담수 남세균, Microcystis protocystis (Chroococcales, Cyanobacteria)의 국내 미기록종에 대한 연구
이남주
1
・ 서요셉2
・ 기장서2
* ・ 이옥민1
*1경기대학교 생명과학과, 2상명대학교 생명공학과
A study of newly recorded species for freshwater cyanobacteria Microcystis protocystis (Chroococcales, Cyanobacteria) in Korea
Nam-Ju Lee
1
, Yoseph Seo2
, Jang-Seu Ki2
* , and Ok-Min Lee1
*1Department of Life Science, College of Natural Science, Kyonggi University, Suwon 16227, Republic of Korea
2Department of Biotechnology, Sangmyung University, Seoul 03016, Republic of Korea
(Received September 26, 2019; Revised January 15, 2020; Accepted January 21, 2020)
*For correspondence. (O.M. Lee) E-mail: [email protected];
Tel.: +82-31-249-9643; Fax: +82-31-241-0860 / (J.S. Ki) E-mail: [email protected];
Tel.: +82-2-2287-5449; Fax: +82-2-2287-0070
Microcystis protocystis was sampled from the Nakdong River, Geum River, and freshwaters of Yongin and Incheon. Within 3 days after sampling, morphologies of colonies and mucilage were observed using India ink. The colony of M. protocystis has irregular outlines and lacks distinct lobes and holes. Cells are distributed sparsely and irregularly within indistinct and homogeneous slime, which is diffuse at the margin. Three unispecific cultured strains from the So-ok stream were classified as Microcystis by phylogenetic analysis using 16S rRNA sequences. Three Korean isolates of M. protocystis showed 16S rRNA sequences completely identical to each other. The phylogenetic tree of the 16S rRNA sequences showed that Korean M. protocystis formed narrow clusters with other Microcystis members and M. protocystis with 99%
or greater similarities. Additional trees of RNA polymerase beta-subunit gene (rpoB) sequences separated the Korean M.
protocystis from other Microcystis. Through these results, M.
protocystis was reported as a new species in Korea.
Keywords: Microcystis protocystis, cyanobacteria, freshwater algae, newly recorded species
오늘날 남세균은 물과 육지의 다양한 서식지에 살면서 다 른 생물체에 필요한 유기화합물을 생산하여 퇴적층과 토양층 을 안정화 시키는 반면, 많은 남세균은 독성물질을 생산하거 나 녹조현상을 일으켜 전 세계적으로 심각한 문제가 되기도 한다(Graham et al., 2009).
남세균은 환경조건에 대한 뛰어난 적응성으로 다양한 환경 에 견딜 수 있으며(Whitton and Potts, 2000), 주로 담수 생태계 에 존재하고, 주요 1차 생산자로서 역할을 한다(Palacio et al., 2015). 그러나 수질의 부영양화, 지구 온난화 그리고 강우량 감소로 인해 남세균의 녹조현상이 발생되고 있다(Reichwaldt and Ghadouani, 2012). 녹조현상은 세계적으로 빈번하게 발생 되고 있으며, 남세균의 독성물질의 생산으로 인해 담수 생태 계 뿐만 아니라 인간의 건강에도 심각한 위협을 주고 있다 (Falconer, 2001).
