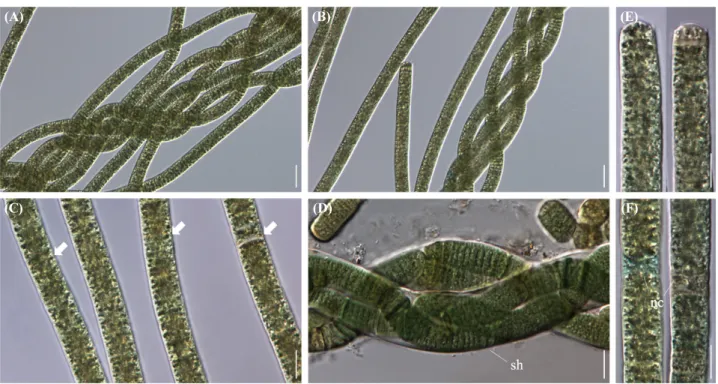
Two strains of Aerosakkonema were isolated from the estuary of Geum River on August 2019. The cells were measured at 3.0~4.1 µm in length and 12.0~15.0 µm in width, larger than taxa belonging to similar genera. Trichomes were either straight or twisted like a rope, and necridic cells were observed in the trichome.
The 16S rRNA gene sequences of the Korean Aerosakkonema were completely identical (100% similarity) to the type species A. funiforme. Phylogenetic analysis of the 16S rRNA gene sequences showed that the Korean Aerosakkonema was clustered with the A. funiforme. Additional analyses by using gvpA and rpoB gene sequences supported that it belonged to the genus Aerosakkonema and/or A. funiforme. Through these results, the Korean Aerosakkonema was identified to A. funiforme, and it was unrecorded genus and species in Korea.
Keywords: Aerosakkonema funiforme, cyanobacteria, freshwater algae, newly recorded genus and species
남세균은 광합성을 통하여 산소를 생성하는 원핵생물로서 담수와 해수에 분포하며, 부유하거나 기질에 부착하여 살아간
다. 또한 기중환경이나 공생관계를 형성하는 등 극한의 환경 속에서도 분포한다(Komárek and Anagnostidis, 1998; Whitton and Potts, 2000).
흔들말목(Oscillatoriales)은 사상체(filament or trichome) 를 형성하고, 이형세포(heterocyte)와 아키네트(akinete) 같은 특수한 세포를 가지지 않으며, 주로 점액질 초(sheath)의 유무 에 따라 분류된다. 또한 점액질 초의 형태, 사상체의 특징, 위분 지(false branching) 방식, 세포의 색 등이 분류를 위한 형태적 형질로 사용된다(Komárek and Anagnostidis, 2005).
최근에는 남세균의 분류에 있어서 형태적 형질뿐만 아니라 전자현미경을 사용한 미세구조, 16S rRNA 유전자 염기서열과 같은 분자적 형질, 그리고 생태적 형질 등을 도입하여 다각도로 분류하고 있다(Komárek et al., 2014; Komárek, 2016). 흔들말 목에 속하는 주요 속인 Phormidium, Microcoleus, Oscillatoria 등은 16S rRNA 유전자 염기서열에 의한 계통 분석 결과에서 다계통 분류군으로 나타남으로써, 신속과 신종들이 분리, 보 고되었다(Casamatta et al., 2012; Chatchawan et al., 2012).
열대지방에서 채집된 Phormidium으로부터 Phormidesmis 가 분리되었고(Komárek et al., 2009), 남극에서 출현한 시료 를 바탕으로 Wilmottia와 W. murrayi (West & G.S.West) Strunecký, Elster & Komárek가 새로운 속과 종으로 명명되었 다(Strunecký et al., 2011). Oscillatoria에서는 초미세구조적 형
담수 남세균, Aerosakkonema funiforme (Oscillatoriales, Cyanobacteria)의 국내 미기록속 및 미기록종에 대한 연구
김도현
1
・ 윤성로1
・ 김태희2
・ 기장서2
* ・ 이옥민1
*1경기대학교 생명과학과, 2상명대학교 생명공학과
Newly recorded genus and species for freshwater cyanobacteria Aerosakkonema funiforme (Oscillatoriales, Cyanobacteria) in Korea
Do-Hyun Kim
1
, Sung-Ro Yoon1
, Taehee Kim2
, Jang-Seu Ki2
* , and Ok-Min Lee1
*1Department of Life Science, College of Natural Science, Kyonggi University, Suwon 16227, Republic of Korea
2Department of Biotechnology, Sangmyung University, Seoul 03016, Republic of Korea
(Received May 11, 2020; Revised July 7, 2020; Accepted July 9, 2020)
*For correspondence. (J.S. Ki) E-mail: [email protected];
Tel.: +82-2-2287-5449; Fax: +82-2-2287-0070 / (O.M. Lee) E-mail: [email protected];
Tel.: +82-31-249-9643; Fax: +82-31-241-0860
질 차이, 16S rRNA 유전자 염기서열 차이 등으로 Planktothrix 와 Planktothricoides가 분리되었다(Komárek and Komárková, 2004).
