J. of Korean Orthopaedic Research Society V o l u m e 9 , N u m b e r 1 , A p r i l , 2 0 0 6
※ 통신저자: 문 형 태
서울특별시 중구 저동 2가 85번지 인제대학교 서울백병원 정형외과학교실
TEL: 02) 2270-0028 FAX: 02) 2270-0023 E-mail: [email protected]
배양 연골세포의 증식과 분화에 대한 히알우로난의 효과
인제대학교 의과대학 서울백병원 정형외과학교실 문형태∙고한석
= Abstract =
Effects of Hyaluronan on Proliferation and Differentiation Cultured Chondrocyte
Hyung Tae Moon, M.D., Han Suk Ko, M.D., Ph.D.
Department of Orthopedic Surgery, Seoul Paik Hospital, Inje University
Objectives: Recent basic science studies continue to further our understanding of the fundamental mecha- nisms that likely underlie the therapeutic benefits of hyaluronan derivatives. The purpose of this study is to elu- cidate the effects of hyaluronan on ATDC5 proliferation and differentiation.
Methods: ATDC5 cells derived from mouse teratocarcinoma have the capacity to differentiate along a num- ber of connective tissue pathways and are an attractive source of chondrocyte precursor cells. In this study, hyaluronan influencing ATDC5 chondrogenesis were investigated using an bone block culture system. The cell proliferation was analyzed by MTT assay. To validate ATDC5 differentiation we studied ALP activity, colla- gen content and western blot of Hsp40.
Results: In cell proliferation, ATDC5 cells didn’t show significant difference between controls and hyaluro- nan-treated cultures. But hyaluronan induced ALP activity and increased collagen accumulation. Hyaluronan- treated ATDC5 cells expressed Hsp40 mRNA and protein within 24 hours.
Conclusions: Hyaluronan-induced chondrogenic differentiation was not associated with ATDC5 cell prolif- eration. Hyaluronan-induced Hsp40 in cells can protect the cell function from damaged protein. These data pro- vide new insights into regulatory mechanism defining pharmacological effects of hyaluronan.
Key Words: Hyaluronan, ATDC5 cell, Chondrocyte differentiation, Bone block culture, Hsp40
Introduction
Hyaluronan (HA) is a critical con- stituent component of normal synovial fluid and an important contributor to joint homeostasis20).
Viscosupplements are HA-based prepara- tions that have been developed to aug- ment the diminished biologic and mechan- ical properties of osteoarthritic synovial fluid. There are at least four mechanisms whereby intra-articular injection of HA derivatives may provide therapeutic bene- fit in symptomatic knee OA23). The benefi- cial effects of viscosupplementation on osteoarthritic synovial fluid may include restoration of (1) elastic and viscous prop- erties, (2) anti-inflammatory effects, (3) antinociceptive effects, and (4) normaliza- tion of HA synthesis by synoviocytes.
In clinical trials and in practice, visco- supplements have effectively modified osteoarthritic pain and functional disabili- ty. Clinical trials of 6~12 months dura- tion have demonstrated significantly greater pain relief for both low molecular weight HA and high molecular weight hylan preparations compared with saline control injections32,25,8,2). Multicenter, prospective, comparative trials have demonstrated that viscosupplementation is at least as effective as NSAIDs. Viscosup- plementation using hylan G-F 20(Synvisc) was well tolerated and had effectiveness that was at least comparable to that of continuous oral NSAID therapy in the management of knee pain in a 6-month study1). A low molecular weight HA preparation (Hyalgan) showed an effect similar to that of naproxen on OA pain2).
In osteoarthritic synovial fluid, the mol- ecular weight and concentration of HA
are decreased due to dilutional effects, aberrant HA synthesis, and free radical degradation. Consequently, the biologic and mechanical properties normally pro- vided to synovial fluid by HA are compro- mised in osteoarthritic synovial fluid.
Accumulating in vitro and in vivo evi- dence from animal models suggests that exogenous HA may have beneficial effects on chondrocyte metabolism that can potentially facilitate maintenance and repair of the cartilage matrix. Identified anabolic effects include enhanced synthe- sis and remodeling of matrix macromole- cules and chondrocyte proliferation.
The effects of HA on the cellular events are poorly understood. The purpose of this study is to elucidate the effects of HA on ATDC5 differentiation.