여름철 우리나라에서도 하천이나 호수에서 남세균의 대량
발생에 의한 녹조현상이 빈번하게 발생하고 있다. 우리나라의
하천이나 호수는 질소와 인이 풍부하기 때문에 여름철 수온이
상승하고 체류시간이 길어지면 남세균의 대발생이 일어난다
(You et al., 2013). 남세균의 대발생은 심미적 또는 생태적 문
제와 더불어, 수처리 공정에서 여과지 폐쇄, 남세균의 대사물
질 생성으로 식수 공급에 여러 가지 문제점을 가져올 수 있다
(Watson et al., 2007). 국내에서도 1998년부터 물 관리 기관에
Table 1. The information about the location of five sites from where the phytoplankton samples were collected in 2018
Site Location Habitat GPS
Gangjeonggolyeong weir Gangjeongbon-gil, Dasa-eup, Dalseong-gun, Daegu Standing water 35°50'29.7"N/128°27'41.3"E So-ok stream Chuso-ri, Gunbuk-myeon, Okcheon-gun, Chungcheongbuk-do Flowing water 36°20'28.3"N/127°33'32.2"E Songwon pond Bongmu-ri, Namsa-myeon, Cheoin-gu, Yongin-si, Gyeonggi-do Standing water 37°06'26.5"N/127°08'35.4"E Idong reservoir Ebi-ri, Idong-eup, Cheoin-gu, Yongin-si, Gyeonggi-do Standing water 37°07'31.8"N/127°12'13.5"E Jangsu stream Bangsan-dong, Siheung-si, Gyeonggi-do Flowing water 37°24'38.4"N/126°45'12.3"E
서 상수원 주요 호소에 대하여 조류 발생상황을 관리함으로써 조류발생으로 인한 피해를 줄이고, 식수의 안정성을 확보하고 자 조류 경보제를 실시하고 있으며, 2013년부터 4대강 보 구간 에서도 시행하고 있다(NIER, 2013). 조류 경보제는 남세균 세 포수와 클로로필a 농도에 의해 발령되었지만, 녹조현상의 대 표성이 낮은 클로로필a 농도는 삭제하여 남세균 세포수로 단 일화 하였다(NIER, 2008).
대표적인 유해 남세균에는 Anabaena, Aphanizomenon, Mi- crocystis, Oscillatoria가 있다. 유해 4속 남세균의 분포를 살펴 보면, Anabaena 속은 세계적으로 222 분류군이 보고되어 있 으나, 국내에는 9 분류군이 보고되어 있고, Aphanizomenon 속 은 22 분류군이 보고되어 있는 반면, 국내에는 3종이 보고되어 있다(Ryu et al., 2016). 녹조현상의 주요 종인 Microcystis 속 은 68 분류군이 수록되었으나, 국내에는 9종이 보고되어 있다 (Yim et al., 2018). 또한 Oscillatoria 속은 세계적으로 501 분 류군이 보고되어 있으나, 국내에는 21 분류군이 보고되어 있 다. 이와 같이 국내 남세균에 대한 보고는 세계적인 분포에 비 해 미흡한 상황이며, 더 많은 연구를 통해 국내 남세균의 분포 를 밝혀야 한다.
녹조현상의 원인종인 Microcystis 속은 세포의 크기와 군체 의 형태적 특징으로 분류된다. 그러나 주변 환경조건이나 실 내배양 조건에 따라 많은 형태변이가 일어나기 때문에 형태적 식별형질에 의존한 동정에는 어려움이 있다(Sanchis et al., 2004). 따라서 최근에는 남세균의 분류학적 연구에 형태적인 형질과 함께 생태적 특징, 분자적인 연구방법이 도입되고 있 다(Komárek, 2016; Martins et al., 2018; Raabová et al., 2019).
국내 연구에서는 Microcystis 속의 세포벽이나 점액질속에 있는 세균을 제거함으로써 무세균으로 순수 배양하는 여러가 지 방법을 연구한 바 있다(Han et al., 2010). 또한 기존의 16S rRNA 유전자보다 rpoB 유전자가 M. aeruginosa를 명확하게 구분하는 결과를 나타냄으로써, rpoB 유전자의 분류 및 분자 검출용 마커의 유용성이 보고된 바 있다(Ki, 2010).