흔들말목에 속하는 Aerosakkonema는 라오스의 중영양수 역에서 채집된 종으로, 전세계적으로 1종이 보고되어 있다 (Thu et al., 2012). 이 연구에서는 gas vesicle structural protein A (GvpA) 유전자를 비교하였고, 16S rRNA 유전자 염기서열 을 사용하여 계통 분석을 실시하였다. A. funiforme Thu &
Watanabe는 형태적으로 유사한 속들에 비해 세포의 크기가 크며, 더 작은 크기의 가스 소포(gas vesicle)를 가진다(Thu et al., 2012). 또한 TEM을 사용한 연구에서 틸라코이드가 불규 칙한 배열을 나타냄으로써 형태적 차이를 나타냈다(Thu et al., 2012).
현재 남세균은 전세계적으로 4,729종이 보고되었으며, 흔 들말목은 1,299종이 보고되었다(Guiry and Guiry, 2020). 국 내에서는 343분류군의 남세균이 보고되었으며, 흔들말목은 151분류군이 포함되어 있다(Kim, 2015). 최근에는 남세균에 대한 국내 미기록종이 지속적으로 보고되고 있다. 한강과 낙 동강의 담수와 기중 서식처를 대상으로 미기록속인 Capsosira 와 6종의 미기록종이 보고되었다(Song and Lee, 2017). 또한 국내 미기록속인 Borzia와 6종의 미기록종이 낙동강의 담수 및 기수 서식처로부터 보고되었다(Yim et al., 2017). Wilmottia murrayi는 형태적 형질, 초미세구조 그리고 16S rRNA 유전자 염기서열을 분석하여 국내 미기록속 및 미기록종으로 보고된 바 있다(Lee et al., 2019).
본 연구에서는 금강수계의 하류에 위치한 금강갑문의 담수 역으로부터 남세균을 채집하여 단일배양을 실시하였고, 광학 현미경과 투과전자현미경을 사용하여 형태적 형질을 관찰하 였다. 또한 16S rRNA 유전자 염기서열, 단백질을 코딩하는 GvpA 그리고 RNA polymerase β-subunit (rpoB) 유전자 염기 서열을 이용하여 계통분석을 실시하였다. 이를 통하여 한국산 남세균의 국내 미기록속 및 미기록종을 보고하고자 하였다.
재료 및 방법
채집지 및 채집방법
본 연구의 시료는 2019년 8월 금강수계의 하류에 위치한 금 강갑문(36°01'26.0"N,126°46'28.7"E) 담수역에서 채집되었 다. 부유성 남세균의 채집은 망목의 직경이 25 µm인 플랑크톤 네트를 사용하였으며(Sournia, 1978), 부착성 남세균은 식물, 바위 등 기질의 표면을 긁어 채집하였다. 채집된 시료들은 현
장에서 밀봉 후, 4°C의 아이스박스에 보관하여 실험실로 운반 되었다.
실내배양
단일배양을 위해 광학현미경 하에서 파스퇴르 피펫을 이용 하여 사상체 한가닥을 뽑아 1.2% agar를 넣은 고체배지에 올 려 배양하였고, 배양된 사상체 다발을 24-well plate (SPL)에 넣어 분리하였다. 24-well plate에서 1~2주간 배양 후, 오염여 부를 확인하고, 사상체 다발을 50 ml cell culture flask (SPL)에 옮겨 대량배양을 실시하였다(Lee et al., 2019). 단일배양 및 대 량배양은 BG-11 배지(Stanier et al., 1971)를 사용하였다. 실 내배양은 20~25°C, 명암 주기 16 h: 8 h, 조도는 25 µmol/m 2 /s 의 조건 하에서 실시되었다. 단일배양 된 2주의 배양체는 ACKU (Algae Culture of Kyonggi University)에서 배양하고, 국립낙 동강생물자원관에 기탁하였다(FBCC-A397 = ACKU621, FBCC- A398 = ACKU622).
형태 관찰
본 연구에서는 광학현미경(Axio Imager A2, Carl Zeiss;
Olympus BX53, Olympus)을 이용하여 Aerosakkonema 배양 주 2주(FBCC-A397, FBCC-A398)를 100~1,000 배율로 관찰 하였고, 400~1,000 배율에서 사진촬영을 실시하였다(AxioCam HRC camera, Carl Zeiss; an Olympus UC-90, Olympus). 남세 균의 분류체계는 Komárek 등(2014)에 따랐고, AlgaeBase를 참조하였다(Guiry and Guiry, 2020). 또한 남세균의 동정에는 Komárek and Anagnostidis (2005), Thu 등(2012) 그리고 da Silva Malone 등(2015)을 참조하였다.
DNA 추출
배양된 시료에서 1 ml을 1.5 ml 튜브에 옮겨 담아 13,000 rpm에서 10분간 원심분리를 하였다. 원심분리 후 상등액을 제 거하고, 호모게나이저를 이용하여 세포를 파쇄하였다. 이 후 i-genomic Plant DNA Extraction Mini Kit (iNtRON)를 사 용하여 genomic DNA (gDNA)를 추출하였다.
