat ventricular myocyte (24 hr culture), B:adult rat ventricular myocyte (6 hr culture). *:p<0.5 between control and ischemia group. Concentration of staurosporin and 12-o-tetradecanoylphorbol 13-acetate (TPA) was 100 mM respectively.. 693.
(6) 군과 같은 정도로 많은 세포들이 생존해 있었으며 그 형. 도를 사전에 아는 것이 필요하였다. 그 결과 같은 허혈. 태도 정상적인 막대 형태(rod shape)를 하고 있음을 보. 조건에서도 허혈에 대한 세포의 손상은 성체쥐에서 보다. 여주었다.. 크고 먼저 발생함을 관찰하였다. 즉 성체쥐에서 분리된. Fig. 4는 각각 허혈 상태의 신생쥐와 성체쥐에서 st-. 심근세포는 신생쥐의 것보다 허혈에 매우 취약하였다. 실. aurosporin과 TPA가 심근 보호 효과에 미치는 효과를. 험에서 신생쥐의 분리 세포는 과반수의 세포 사망이 일. 정량적으로 도표화한 것이다. 이 실험에서 신생쥐는 허. 어나는 것이 허혈 후 24시간이 걸렸으나 성체쥐의 경우. 혈 24시간에서 평균 31.8±5.8%의 생존율을 보였으며. 허혈 6시간이 지나면 생존 세포가 절반 이하로 감소하였. staurosporin 투여군에서 16.9±6.4%로 감소하였으며. 다. 이는 기본적으로 신생쥐의 세포가 젊다는 사실 외에. TPA 투여군에서의 생존율은 28.5±7.3%로서 PKC 활. 도 세포들이 교회(confluence)에 이를 때까지 혈청이 함. 성화에 따른 보호 효과는 보이지 않았다. 반면 성체쥐 심. 유된 배지에서 키운다는 점이 전반적인 세포의 생존능력. 근세포의 생존율은 허혈 6시간에 38.5±8.3%이나 PKC. 을 높이기 때문으로 생각된다. 그리고 신생쥐의 세포는. 억제제인 staurosporin에 의해서 생존율이 0%로 떨어. 교회에서 서로 접촉을 하게 됨으로써 세포간 교류나 상. 졌으며 이 같은 감소는 허혈 조건이 없는 대조군에서도. 호작용 등에 의해 생존을 향상시킬 수 있다. 이에 반해. 발견되었다. 그러나 TPA로 인한 PKC의 활성화로 심근. 성체쥐의 심근세포는 분리되면 따로 따로 고립된 세포. 세포는 허혈하에서도 62.1±7.3%의 생존율을 유지함을. 로서 존재하므로 상호 접촉에 의한 자극을 상실하여 고. 보여주었고 이는 허혈 조건을 주지 않은 대조군(65.4±. 사에 이르는 소위 anoikis 현상20)에 의해 세포가 사멸할. 6.2%)과 대등한 수준이었다.. 수 있다. 또한 성체쥐의 심근세포는 분리 후 배양 배지 로 기본적인 대사 기질로 구성된 용액을 공급받는데 이. 고. 찰. 는 혈청과 달리 성장인자 등이 전혀 없으므로 생존을 제고하는 요인이 없는 것도 한 원인일 수 있다.21) 비록. 허혈에 대한 세포 고유의 방어 기전인 허혈성 전처치. 허혈 조건을 공급할 때 양쪽 세포가 동일한 허혈 배지. 에 대하여 다수의 연구 결과로서 PKC의 역할이 밝혀졌. 로 배양되었지만 이러한 요소가 생존에 영향을 미쳤을. 음에도 불구하고 그 작용 기전이 아직 완전하게 규명된. 가능성은 완전히 배제하기 어렵다. 이러한 결과들은 세. 것은 아니다. PKC의 경우 많은 아형이 존재하므로 어떤. 포의 분리와 배양과정을 고려하면 예상할 수 있는 결. 아형이 주도적인 역할을 하는지 혹은 각 아형별로 다른. 과이며 본 실험은 이를 확인하고 이를 바탕으로 PKC. 