국외연구에서는 생태적특징과 함께 군체의 형태와 생활 사를 비교하여 M. panniformis를 신종으로 발표한 바 있다
(Komárek et al., 2002). Le Ai Nguyen 등(2012)은 베트남 열대 지방의 57 개체군의 Microcystis 속을 채집하여, 형태적 동정 과 16S rRNA 유전자 및 16S–23S ITS 염기서열의 계통학적 분 석을 비교하였다. 또한 마케도니아 Dojran 호수의 Microcystis 속 9개(M. aeruginosa, M. botrys, M. flos-aquae, M. ichthyoblabe, M. novacekii, M. protocystis, M. smithii, M. viridis, and M.
wesenbergii)의 종을 형태적 형질만으로 동정하였고, Micro- cystis 속의 형태종(morphospecies) 개념을 확립해야 한다고 보고하였다(Krstić et al., 2017).
본 연구에서는 Microcystis에 속하는 분류군을 국내 금강수 계와 낙동강 수계 및 경기도에 위치한 이동저수지로부터 채집 하였고, 일부는 단일 배양을 실시하였다. 종 동정을 위하여 생 체시료의 군체 및 세포의 형태적 형질을 관찰하였다. 또한 16S rRNA와 RNA polymerase β-subunit (rpoB) 유전자를 사용하 여 Microcystis속에 해당하는 종을 대상으로 분자계통분석을 실시하였다. 이를 통하여 Microcystis에 속하는 국내 미기록종 을 밝히는 연구를 수행하였다.
재료 및 방법
채집지 및 채집방법
본 연구의 Microcystis 속 시료는 2018년 7월에 낙동강 수계 의 강정고령보, 금강수계의 소옥천, 경기도 용인에 위치한 송 원지와 이동저수지, 그리고 인천광역시의 장수천 등의 담수역 에서 채집되었다(Table 1). 부유성 남세균의 채집은 망목의 직 경이 25 µm인 플랑크톤네트를 사용하였다(Sournia, 1978). 채 집된 시료들은 현장에서 밀봉 후, 4°C의 아이스박스에 보관하 여 실험실로 운반되었다.
실내배양
단일 배양을 위해 광학현미경 하에서 파스퇴르 피펫을 이
용하여 한 군체를 뽑아서 24-well plate (SPL)에 넣어 분리하
였다. 24-well plate에서 1~2주간 배양 후, 오염여부를 확인하
고, 단일 배양된 배양주를 50 ml cell culture flask (SPL)에 옮겨 대량배양을 실시하였다. 단일 배양 및 대량배양은 CB 배지 (Shirai et al., 1989)와 BG-11 배지(Stanier et al., 1971)를 사용 하였다. 실내배양은 20~25°C, 명암 주기 16 h : 8 h, 조도는 25 μmol/m 2 /sec의 조건 하에서 실시되었다. 단일 배양된 배양체 3 주는 국립낙동강생물자원관에 기탁하였다(FBCC-A403, FBCC- A404, FBCC-A405).
형태관찰
본 연구에서는 채집된 분류군의 생체시료를 3일 이내에 광 학현미경(Axio Imager A2, Carl Zeiss ; Olympus BX53, Olympus) 을 이용하여 100~1000 배율로 관찰하였고, 200~400배 하에 서 사진촬영을 실시하였다(AxioCam HRC camera, Carl Zeiss;
an Olympus UC-90, Olympus). 사진 촬영 시 군체의 점액질 형 태 관찰을 위해서 먹물(India ink)을 사용하였다.
남세균의 분류체계는 Komárek 등(2014)에 따랐고, Algae- base를 참조하였다(Guiry and Guiry, 2019). 또한 남세균의 동 정에는 Crow (1923), Komárek와 Anagnostidis (1998), Komárek 등(2002), Palacio 등(2015) 그리고 Krstić 등(2017)을 참조하 였다.