효소중합연쇄반응(PCR) 및 DNA 시퀀싱
PCR에 사용된 16S 프라이머는 27F1 (Neilan et al., 1997)과 23S30R (Taton et al., 2003)이고, gvpA 유전자는 GVPAF와 GVPAR이다(Beard et al., 1999). rpoB 유전자 증폭은 신규로 디자인한 Aerosakkonema 특이적 프라이머(Aero-rpob-F980:
5'-ATC CGG GAA CGG ATG ACC GT-3'; Aero-rpob-R1970:
5'-TAG CCT TCC CAA GGC ATA TA-3')를 이용하였다. 유전 자 증폭을 위해 Maxime TM PCR PreMix Kit (i-StarTaq TM GH) (iNtRON)에 멸균한 3차 증류수 17 µl와 추출한 gDNA (1 µl), 각각의 프라이머(10 pmole, 1 µl)를 포함하여 총 20 µl를 혼합 하여 PCR 튜브를 조성하였다. 조성된 PCR 튜브를 Mastercycler gradient (Eppendorf) 기기에 넣어 반응조건을 입력 후 PCR을 진행하였다.
PCR 반응은 초기 94°C, 5분간 DNA를 변성시킨 후, 94°C 1 분, 55°C 2분, 72°C 3분을 33회 반복하여 대상 유전자 영역을 증폭하였다. PCR 증폭 반응이 종료되면 추가로 72°C, 10분간 유지한 후 반응을 종결하였다. 증폭된 PCR 산물은 1% agarose gel에서 전기영동하여 관찰하였다.
DNA sequencing은 PCR 산물을 MEGAquick-spin TM Plus DNA Fragment Purification Kit (iNtRON)로 정제하였다. 정 제된 PCR 생성물의 DNA 염기서열을 ABI 3730XL automated sequencer (Applied Biosystems)로 분석하였다.
염기서열 유사도, 유전 거리, 계통 분석
염기서열 분석을 통해 얻어진 뉴클레오타이드 시퀀스는 SeqMan 프로그램(DNASTAR)을 사용하여 불명확한 말단 부분 을 제거하였다. 본 연구에서 규명한 염기서열을 NCBI (National Center for Biotechnology Information)의 Nucleotide BLAST (http://blast.ncbi.nlm.nih.gov/Blast.cgi)에 입력 후 유전적 거 리가 비교적 가까운 염기서열 정보를 수집하였다. 본 연구에 서 얻은 남세균의 염기서열과 NCBI 데이터베이스를 통해 수 집된 염기서열은 MEGA6.0 (Tamura et al., 2013)를 이용하여 염기서열을 재배열한 후, 같은 길이가 되도록 양 끝을 제거하 였다. 이후 BioEdit (http://www.mbio.ncsu.edu/bioedit/bioedit.
html)를 이용하여 유사도를 계산하고, MEGA 6.0에서 Kimura 2-parameter 모델을 이용하여 유전 거리를 계산하였다(Kimura, 1980).
본 연구에서 채집된 FBCC-A397과 GenBank로부터 얻은 총 37주의 16S rRNA 유전자 염기서열을 이용하여, Maximum- Likelihood (ML)과 Neighbor-Joining (NJ) 계통 분석을 실시 하였다. ML 계통수는 General Time Reversible (GTR)을 이용 하였고, NJ 계통수는 Kimura 2-parameter 모델을 이용하여 MEGA 6.0에서 추정하였다. 추가로 gvpA와 rpoB 유전자의 계통분석을 실시하였다.
초미세구조 관찰
초미세구조는 TEM (transmission electron microscope)을
이용하여 분석하였다. 원심분리기를 이용하여 시료를 모은 후, Glutaraldehyde 2%, Paraformaldehyde 2% 용액을 사용하여 고정하였다. 후고정은 Osmium tetroxide 2% 용액으로 1시간 동안 고정하였다. Ethanol 50%, 60%, 70%, 80%, 90%, 100%
에 넣어 각 20분씩 탈수를 진행하였으며, 에탄올과 propylene oxide를 각각 2:1, 1:1, 1:2에서 총 3번, 각 20분씩 치환을 실시 한 뒤, 마지막으로 propylene oxide 100%에서 20분간 치환하 였다. 포매는 Propylene oxide와 Epon-812을 2:1, 1:1, 1:2에서 총 3번 각 20분씩 진행하였다. 마무리로 Epon-812 100%에서 20분 동안 포매하였다. 실험이 진행된 시료를 60°C에서 48시 간 동안 정치하였다. 이후 초박절편을 하여 grid에 올려 JEM- 2100F (Jeol)을 사용하여 관찰하였고, OneView camera (Gatan) 소프트웨어를 통하여 초미세구조 촬영을 하였다(Kim et al., 2015).