Materials and Methods 1. Materials
HA preparations were extracted and purified from rooster comb tissue and had a molecular weight of 5-7×105 Da (Hyru- an®: LG Life Sciences, Korea). Two con- centrations were evaluated for the effect on the viability of normal chondrocytes in primary culture: 0.4 and 50 μg/ml. The more effective one of these doses was then chosen for the subsequent study.
2. Cells and cell cultures
A chondrogenic mouse embryonic carci- noma cell line, ATDC5, was obtained from RIKEN cell bank (Tsukuba, Japan).
Cells were maintained in a 1:1 mixture of Dulbecco’s modified Eagle’s medium and Ham’s F12 (D-MEM/F-12, Sigma) supple-
mented with 5% fetal bovine serum (Intergen, Purchase, NY) and penicillin (50 IU/ml) streptomycin (50 μg/ml) (ICN Biomedicals, Aurora, OH). In this study, ATDC5 chondrogenesis were investigated using bone block in a three-dimensional format culture system.
Cells were maintained at 37�C in a humidified 5% CO2/95% air atmosphere.
Cells were plated at 3.5×105 cells/bone block in 24-well plates. After cells attached to the plate, the medium was replaced with fresh medium containing agents of various concentrations or vehi- cle, and the culture was continued for 24 days with medium change three times per week.
3. Biochemical determination
1) MTT assay
Viability rates were determined using the MTT-based (Thiazolyl blue) cytotoxici- ty test. This test is based on mitochondria viability: only functional mitochondria can oxidize an MTT solution, giving a typical blue-violet end product. Cells were plated at 5,000 cells/well in 24-well plates. After cells attached to the plate, the medium was replaced with fresh medium contain- ing agents of various concentrations or vehicle, and the culture was continued for 7 days with medium change three times.
After addition of MTT solution (50 μ l/well, 5 mg/ml in PBS, Sigma), the plates were incubated for 4 h. The medi- um was removed and the blue dye formed was dissolved in 300 μl of 0.04 N HCl in isopropanol. Absorbance was measured at 580 nm.
2) Total protein content
The protein content of the cell culture was determined using a Bradford method4). It was estimated with the pro- tein assay kit (Bio-Rad) in the cell lysates. Reaction product of samples and dye reagent were read on a microplate reader at 595 nm. The total protein con- tent was normalized by linear range of bovine serum albumin (BSA) standard.
3) Alkaline Phophatase (ALP) activity ALP activity was determined with Sigma diagnostic kit (104-LL). To mea- sure ALP activity quantitatively, the supernatant of cell lysates was assayed by incubating with reaction solution (0.6 M 2-amino-2-methyl-l-propanol (pH 10.0) and 10 mM p-nitrophenyl phosphate) at 37�C for 15 min. The reaction was stopped by adding 0.05 N NaOH. The absorbance was measured at 405 nm and converted to p-nitrophenol (Sigma) concentration based on standard solutions prepared in parallel.
4) Collagen assay
Total monolayer collagen content was determined as the total soluble collagen by using the Sircol Collagen Assay kit (Bio- color, Belfast, Northern Ireland). Cells were hydrolyzed in .5 M acetic acid at 4�C for 17-18 h. Aliquots of the cell lysates were added to 1 ml of Sircol dye reagent that binds to collagen, followed by 30 min of mixing. After centrifugation at 10,000g for 10 min, the supernatant was carefully decanted, and 1 ml of 0.5 M NaOH was added. Samples and collagen standards were then read at 540 nm on a spec- trophotometer. Collagen concentration in the lysate buffer was then read from a standard curve with the use of acid-solu-
ble type 1 collagen. Collagens contain 14%
hydroxyproline, and collagen concentration obtained using Sirus Red binding corre- lates well with the hydroxyproline content (Sircol Collagen Assay, Biocolor).
4) Western blot analysis
ATDC5 cells were incubated in α MEM/0.5% FBS for 24 hours before treat- ment with HA for 24 hours. The protein concentration in the cell lysate was mea- sured using a Protein Assay Kit II (Bio- Rad). Equivalent amounts of cell lysates were electrophoresed by 10% SDS-PAGE22) and transferred to nitrocellulose mem- brane. After blocking with 5% bovine serum albumin, the membrane was incu- bated with polyclonal goat antibody against Hsp40 (sc-1801, Santa Cruz Biotechnology) and with peroxidase-conju- gated anti-goat IgG antibody. Hsp40 was visualized using the ECL advanced west- ern blotting detection kit (Amersham, Arlington Heights, IL) following the man- ufacturer’s instructions. Reactive polypep- tides were detected by exposure to Hyper-
filmTM ECL (Amersham, Arlington Heights, IL).