DNA 추출
남세균의 genomic DNA (gDNA) 추출을 위해 3개의 배양 주 세포(50 ml)를 3,000 rpm에서 10분 동안 원심 분리하여 수확 하였다. 농축된 세포를 200 µl 1 × TE buffer (10 mM Tris-HCl;
pH 8.0, 1 mM EDTA)에 희석하여 -20°C에서 DNA를 추출하 기 전까지 보관하였으며, 이후 cetyltrimethylammonium bromide (CTAB) 방법을 이용하여 gDNA를 추출하였다(Richards et al., 2003).
효소중합연쇄반응(PCR) 및 DNA 시퀀싱
남세균 Microcystis 배양주 3주의 16S rRNA 유전자를 박테 리아 16S 대상의 범용 프라이머(27F, 5'-AGA GTT TGA TCC TGG CTC AG-3'; 1492R, 5'-GGT TAC CTT GTT ACG ACT T-3')를 사용하여 PCR 기법으로 증폭하였다. 추가적으로 rpoB 유전자를 기존에 보고된 프라이머(MarpoBF1026, 5'- CAT TCG GGA ACG GAT GAC C-3'; Ma-rpoBF2013, 5'- CTT CGT AGT TAT AGC CTT CC-3')를 이용하여 분석하였 다(Ki, 2010). PCR 반응은 추출한 gDNA (1 µl), 각각의 프라이 머(10 pmole, 1 µl)와 PCR 반응액(17 µl)을 혼합하여 실시하였 다. PCR 반응액으로 1× PCR buffer, 0.2 mM dNTPs, 0.5 mM
forward-reverse 프라이머, 1 unit Ex Taq polymerase (Takara Bio)를 사용하였다. PCR 반응은 iCycler TM (BioRad)를 이용 하여 초기 94°C 5분간 DNA를 변성 시키고, 이후 94°C 20초, 50°C 30초, 72°C 60초를 40회 반복하여 대상 유전자 영역을 증폭하였다. PCR 증폭 반응이 종료되면 추가로 72°C 10분간 유지하여 반응을 종결하였다. 증폭된 PCR 산물은 1% 아가로 스 겔에서 전기영동하여 관찰하였다.
DNA sequencing은 PCR 산물을 QIAquick PCR purification kit (Qiagen GmbH)로 정제하여, PCR 프라이머(27F, CY-23R 600, MarpoBF1026, Ma-rpoBF2013, 시퀀스-walking 프라이 머)와 ABI PRISM® BigDye TM Terminator Cycle Sequencing Ready Reaction kit (PE Biosystems)를 이용하여 실시하였다.
DNA sequencing 단편은 자동 DNA 분석기(Model 3700, Applied Biosystems)로 분석하였다. 각각의 배양주로부터 얻은 염기 서열 단편을 Sequencher 4.1.4 (Gene Codes, MI)을 이용하여 조합된 단일 염기서열로 만들고, 염기서열을 GenBank에 등 록하였다(Table 2).
염기서열 유사도, 유전거리, 계통 분석
남세균 Microcystis 배양주 3주의 16S rRNA 유전자의 염기 서열 유사도(DNA similarity)와 유전거리(genetic distance)를 분석하였다. 본 연구에서 규명한 염기서열과 GenBank로부터 얻은 Microcystis 염기서열로 데이터 조합을 만들고, MAFFT 소프트웨어(Katoh and Standley, 2013)를 이용하여 염기서열 을 재배열(alignment)한 후, 같은 길이가 되도록 양 끝을 제거 하였다. 이후, BioEdit를 이용하여 유사도(similarity)를 계산 하고, MEGA 6.0 (http://www.megasoftware.net)에서 Kimura 2-parameter 모델을 이용하여 유전거리를 계산하였다.