결 과
본 연구에서는 2019년 8월에 금강수계의 금강갑문의 담수 역에서 Aerosakkonema로 추정되는 남세균을 채집하여 배양 주 2주를 단일배양 하였다. 종 동정은 세포의 형태적 특징을 사 용하였고, 초미세구조 관찰을 통하여 흔들말과에 속하는 것을 확인하였다. 2주의 배양주는 16S rRNA 유전자 염기서열과 GvpA 단백질 아미노산 서열을 분석하여 A. funiforme로 동정 되었다. 또한 이들은 rpoB 유전자 염기서열 분석을 통하여 다 른 속들과는 분리됨을 확인하였다. 이상의 연구를 통하여 금 강갑문에서 출현한 Aerosakkonema funiforme Thu & Watanabe 2012는 국내 미기록속 및 미기록종으로 추가되었다.
형태적 특징
Family Oscillatoriaceae Engler 1898
Genus Aerosakkonema Thu & Watanabe 2012: 사상체는 청록 색 또는 황록색의 색을 띠며, 단일가닥 이거나 또는 몇 개의 가 닥이 다발을 이루기도 하고, 자유롭게 부유한다. 사상체는 대 부분 곧게 뻗어 있으며, 간혹 굽어 있기도 한다. 세포간 격벽은 함입 되어있지 않거나, 간혹 약하게 함입되기도 한다. 말단세 포(apical cell or terminal cell)로 갈수록 폭이 좁아지지 않으나 드물게 좁아지기도 한다. 점액질 초는 매우 얇게 나타나기도 한다. 틸라코이드는 불규칙적인 배열을 한다. 분열은 괴사세 포(necridic cell)에 의해 이루어진다.
Aerosakkonema funiforme Thu & Watanabe 2012 (Fig. 1): 사
상체는 청록색 또는 황록색의 색을 띠며, 단독이거나 다발을
이루고, 자유롭게 부유한다. 때로는 밧줄처럼 꼬여있다. 사상 체는 곧게 뻗어 있으며, 약하게 굽어 있기도 하다. 세포 내 격 벽은 함입 되어있지 않고, 간혹 약하게 함입되기도 한다. 말단 세포는 둥글고, 칼립트라(calyptra)가 존재하지 않는다. 점액 질 초는 거의 없으나 배양 시 매우 얇게 보이기도 한다. 위분지 가 일어나지 않으며, 이형세포와 아키네트를 형성하지 않는 다. 분열은 괴사세포에 의해 이루어진다. 세포의 길이는 3.0~
4.1 µm, 폭은 12.0~15.0 µm이며, 세포의 길이보다 폭이 넓은 형태이다.
Ecology: 금강갑문 수역의 부유성 종으로 출현하였고, 본 수역 은 중영양-부영양수역이다(NIER, 2020).
Distribution: 라오스인민민주공화국(Thu et al., 2012) Site of collection: 전북 군산시 성산면 성덕리(36°01'26.0"N, 126°46'28.7"E)
Date of collection: 2019년 08월 26일
Specimen Locality: FBCC- A397, FBCC- A398
염기서열 특성 및 분자 계통
본 연구에서 남세균 Aerosakkonema funiforme의 배양주 2주로부터 1,330 bp (MN911406)와 1,315 bp (MN911419)의 16S rRNA 유전자 염기서열과 1,068 bp의 rpoB 유전자 염기서
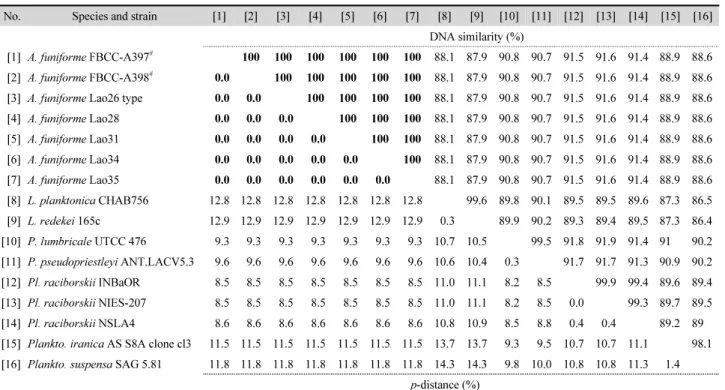
열을 규명하였으며(Table 1), 2주의 배양주는 100% 동일한 서 열을 갖는 것으로 파악되었다. 또한 Aerosakkonema 배양주 (FBCC-A397)로부터 gvpA 유전자 염기서열 140 bp를 규명하 였다. 16S rRNA 유전자 염기서열 유사도와 유전거리 분석에 서 기존에 보고된 A. funiforme와 100% 일치하는 것을 확인하 였다(Table 2).