경로를 통하여 나름대로 심근 보호에 기여하는지 등이. 실험에서 비슷한 생존을 나타내는 허혈 조건 즉 시간을. 아직 확실하지 않다. 특히 PKC는 쥐의 경우 태생기에서. 결정하였다.. 성체로 성장하면서 심근세포내의 각종 아형 함량이 크. 신생쥐는 허혈 24시간이 지나, 성체쥐는 6시간이 지. 게 변화하므로 허혈시 이에 대한 보호 작용도 영향을 받. 나 생존율이 절반 이하로 떨어지므로 각각 이 조건하에. 13). 그러나 성장에 동반된 PKC의 아. 서 PKC 활성화에 따른 생존율 차이를 관찰하였다. 결. 형 변화가 실제 허혈 발생시 심근세포의 생존에 변화를. 을 가능성이 많다.. 과에서 신생쥐는 TPA 투여로 인한 PKC 활성화가 심. 가져올지를 직접 관찰한 연구는 이제까지 보고된 바가. 근세포의 생존을 전혀 높이지 못한 반면, 성체쥐에서는. 없다. 따라서 본 연구는 이를 확인할 목적으로 고안되었. 심근세포의 생존율이 현저하게 향상되어 허혈을 가하지. 으며 생후 1~2일된 신생쥐와 성장한 성체쥐에서 심근세. 않은 대조군과 동일한 수준을 나타내었다. 성체쥐의 심. 포를 분리하고 이들에게 허혈을 부과한 후 PKC의 활성. 근세포가 전체적으로 허혈에 대해 신생쥐에 비해 많이. 도를 변조하여 그에 따른 세포의 생존, 사멸 양식에 차이. 취약하였음에도 불구하고 PKC 활성화에 의한 심근 보. 가 있는지를 보고자 하였다.. 호 효과는 성체쥐에서 매우 두드러지게 나타났다. 또한. 먼저 허혈 상태에서의 양쪽 세포의 생존율을 비교하. staurosporin을 사용하여 PKC를 억제한 경우에도 신. 였다. 비록 대상 동물은 같은 종이지만 신생쥐와 성체쥐. 생쥐에서는 심근세포의 생존에 큰 차이가 없었던 반면. 에서 체외 실험으로 분리된 세포는 그 분리과정이나 배. 허혈 상태의 성체쥐 심근세포는 다수가 사망하여 생존. 양 등이 다르므로 허혈에 대한 세포의 반응 혹은 예민. 율이 0%로 떨어졌으며 이 같은 감소 추세는 허혈을 가. 694. Korean Circulation J 2002; 32(8):689-696.
(7) 하지 않은 대조군에서도 역시 관찰되었다. 이러한 소견. 이나 형질 변환 동물을 이용하는 것이 유용한 수단이 될. 들은 그 원인은 아직 정확하게 알려져 있지 않으나 신. 것으로 생각된다. 근래에 Dorn 등24)은 PKCε 아형의. 생쥐의 심근세포는 생존에 필요한 필요한 보호장치를 따. 수용체 단백으로 PKCε의 특이 활성제제인 psiepsilon. 로 가지고 있는데 비해 성체쥐 심근세포는 생존을 유지. RACK(receptor for activated C kinase)을 과발현시. 하는데 PKC에 매우 의존하고 있음을 나타낸다. 성체. 킴으로써 생체에서의 허혈에 대한 심근 보호 작용이 있. 쥐에서 PKC의 심근 보호 효과는 특히 허혈 상태에서. 음을 관찰하였다.. 괄목하게 나타났으나 PKC 억제시 허혈 상태가 아닌 세. 결론적으로 본 연구는 허혈 상태에서 배양된 신생쥐와. 포들의 생존율이 감소하는 소견은 허혈 뿐 아니라 다른. 성체쥐의 심근세포에서 PKC 활성화에 따른 생존에 유. 자극에 의한 세포 손상도 보호하는 기능이 있음을 시. 의한 차이가 있음을 관찰하였으며 이는 아마도 성장에. 사한다.. 따른 PKC 아형 변화에 의해서 영향을 받았을 것으로 추. 본 연구에서 허혈에서 PKC의 활성으로 인한 심근 보. 정하였다. 향후 심근 보호에서 주 역할을 하는 PKC의 아. 호 효과는 개체가 성장하면서 크게 상승함을 보여 주었. 형의 규명과 이를 근거로 한 허혈성 전처치의 신호전달. 