본 연구에서 규명한 국내 Microcystis protocystis와 Gen-
Bank로부터 얻은 14주, 총 15주의 Microcystis 속의 16S rRNA
염기서열을 이용하여(Table 2), Maximum Likelihood (ML) 계
통분석을 실시하였다. 분석은 16S rRNA 염기서열을 MAFFT
소프트웨어(Katoh and Standley, 2013)로 재배열하고, 이후 양
끝을 동일한 크기로 자른 후 데이터 세트를 준비하였다(16S
rRNA, 1,487 sites에서 1,437 sites 선택). ML 계통도는 General
Time Reversible (GTR)을 이용하여 RAxML ver. 8.0에서 추
정하였다(Stamatakis, 2014). 추가로 동일한 모델과 염기서
열 데이터를 이용하여 MrBayes ver. 3.1.2 (Huelsenbeck and
Ronquist, 2001)로 Bayesian tree를 추정하였다. 같은 방법으
로 rpoB 유전자(885 sites에서 882 sites 선택)의 분자계통분석
을 실시하였다.
Table 2. List of species, genes, and their GenBank accession numbers used in this study
Species Strains Isolation locality Accession No.
16S rRNA rpoB
Microcystis protocystis FBCC-A403 Korea MN472745
#MN919197
#Microcystis protocystis FBCC-A404 Korea MN472746
#Microcystis protocystis FBCC-A405 Korea MN472747
#Microcystis aeruginosa NIES-298 Japan FJ461749 HM991461
Microcystis bengalensis VN486 Viet Nam AB666082
Microcystis botrys KLL-C017 Israel KP726251
Microcystis flos-aquae NRERC-218 Korea MK598764
Microcystis ichthyoblabe NRERC-217 Korea MK972443
Microcystis novacekii BC18 Japan AB035551
Microcystis panniformis VN425 Viet Nam AB666076
Microcystis panniformis FACHB-1757 China CP011339
Microcystis protocystis VN111 Viet Nam AB666054
Microcystis pseudofilamentosa VN511 Viet Nam AB666084
Microcystis ramosa VN493 Viet Nam AB666083
Microcystis robusta VN432 Viet Nam AB666078
Microcystis smithii CHAB2183 China GQ496076
Microcystis viridis TAC17 Japan AB012328
Microcystis viridis NIES-102 Japan AP019314
Microcystis wesenbergii NRERC-212 Korea MK972442
Microcystis wesenbergii NIES-107 Japan HM991460
#
Sequences determined in this study.
결과 및 고찰
본 연구에서는 2018년 7월에 낙동강 수계의 강정고령보, 금 강수계의 소옥천, 경기도 용인에 위치한 송원지와 이동저수 지, 그리고 인천광역시의 장수천 등의 담수역에서 Microcystis protocystis를 채집하였다. 종 동정은 군체 및 세포의 형태적 특 징을 사용하였으며, 이를 통하여 M. protocystis로 동정 되었 다. 또한 단일 배양된 금강수계 소옥천의 배양주는 16S rRNA 염기서열에 의한 계통분석을 통해 Microcystis 속의 clade에 포함 되었고, rpoB 유전자를 이용한 계통분석에서 각각의 종 이 뚜렷하게 분리되었다.
본 연구의 M. protocystis는 국내 미기록종으로 추가되었으 며, 따라서 보고된 국내 Microcystis 속은 총 10종이다.