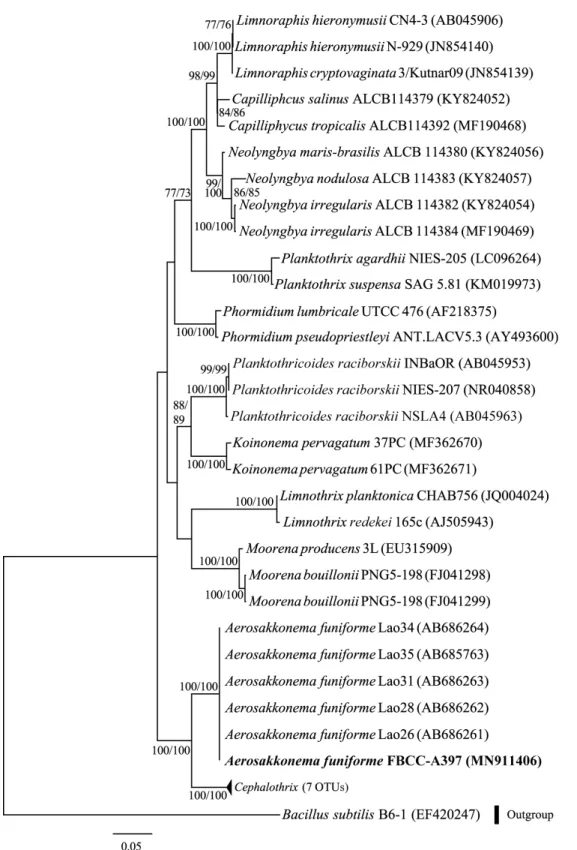
NCBI에 등록된 남세균 A. funiforme와 유사한 형태를 가진 종들을 포함하여 총 37주의 16S rRNA 유전자 염기서열(Table 1)을 이용하여 ML tree를 추정하였다(Fig. 2). 동일한 데이터 를 이용한 Neighbor-joining (NJ) 분석에서 유사한 branch 양 상을 나타냈다. 한국의 Aerosakkonema는 기존에 보고된 A.
funiforme (AB686261-AB686264)와 같은 분계(clade)를 형성 하였다.
추가적인 GvpA 단백질 아미노산 서열을 이용한 ML tree 분 석에서 한국의 Aerosakkonema는 기존에 보고된 A. funiforme 가 같은 분계를 형성하였다(Fig. 3A). 또한, rpoB 유전자를 사용한 남세균 흔들말과에 속하는 분류군의 ML tree 분석 에서 A. funiforme 2주의 배양주가 같은 분계를 형성하였으 며, Oscillatoria nigro-viridis와 자매 관계로 파악되었다(Fig.
3B).
(A) (B)
(D)
(E)
(F) (C)
Fig. 1. Microscopic photographs of Aerosakkonema funiforme strain ACKU621 (FBCC-A397). (A, B) Trichomes twisted into a rope, (C) Gas vesicles
(arrows) scattered inside the trichomes, (D) Old trichomes with sheaths (sh), (E) Apical cells, (F) Trichomes with necridic cells (nc), Scale bars = (A, B) 20
µm, (C-F) 10 µm.
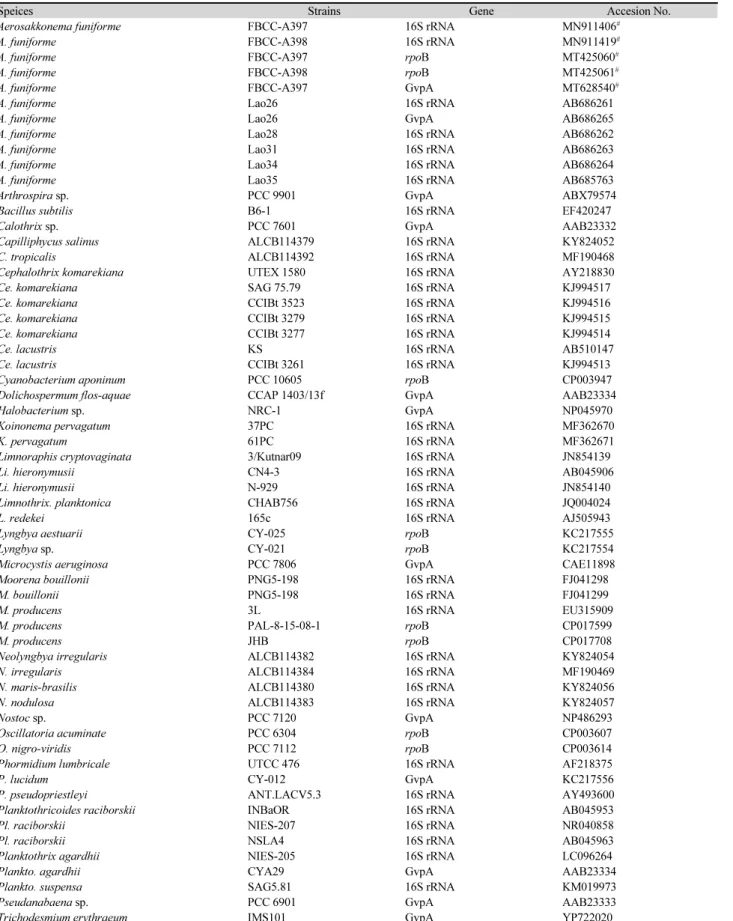
Table 1. List of species, genes and their GenBank accession numbers used in this study
Speices Strains Gene Accesion No.