5) Statistic al analysis
Results are presented as mean±S.D.
The analysis was conducted with SPSS software (SPSS Inc., Chicago, IL, U.S.A.). Statistical comparison among the groups was performed with one way analysis of variance (ANOVA) followed by Dunnett t-test. A probability below 5%
was considered statistically significant.
Results
1. Effects of HA on ATDC5 cell prolifer- ation and protein synthesis
ATDC5 cells were induced to chondro- genic differentiation through a cellular condensation process, resulting in the for- mation of cartilage nodule like cell aggre- gates. ATDC5 cells grew until confluence on day 5, as indicated by MTT assay (Fig. 1). In cell proliferation, they didn’t show significant difference between con-
Fig. 1. Effect of HA (0.4 μg/ml) on proliferation of ATDC5 cells in medium containing 5% FBS.
Cells were plated at a density of 5,000 cells/well and medium replaced every 2-3 day. The growth was estimated by MTT assay. Data expressed as mean±S.D. for five cultures.
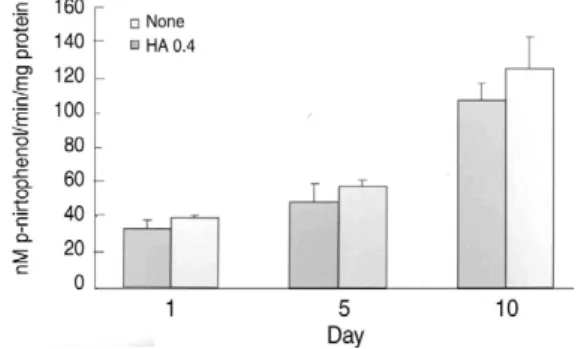
Fig. 2. Effect of HA on the ALP activity in ATDC5 cells. The cells were plated at a density of 3.5×
105cells/bone block in 24-well plates and treated with HA. The ALP activity was determined using the cell lysates. The data are expressed as nmol substrate cleaved/min/mgprotein.
trols and HA-treated cultures at all time- points.
2. Effects of HA on chondrogenic matura- tion
In control cultures, ATDC5 cells began to undergo differentiation and nodular condensation from day 7. By day 10, a significant increase in ALP activity was seen (p<0.05) (Fig. 2). Treatment with HA resulted in slightly increased ALP staining and induction of ALP activity in HA-treated cultures at day 10 compared with controls (Fig. 2). Furthermore, there was increased collagen accumulation in HA-treated cultures at day 20 (Fig. 3).
These data indicate that HA-induced chondrogenic differentiation was not asso- ciated with proliferation of ATDC5 cells (Fig. 1).
3. Effects of HA on Hsp40 expression
HA actions, in part, are mediated by expression of some genes. Among these genes, Hsp40 expression was increased with HA treatment (Fig. 4). Immunoblot
analysis shows the alteration of Hsp40 expression with the HA-treatment (Fig.
5). This result suggests that HA induces Hsp40 in ATDC5 cells.
Discussion
HA is a polymer of molecular mass from 105 to 106 comprising repeating disaccha- ride units of glucuronic acid and N-acetyl- D-glucosamine. HA is abundant, present in virtually all biologic fluids. Moreover, there is a brisk metabolism of HA in humans, with approximately one-third of total HA degraded and replaced per day predominantly by the reticuloendothelial system9). HA performs three functions: It expands the extracellular space by binding salt and water, interacts with a variety of extracellular molecules to form a compos- ite extracellular matrix, and is recognized by several cell surface receptors that acti- vate intracellular signaling pathways in response to HA (and likely, costimulatory signals) or function in HA internaliza- tion30). HA is especially prominent during embryonic development and at sites of wound healing.
In a partial menisectomy rabbit model of OA, total proteoglycan synthesis by chon- drocytes was significantly higher in the HA group compared with the control group14). Proteoglycan synthesis was also stimulated by exogenous HA in equine cultured chondrocytes and cartilage explants10). HA was capable of fully restoring proteoglycan content to both osteoarthritic human cartilage and carti- lage that had been previously damaged by fibronectin18). A recent study in injured rabbit menisci found that intra-articular HA administered in five weekly injections Fig. 3. Effect of HA on the collagen of ATDC5 cells.