형태적 특징
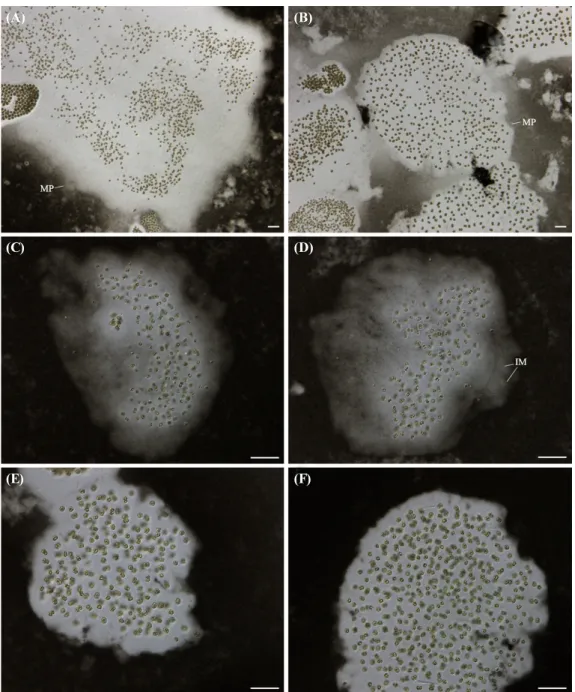
Family Microcystaceae Elenkin 1933 Genus Microcystis Lemmermann 1907 Microcystis protocystis Crow 1923 (Fig. 1)
Synonyms: Microcystis aeruginosa f. protocystis (Crow)
Elenkin, 1938
군체는 부유성이며, 매우 미세하여 육안으로 잘 보이지 않 는다. 군체의 윤곽은 불규칙하며, 구획화 되어 있지 않고 연결 되어 있으며, 점액질 내에 빈 공간이 없다. 무색의 점액질은 현 미경 상에서 잘 구분되지 않고, 점액질의 경계는 뚜렷하지 않 아서 먹물을 넣어서 경계를 확인할 수 있다. 세포는 구형이며, 분열 전에 약간 길어진다. 무색의 불분명한 점액질 내에서 세 포는 밀집해 있지 않고, 불규칙하게 분산되어 있으며, 간혹 각 각의 세포에 점액질이 약간 발달한다. 세포의 직경은 (3.4)4- 5.6(7.6) μm이다.
Ecology: 담수에 분포하는 종이다(Palacio et al., 2015; Krstić et al., 2017).
Distribution: 스리랑카(Komárek and Anagnostidis, 1998), 브 라질(Werner, 2010), 오스트레일리아(Bostock and Holland, 2010), 인도(Gupta, 2012)
Site of collection: 충청북도 옥천군 군북면 추소리 636-1
(36°20'28.3"N/127°33'32.2"E)
(A) (B)
(C) (D)
(F) (E)
Fig. 1. Microscopic photographs of Microcystis protocystis (MP) collected from Korea freshwaters (mucilage stained by India ink). (A) Gangjeonggolyeong- bo, (B) Sookcheon, (C–D) Idong reservoir, (E–F) Jangsucheon. (A–B) Distributed sparsely and irregularly cell, (C) hyaline and diffluent mucilage, (D) individual mucilaginous envelopes (IM), (E–F) irregular outline and without lobes. Scale bars represent 20 μm.
Date of collection: 2018년 07월 13일
Specimen Locality: FBCC-A403, FBCC-A404, FBCC-A405
염기서열 특성 및 분자계통
본 연구에서 남세균 Microcystis protocystis의 배양주 3개 의 16S rRNA 염기서열을 규명하였으며(Table 2), 100% 동 일한 서열로 파악되었다. 또한, NCBI (National Center for Biotechnology Information) 데이터베이스에서 BLAST 검색
을 통해 Microcystis와 99% 이상의 유사도를 갖는 것으로 파악 되었다.
NCBI에 등록된 남세균 Microcystis 15종과 국내 배양주의 16S rRNA와 rpoB 유전자 염기서열을 이용하여 분자계통분 석을 실시하였다(Fig. 2). 16S rRNA ML tree에서 국내 M.