Aerosakkonema funiforme FBCC-A397 16S rRNA MN911406
#A. funiforme FBCC-A398 16S rRNA MN911419
#A. funiforme FBCC-A397 rpoB MT425060
#A. funiforme FBCC-A398 rpoB MT425061
#A. funiforme FBCC-A397 GvpA MT628540
#A. funiforme Lao26 16S rRNA AB686261
A. funiforme Lao26 GvpA AB686265
A. funiforme Lao28 16S rRNA AB686262
A. funiforme Lao31 16S rRNA AB686263
A. funiforme Lao34 16S rRNA AB686264
A. funiforme Lao35 16S rRNA AB685763
Arthrospira sp. PCC 9901 GvpA ABX79574
Bacillus subtilis B6-1 16S rRNA EF420247
Calothrix sp. PCC 7601 GvpA AAB23332
Capilliphycus salinus ALCB114379 16S rRNA KY824052
C. tropicalis ALCB114392 16S rRNA MF190468
Cephalothrix komarekiana UTEX 1580 16S rRNA AY218830
Ce. komarekiana SAG 75.79 16S rRNA KJ994517
Ce. komarekiana CCIBt 3523 16S rRNA KJ994516
Ce. komarekiana CCIBt 3279 16S rRNA KJ994515
Ce. komarekiana CCIBt 3277 16S rRNA KJ994514
Ce. lacustris KS 16S rRNA AB510147
Ce. lacustris CCIBt 3261 16S rRNA KJ994513
Cyanobacterium aponinum PCC 10605 rpoB CP003947
Dolichospermum flos-aquae CCAP 1403/13f GvpA AAB23334
Halobacterium sp. NRC-1 GvpA NP045970
Koinonema pervagatum 37PC 16S rRNA MF362670
K. pervagatum 61PC 16S rRNA MF362671
Limnoraphis cryptovaginata 3/Kutnar09 16S rRNA JN854139
Li. hieronymusii CN4-3 16S rRNA AB045906
Li. hieronymusii N-929 16S rRNA JN854140
Limnothrix. planktonica CHAB756 16S rRNA JQ004024
L. redekei 165c 16S rRNA AJ505943
Lyngbya aestuarii CY-025 rpoB KC217555
Lyngbya sp. CY-021 rpoB KC217554
Microcystis aeruginosa PCC 7806 GvpA CAE11898
Moorena bouillonii PNG5-198 16S rRNA FJ041298
M. bouillonii PNG5-198 16S rRNA FJ041299
M. producens 3L 16S rRNA EU315909
M. producens PAL-8-15-08-1 rpoB CP017599
M. producens JHB rpoB CP017708
Neolyngbya irregularis ALCB114382 16S rRNA KY824054
N. irregularis ALCB114384 16S rRNA MF190469
N. maris-brasilis ALCB114380 16S rRNA KY824056
N. nodulosa ALCB114383 16S rRNA KY824057
Nostoc sp. PCC 7120 GvpA NP486293
Oscillatoria acuminate PCC 6304 rpoB CP003607
O. nigro-viridis PCC 7112 rpoB CP003614
Phormidium lumbricale UTCC 476 16S rRNA AF218375
P. lucidum CY-012 GvpA KC217556
P. pseudopriestleyi ANT.LACV5.3 16S rRNA AY493600
Planktothricoides raciborskii INBaOR 16S rRNA AB045953
Pl. raciborskii NIES-207 16S rRNA NR040858
Pl. raciborskii NSLA4 16S rRNA AB045963
Planktothrix agardhii NIES-205 16S rRNA LC096264
Plankto. agardhii CYA29 GvpA AAB23334
Plankto. suspensa SAG5.81 16S rRNA KM019973
Pseudanabaena sp. PCC 6901 GvpA AAB23333
Trichodesmium erythraeum IMS101 GvpA YP722020
#
Sequences determined in this study
TEM의 초미세구조
세포의 장축 방향 절단면의 틸라코이드의 분포를 확인한 결과, 틸라코이드는 세포 내에서 불규칙한 배열을 나타냈고, 이는 A. funiforme가 속하는 흔들말과의 특징이다(Fig. 4).