으며 이는 성장에 따른 PKC 아형의 변화에 기인하였. 체계를 알아내어 세포의 생존을 향상시키는 기전을 밝히. 을 가능성이 많다. 그러나 본 실험에서의 PKC 활성화. 는 것이 의미있는 연구과제가 될 것으로 생각된다.. 제제인 TPA는 atypical PKC(aPKC)를 제외한 모든. 요. PKC를 활성화시키며 staurosporin은 전체 PKC에 대. 약. 해 비특이적인 억제 작용을 나타내므로 PKC의 어떤 아 형이 성체쥐의 심근 보호 효과를 주로 매개하였는지의. 배경 및 목적:. 여부는 알기 어렵다. PKC 아형 중 성체 심근 조직 내에. Protein kinase C(PKC)는 허혈에 대한 세포의 방어. 가장 많이 함유된 것은 ε아형으로 알려져 있고22) PKCε. 기전에 중요한 역할을 하며 PKC의 아형은 세포내 함량. 아형의 보호 효과는 여러 실험을 통하여 보고된 바 있. 은 신생 백서에서 성체에 이르는 성장 과정에서 현저히. 11)12). 현재로서는 PKCε가 본 실험 결과에서 나타난. 변한다고 알려졌다. 본 연구는 성장에 따른 아형 변환에. 성체쥐와 신생쥐의 차이를 설명할 가장 유력한 후보 물. 의해서 실제 허혈 상태에서 PKC 활성이 심근세포 생존. 질이다. 그러나 PKCδ 아형 역시 성체쥐의 심근 조직. 향상에 다른 효과를 나타내는지를 보고자 하였다.. 에 존재하며 일부의 실험에서는 PKCd가 약제나 허혈. 방 법:. 어. 성 전처치에 의한 심근 보호작용을 중재한다고 보고하 10)23). 고 있어. Sprague-Dawley 종의 생후 1일된 신생쥐와 성체쥐. 이의 역할에 대해 향후 규명이 필요한 분. 의 심근세포를 효소 소화법에 의해 1차 분리 배양한 후. 야이다. PKCα는 허혈시 세포내 이동이 관찰되어 보호. 95% N2와 5% CO2의 혼합기체를 주입하고 배지내 de-. 9). 기능의 중재자일 가능성은 있으나 심근내의 함량에 대. xtrose를 deoxy-dextrose로 대치하여 허혈을 조성하. 해서는 일치된 견해가 없다. 즉 PKCα는 신생쥐에 비해. 였다. PKC의 보호 효과를 알기 위해 효현제(TPA 100. 성체쥐에서 세포당 함량이 증가한다는 주장이 있는 반. nM)와 길항제(staurosporin 100 nM)을 사용하였으. 13). 면. 성체쥐 심근 조직에서의 함량은 작으며 그나마 주. 로 섬유모세포(fibroblast)와 같은 비심근세포에서 발견. 며 세포 생사는 trypan blue 염색으로 관측하였다.. 결 과:. 된다는22) 상이한 보고도 있다. 따라서 PKCα 아형의 역. 신생쥐는 허혈 24시간에 성체쥐는 허혈 6시간 이후. 할은 좀더 불확실한 것으로 사료된다. PKC 아형의 역할. 심근세포의 과반수가 사망하였다. 신생쥐의 경우 24시간. 에 대한 평가는 각각의 아형에 대한 특이적(specific). 허혈후 31.8±5.8%의 세포가 생존하였으며 TPA에 의. 인 활성제제나 억제제를 사용하면 얻을 수 있겠지만 현. 한 생존 증가는 없었다(28.5±7.3%). 성체쥐 심근세포. 8)10). 재까지는 일부를 제외하고. 특이 제제가 개발되어 있. 는 허혈 6시간 이후 31.8±5.8%의 세포가 생존하였으나,. 지 않아서 그러한 연구를 시행하는데 제한이 되고 있다.. TPA 투여후 생존은 현저히 증가(62.1±7.3%)하였. PKC 아형의 효과에 대한 직접적인 평가를 하기 위해서. 으며 허혈 조치를 하지 않은 세포의 생존(65.4±6.2%). 는 특정 아형의 adenoviral vector를 통한 유전자 전달. 과 유사하였다. 695.