protocystis와 다른 Microcystis 종(M. aeruginosa, M. robusta,
M. smithii 등)이 같은 cluster를 형성하여, 각각의 종을 뚜렷하
게 구분할 수 없었다. 16S rRNA Bayesian tree 분석에서 유사
(A) (B) (C)
Fig. 2. Phylogenetic relationships of cyanobacteria genus Microcystis, including a Korean M. protocystis. (A) Maximum Likelihood (ML) tree of Microcystis 16S rRNA, (B) Bayesian tree of Microcystis 16S rRNA, (C) ML tree of Microcystis rpoB, (D) Bayesian tree of Microcystis rpoB. Each 16S rRNA or rpoB sequence of Cyanothece sp. (Oscillatoriales) was included as the separate outgroup. Additional Neighbor-Joining (NJ) tree generated similar topology of the ML tree, and thus its bootstrap proportions (BP) were incorporated into the tree. The numbers at the nodes display bootstrap proportions (> 50%) or posterior probabilities (0.5) obtained in the ML and Bayesian analyses, respectively. Branch lengths are proportional to the scale given. Bold represents data obtained in this study. Bold represents a representative Korean isolate, which sequence was completely identical to those of FBCC-A404 and FBCC-A405.
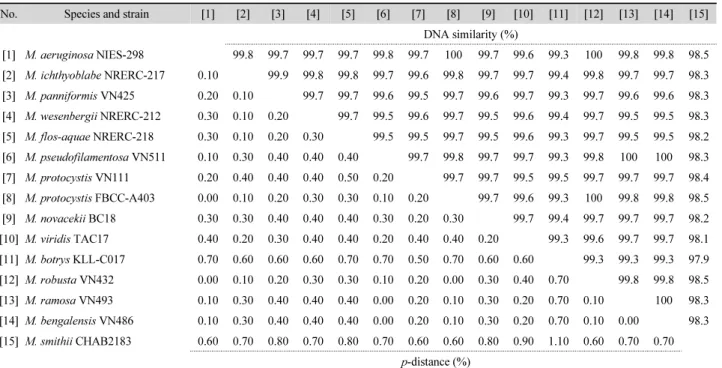
Table 3. Similarity scores (above diagonal) and % genetic p-distance (below diagonal) estimated by the Kimura 2-parameter model between 15 pairs of the aligned sequence data of the nearly complete 16S rRNA gene from Microcystis
No. Species and strain [1] [2] [3] [4] [5] [6] [7] [8] [9] [10] [11] [12] [13] [14] [15]
DNA similarity (%)
[1] M. aeruginosa NIES-298 99.8 99.7 99.7 99.7 99.8 99.7 100 99.7 99.6 99.3 100 99.8 99.8 98.5 [2] M. ichthyoblabe NRERC-217 0.10 99.9 99.8 99.8 99.7 99.6 99.8 99.7 99.7 99.4 99.8 99.7 99.7 98.3 [3] M. panniformis VN425 0.20 0.10 99.7 99.7 99.6 99.5 99.7 99.6 99.7 99.3 99.7 99.6 99.6 98.3 [4] M. wesenbergii NRERC-212 0.30 0.10 0.20 99.7 99.5 99.6 99.7 99.5 99.6 99.4 99.7 99.5 99.5 98.3 [5] M. flos-aquae NRERC-218 0.30 0.10 0.20 0.30 99.5 99.5 99.7 99.5 99.6 99.3 99.7 99.5 99.5 98.2 [6] M. pseudofilamentosa VN511 0.10 0.30 0.40 0.40 0.40 99.7 99.8 99.7 99.7 99.3 99.8 100 100 98.3 [7] M. protocystis VN111 0.20 0.40 0.40 0.40 0.50 0.20 99.7 99.7 99.5 99.5 99.7 99.7 99.7 98.4 [8] M. protocystis FBCC-A403 0.00 0.10 0.20 0.30 0.30 0.10 0.20 99.7 99.6 99.3 100 99.8 99.8 98.5 [9] M. novacekii BC18 0.30 0.30 0.40 0.40 0.40 0.30 0.20 0.30 99.7 99.4 99.7 99.7 99.7 98.2 [10] M. viridis TAC17 0.40 0.20 0.30 0.40 0.40 0.20 0.40 0.40 0.20 99.3 99.6 99.7 99.7 98.1 [11] M. botrys KLL-C017 0.70 0.60 0.60 0.60 0.70 0.70 0.50 0.70 0.60 0.60 99.3 99.3 99.3 97.9 [12] M. robusta VN432 0.00 0.10 0.20 0.30 0.30 0.10 0.20 0.00 0.30 0.40 0.70 99.8 99.8 98.5 [13] M. ramosa VN493 0.10 0.30 0.40 0.40 0.40 0.00 0.20 0.10 0.30 0.20 0.70 0.10 100 98.3 [14] M. bengalensis VN486 0.10 0.30 0.40 0.40 0.40 0.00 0.20 0.10 0.30 0.20 0.70 0.10 0.00 98.3 [15] M. smithii CHAB2183 0.60 0.70 0.80 0.70 0.80 0.70 0.60 0.60 0.80 0.90 1.10 0.60 0.70 0.70
p-distance (%)
한 branch 양상을 보였으며, 특히 M. pseudofilamentosa, M.