고 찰
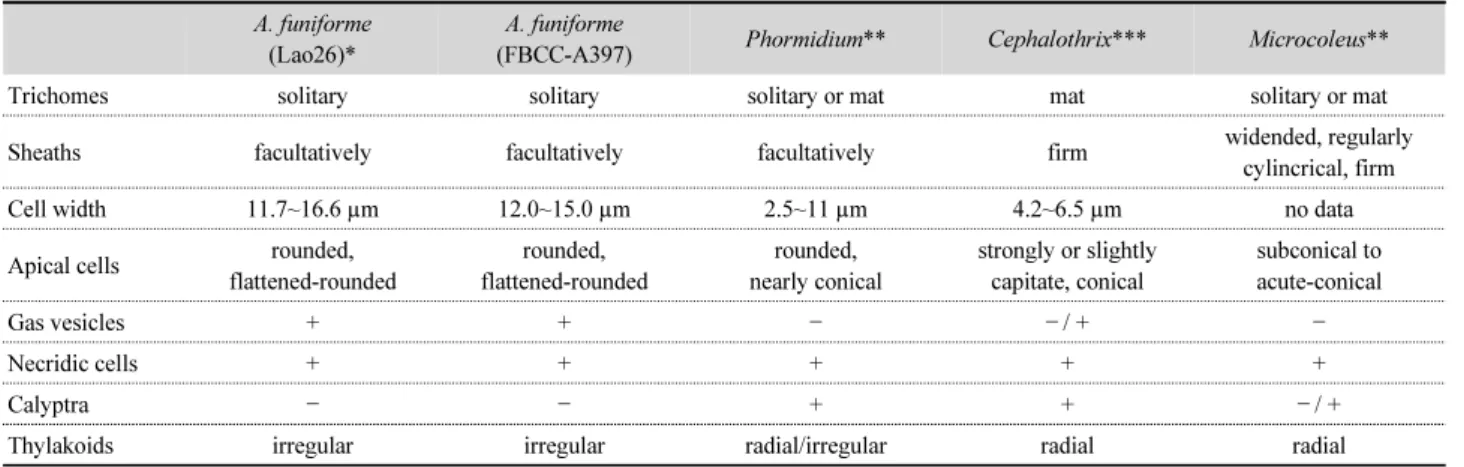
Aerosakkonema와 형태적으로 유사한 속 간의 식별형질은 주로 사상체의 폭과 길이의 크기, 점액질의 유무, 그리고 말단 세포의 형태 등이다(Komárek and Anagnostidis, 2005; da Silva Malone et al., 2015). Aerosakkonema와 같은 과에 속하 는 Phormidium은 형태적으로 가장 유사하지만, 세포의 폭 이 상대적으로 작으며 다양한 형태의 말단세포를 가지는 특 징으로 구분되었다. 또한 형태적으로 유사한 Cephalothrix (Phormidiaceae)는 뚜렷한 사상체의 점액질 초와 두상형(capitate) 의 말단세포를 가지는 점이 다르다. Microcoleus (Microcoleaceae) 는 두꺼운 점액질 초와 원추형의 말단세포를 가지는 특징으로 구분되었다(Table 3).
남세균에서 과(family)를 구분하는 중요한 형질 중 하나는 세포내 틸라코이드의 배열이다(Komárek et al., 2014). 본 연
구의 A. funiforme는 TEM의 미세구조에서 세포내 틸라코이 드가 불규칙한 배열을 나타냈다(Fig. 4). 이는 흔들말과의 특 징으로, 라오스에서 보고된 A. funiforme의 결과와 일치하였 다(Thu et al., 2012).
본 연구의 시료(FBCC-A397)는 16S rRNA 유전자 염기서 열과 GvpA 단백질 서열을 사용한 계통수(ML tree)에서 A.
funiforme의 기준종과 같은 분계를 형성하였다(Figs. 2 and 3A).
또한, 16S rRNA 유전자 염기서열과 GvpA 단백질 서열 유사도 는 A. funiforme (Lao26)와 100%로 일치하였으며, 16S rRNA 유 전자의 유전적 거리는 0.0% 값을 나타냈다. 따라서 본 연구의 시료를 유전적으로 A. funiforme로 동정하였으며, 이는 남세균 에서 16S rRNA 유전자 유사도가 98.65% 이상으로 일치할 경우 동일 종으로 동정하는 결과와 일치하였다(Kim et al., 2014).
현재 NCBI 유전자은행에 Aerosakkonema의 rpoB 유전자 염기서열이 보고되어 있지 않으며, 본 연구에서 처음으로 보고 하는 것이다. rpoB 유전자를 이용한 계통분석에서 A. funiforme (FBCC-A397, FBCC-A398)는 흔들말과의 다른 종들과 뚜렷 하게 분리되었다(Fig. 3B). rpoB 유전자는 16S 유전자보다 변 이가 약 2.4배 높은 해상도를 가지며, 향후 이들 남세균의 유전 학적 분류에 유용하게 활용할 수 있을 것으로 사료된다(Cheon et al., 2012).