(8) 11) Gray MO, Karliner JS, Mochly-Rosen D. A selective e-. 결 론: 분리된 심근세포는 성체쥐가 신생쥐에 비해 허혈에 취 약하였으나, protein kinase C 활성화에 따른 생존향상 은 성체쥐에서 현저히 나타난데 비해 신생쥐에서는 미. 12). 약하였다.. 중심 단어:Protein kinase C;허혈성 전처치;심근. 13). 세포.. REFERENCES. 14). 1) Fibrinolytic Therapy Trialist’ (FTT) Collaborative Group.. 2). 3). 4). 5). 6) 7). 8). 9). 10). Indications for fibrinolytic therapy in suspected acute myocardial infarction: collaborative overview of early mortality and major morbidity from all randomized trials of more than 1000 patients. Lancet 1994;343:311-22. Weaver WD, Simes RJ, Betriu A, Grines CL, Zijlstra F, Garcia E, Grinfeld L, Gibbsons RJ, Ribeiro EE, DeWood MA, Ribichini F. Comparison of primary coronary angioplasty and intravenous thrombolytic therapy for acute myocardial infarction: a quantitative review. JAMA 1997;278:2093-8. Rogers WJ, Bowlby LJ, Chandra NC, French WJ, Gore JM, Lambrew CT, Rubison RM, Tiefenbrunn AJ, Weaver WD. Treatment of myocardial infarction in the United States (1990 to 1993). Circulation 1994;90:2103-14. Reimer KA, Murry CE, Yamagawa I, Hill ML, Jennings RB. Four brief periods of ischemia cause no cumulative ATP loss or necrosis. Am J Physiol 1986;251:H1306-15. Murry CE, Jennings RB, Reimer KA. Preconditioning with ischemia: a delay of lethal cell injury in ischemic myocardium. Circulation 1986;74:1124-36. Baxter GF. Ischaemic preconditioning of myocardium. Ann Med 1997;29:345-52. Nishizuka Y. Intracellular signalling by hydrolysis of phospholipids and activation of protein kinase C. Science 1992;258:607-14. Steinberg SF, Goldberg M, Rybin VO. Protein kinase C isoforms diversity in the heart. J Mol Cell Cardiol 1995; 27:141-53. Goldberg M, Zhang HL, Steinberg SF. Hypoxia alters the subcellular distribution of protein kinase C isoforms in neonatal rat ventricular myocytes. J Clin Invest 1997; 99:55-61. Inagaki K, Kihara Y, Hayashida W, Izumi T, Iwanaga Y, Yoneda T, Takeuchi Y, Suyama K, Muso E, Sasayama S. Anti-ischemic effect of a novel cardioprotective agent, JTV519, is mediated through specific activation of disoform of protein kinase C in rat ventricular myocardium. Circulation 2000;101:797-804.. 696. 15). 16). 17). 18). 19) 20) 21). 22). 23). 24). protein kinase C antagonist inhibits protection of cardiac myocytes from hypoxia-induced cell death. J Biol Chem 1997;272:30945-51. Miyamae M, Rodriguez MM, Camacho SA, Diamond I, Mochly-Rosen D, Figueredo VM. Activation of epsilon protein kinase C correlates with a cardioprotective effect of regular ethanol consumption. Proc Natl Acad Sci USA 1998;95:8262-7. Clerk A, Bogoyevitch MA, Fuller SJ, Lazou A, Parker PJ, Sugden PH. Expression of protein