bengalensis, M. ramosa를 제외하고 모든 종이 polytomic 패턴 을 보였다. 이것은 이들 사이의 염기서열 유사도 높고(> 99%)
유전적 거리가 낮기(< 0.5%) 때문이다(Table 3). 반면, 한국 M.
protocystis와 다른 4종의 Microcystis의 rpoB 유전자를 이용
한 계통분석에서 각각의 종이 뚜렷하게 분리되었다. 남세균과
Microcystis의 16S rRNA와 RNA polymerase beta subunit (rpoB) 유전자를 이용한 선행연구에서도(Ki, 2010; Cheon et al., 2011), 16S rRNA의 해상도가 낮아 종을 구분하지 못하는 것으로 알려졌지만, rpoB 유전자 마커는 종을 뚜렷하게 구분 하는 것으로 파악되었다. 현재, 유전자은행에 등록된 Micro- cystis의 rpoB 유전자 염기서열은 매우 제한적이다. 향후, Micro- cystis 종을 분자적으로 명확히 구분하기위해, 추가 종의 rpoB 유전자 염기서열 발굴 및 다른 유전자 마커를 이용한 분석이 필요하다.
국내 M. protocystis 16S rRNA는 베트남에서 보고된 종의 염기서열(AB666054)과 99.7% 유사도와 0.205% 유전거리를 갖는 것으로 파악되었다. 또한, 한국 M. protocystis rpoB 유전 자는 근연종인 M. aeruginosa NIES-298와 98.2% 유사도와 1.721% 유전거리를 보였다. 본 연구에서 실시한 분자계통분 석, rpoB 유전자 염기서열 유사도 및 유전거리 결과는 우리나 라 Microcystis 배양주가 M. protocystis라는 것을 분자적으로 제시하였다.
적 요
국내 낙동강 수계, 금강수계, 경기도 용인 그리고 인천광역 시의 담수역으로부터 Microcystis protocystis를 채집하였다.
채집 후 3일 이내의 시료에 먹물을 사용하여 군체 및 점액질의 형태를 관찰하였다. M. protocystis는 군체의 점액질 초 윤곽의 경계가 불분명하고, 세포가 점액질 속에서 분산되어 있는 특 징을 나타냈다. 단일 배양된 금강수계 소옥천의 3주의 배양주 는 모두 동일한 16S rRNA 염기서열을 나타냈고, 계통수 분석 을 통해 Microcystis 속으로 분류되었다. 분자계통분석에서 국내 배양주는 기존에 보고된 M. protocystis를 포함한 다른 Microcystis 종과 99% 이상의 유사도로 같은 클러스터를 형성 하였다. 추가적인 rpoB 유전자를 이용한 분석은 한국의 M.
protocystis를 다른 종과 분리하였다. 이와 같은 결과를 통하여 M. protocystis를 국내 미기록종으로 보고하였다.
감사의 말
본 논문은 환경부의 재원으로 국립낙동강생물자원관의 지 원을 받아 수행하였습니다(NNIBR201902110).
References