Table 2. Similarity scores (above diagonal) and % genetic p-distance (below diagonal) estimated by the Kimura 2-parameter model between 16 pairs of the aligned sequence data of the partial 16S rRNA gene from Aerosakkonema, Limnothrix, Phormidium, Planktothricoides, and Planktothrix
No. Species and strain [1] [2] [3] [4] [5] [6] [7] [8] [9] [10] [11] [12] [13] [14] [15] [16]
DNA similarity (%)
[1] A. funiforme FBCC-A397
#100 100 100 100 100 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [2] A. funiforme FBCC-A398
#0.0 100 100 100 100 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [3] A. funiforme Lao26 type 0.0 0.0 100 100 100 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [4] A. funiforme Lao28 0.0 0.0 0.0 100 100 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [5] A. funiforme Lao31 0.0 0.0 0.0 0.0 100 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [6] A. funiforme Lao34 0.0 0.0 0.0 0.0 0.0 100 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [7] A. funiforme Lao35 0.0 0.0 0.0 0.0 0.0 0.0 88.1 87.9 90.8 90.7 91.5 91.6 91.4 88.9 88.6 [8] L. planktonica CHAB756 12.8 12.8 12.8 12.8 12.8 12.8 12.8 99.6 89.8 90.1 89.5 89.5 89.6 87.3 86.5 [9] L. redekei 165c 12.9 12.9 12.9 12.9 12.9 12.9 12.9 0.3 89.9 90.2 89.3 89.4 89.5 87.3 86.4 [10] P. lumbricale UTCC 476 9.3 9.3 9.3 9.3 9.3 9.3 9.3 10.7 10.5 99.5 91.8 91.9 91.4 91 90.2 [11] P. pseudopriestleyi ANT.LACV5.3 9.6 9.6 9.6 9.6 9.6 9.6 9.6 10.6 10.4 0.3 91.7 91.7 91.3 90.9 90.2 [12] Pl. raciborskii INBaOR 8.5 8.5 8.5 8.5 8.5 8.5 8.5 11.0 11.1 8.2 8.5 99.9 99.4 89.6 89.4 [13] Pl. raciborskii NIES-207 8.5 8.5 8.5 8.5 8.5 8.5 8.5 11.0 11.1 8.2 8.5 0.0 99.3 89.7 89.5 [14] Pl. raciborskii NSLA4 8.6 8.6 8.6 8.6 8.6 8.6 8.6 10.8 10.9 8.5 8.8 0.4 0.4 89.2 89 [15] Plankto. iranica AS S8A clone cl3 11.5 11.5 11.5 11.5 11.5 11.5 11.5 13.7 13.7 9.3 9.5 10.7 10.7 11.1 98.1 [16] Plankto. suspensa SAG 5.81 11.8 11.8 11.8 11.8 11.8 11.8 11.8 14.3 14.3 9.8 10.0 10.8 10.8 11.3 1.4
p-distance (%)
#
Sequences determined in this study. Bolds represent more than 100% similarity and less than 0.0% of genetic distance.
Fig. 2. Phylogenetic relationships of cyanobacteria genus Aerosakkonema. It was inferred from partial 16S rRNA gene sequences with Maximum
Likelihood (ML) algorithm. Additional Neighbor-Joining (NJ) tree generated similar topology of the ML tree, and thus its Bootstrap Proportions (BP) were
incorporated into the tree. The first and second numbers at the nodes display BP (> 50%) in ML and in NJ, respectively. Branch lengths are proportional
to the scale given. Bold represents data obtained in this study. Bold represents a representative Korean isolated, which sequence was completely identical
to those of FBCC-A397.
(A)
(B)
Fig. 3. Phylogenetic relationships of cyanobacteria Korean Aerosakkonema and other species. Each tree was inferred from partial gvpA (A) and rpoB (B) sequence with Maximum Likelihood (ML) algorithm. Additional Neighbor-Joining (NJ) tree generated similar topology of the ML tree, and thus its Bootstrap Proportions (BP) were incorporated into the tree. The first and second numbers at the nodes display BP (> 50%) in ML and in NJ, respectively.
Branch lengths are proportional to the scale given. Bold represents data obtained in this study. Bold represents Korean Aerosakkonema isolates.
(A)
(B)
Fig. 4. Transmission electron micrographs of Aerosakkonema funiforme strain FBCC-A397. (A, B) Longitudinal section. th, thylakoids; cw, cell wall.
적 요
국내 금강수계의 하류에 위치한 금강갑문 담수역으로부터 Aerosakkonema를 채집하여, 2주의 배양주를 확보하였다. 세 포의 길이는 3.0~4.1 µm, 폭은 12.0~15.0 µm로 나타났으며, 유 사 속들에 비해 큰 편이다. 이들 세포는 사상체가 로프처럼 꼬 이는 현상을 나타내며, 사상체 내에 괴사세포가 관찰되었다.
이들의 16S DNA 유사도는 기존에 보고된 A. funiforme와 100%로 일치하였다. 16S rRNA 유전자 염기서열을 사용하 여 계통분석을 실시한 결과, 배양주 2주는 A. funiforme와 같은 분계를 형성하였다. 추가적인 gvpA와 rpoB 유전자 서열을 사 용한 계통분석에서도 본 연구의 배양주는 Aerosakkonema 와 A. funiforme에 포함되는 것을 확인하였다. 따라서 금강갑문 담수역으로부터 채집된 Aerosakkonema 배양주는 A. funiforme 으로 동정되었으며, 국내 미기록속 및 미기록종으로 보고되었다.
감사의 말
본 연구는 환경부의 재원으로 국립낙동강생물자원관의 지 원(NNIBR201902110)과 2020년 경기대학교 대학원 연구원 장학생 장학금 지원에 의하여 수행되었습니다.
References