kinase C isoforms during cardiac ventricular development. Am J Physiol 1995;269:H1087-97. Park SK, Kim HS, Sohn CS, Dokko YC, Jeon YH. Expression of protein kinase C isoform mRNAs in the developing rat heart. Korean Circ J 1998;28:1341-9. Berger HJ, Prasad SK, Davidoff AJ, Pimental D, Ellingsen O, Marsh JD, Smith TW, Kelly RA. Continued electric field stimulation preserves contractile function of adult ventricular myocytes in primary culture. Am J Physiol 1994;266:H341-9. Volz A, Piper HM, Sigmund B, Schwartz P. Longevity of adult ventricular rat heart muscle cells in serum-free priamry culture. J Mol Cell Cardiol 1991;23:161-73. Springhorn JP, Claycomb WC. Preproenkephalin mRNA expression in developing rat heart and in cultured ventricular cardiac muscle cells. Biochem J 1989;258:73-8. McGahon AJ, Martin SJ, Bissonnette RP, Mahboubi A, Shi Y, Mogil RJ, Nishioka WK, Green DR. The end of the cell line: methods for the study of apoptosis in vitro. Methods Cell Biol 1995;46:153-85. Parker PJ. Protein kinase C and its relatives. In: Woodget JR, editor. Protein kinases. Oxford; IRL press;1996. p.68-84. Ruoslahti E. Stretching is good for a cell. Science 1997; 276:1345-6. Koh JS, Lieberthal W, Heydrick S, Levine S. Lysophosphatidic acid is a major serum noncytokine survival factor for murine macrophages which acts via the phosphatidylinositol 3-kinase signalling pathway. J Clin Invest 1998; 102:716-27. Rybin VO, Steinberg SF. Protein kinase C isoform expression and regulation in the developing rat heart. Circ Res 1994;74:299-309. Zhao J, Renner O, Wightman L, Sugden PH, Stewart L, Miller AD, Latchman DS, Marber MS. The expression of constituitively active isotype of protein kinase C to investigate preconditioning. J Biol Chem 1998;273:23072-9. Dorn GW 2nd, Souroujon MC, Liron T, Chen CH, Gray MO, Zhou HZ, Csukai M, Wu G, Lorenz JN, MochlyRosen D. Sustained in vivo cardiac protection by rationally designed peptide that causes epsilon protein kinase C translocation. Proc Natl Acad Sci USA 1999;96:12798-803.. Korean Circulation J 2002; 32(8):689-696.
(9)
수치
관련 문서
나는 받는 사람에게 가장 좋은 패션상품을 선물하기 위해 여기 저기 돌아다 9 니며 쇼핑하는 것을 좋아한다.. 10 내가 원하는 패션상품을 사기 위해
First, same words undergo the same phonological phenomena such as vowel shortening(diphthongs become simple vowels by glide deletion), tensing and
문제란 현실과 원하는 것의 차이-카우프만(kaufmann) 목표와 현상과의 차이(Gap) 이고 해결해야할 사항 바람직한 모습과 실제
손상에 대한 즉각적인 반응으로부터 수복(치유)에 이르는 일련의 형태 학적 및 기능적인 모든 과정.. 염증의
낯선 상황의 의미에 대한 문화적 차이.. 문화와 애착. 애착
[r]
분열지연중심부 (quiscent centre) : 분열조직 손상에 대비한 대체 저장... 표피 (epidermis) :
- 미국 화장품 안전성은 미국 식품의약청(FDA)이 ‘연방 식품, 의약품, 화장품법 (Federal Food, Drug and Cosmetic Act)’에 의거해 규제.. -