The cells were plated at a density of 3.5×105 cells/bone block in 24-well plates and treated with HA.
increased the degree of collagen remodel- ing and reduced swelling when compared with saline vehicle28). Importantly, HA has been found to have a dose-dependent, pro- liferative effect on cultured chondrocytes, as well as enhancing chondroitin sulfate synthesis19).
In the course of chondrogenesis there are at least two steps regulated by local and systemic factors. The first is mes- enchymal condensation, which gives rise to proliferating chondrocytes, and the sec- ond comprises differentiation of proliferat- ing chondrocytes to hypertrophic cells fol- lowed by mineralization of cartilage matrix13). In medium containing stripped serum, the presence of insulin promoted condensation of ATDC5 cells during a rapid growth phase, followed by nodule formation during a much slower growth phase. After proliferation ceased, the cells
differentiated to the hypertrophic stage as indicated by increased ALP activity and secretion of a matrix that is competent for mineralization. These features repro- duced those described in the initial char- acterization of ATDC5 cell chondrogene-
sis3,26). In this study, HA induces chondro-
genic differentiation but not affects pro- liferation of ATDC5 cells
It is now widely accepted that most Hsps have a molecular chaperone activity involved in various aspects of the life cycle of the proteins. A molecular chaper- one is a protein that binds to and stabi- lizes an otherwise unstable conformer of another protein and, by controlled binding and release of the substrate protein, facil- itates its correct fate in vivo: be it fold- ing, oligomeric assembly, transport to a particular subcellular compartment, or controlled switching between active/inac-
Fig. 4. Induction of Hsp40 mRNA with hyaluronan.
tive conformations16). A recent under- standing of the general functions of mole- cular chaperones has emerged from research on the Hsp70s and chaperonins Hsp60s. Both Hsp70 and Hsp60 have a weak ATPase activity15).
In mammals, one of the activities asso- ciated with Hsp70 is the disassembly of clathrin cages into triskelions. This disas- sembly process is mediated by a `clathrin- uncoating ATPase’, which proved to be identical to the constitutive form of Hsp707). Recently, one of the DnaJ homologues, auxilin, has been shown to be involved in this disassembly process31). Cytosolic Hsp70 and Hsp40 are known to bind to the nascent polypeptide chains emerging from translating ribosomes and facilitate their correct folding12,11)Also, as demonstrated in an in vitro system, the binding of Hsp70 to a substrate prone to thermal aggregation was found to be dependent on Hsp40-enhanced ATP hydrolysis. Together with Hsp40, Hsp70 can protect luciferase from heat-induced aggregation and promote the renaturation of thermally denatured luciferase24).
The beneficial functions of molecular chaperones are observed not only at the cellular level but also at the tissue level and for the whole body. For example, induction of molecular chaperones in ani- mals by whole body hyperthermia can protect the brain and heart from tissue injury induced by ischemia21,5,6). Also, mod- erate overexpression of molecular chaper- ones resulted in an extended life span in nematoda and the fruit fly29). It is believed that the accumulation of dam- aged protein induced by oxidative stress is one of the major causal factors of senes- cence17,27). In these studies we demonstrat- ed for the first time that HA-treated ATDC5 cells express Hsp40.
Collectively, the results of these studies suggest that viscosupplementation with HA preparations may have the potential to mitigate the development of OA or pre- vent its progression. Despite differing study designs, OA models, endpoints, parameters, and HA preparations, evi- dence suggesting that intra-articular HA therapy can have a beneficial effect on chondrocyte metabolism and cartilage matrix integrity is increasing. This is an important area for further study.
Conclusions
In cell proliferation, ATDC5 cells didn’t show significant difference between con- trols and HA-treated cultures. But HA induced ALP activity and increased colla- gen accumulation. These data indicate that HA-induced chondrogenic differentia- tion was not associated with ATDC5 cell proliferation.
HA-treated ATDC5 cells expressed Hsp40 mRNA and protein. Induction of Hsp40 in Fig. 5. Immunoblot analysis of Hsp40 in ATDC5 cells.
Extracts of cell culture on day 1 were subjected to 10% SDS-polyacrylamide gel electrophoresis.
E. coli expressed Hsp40(lane1), none-induced ATDC5 cell (lane2) and HA-induced ATDC5 cell (lane3).
cells can protect the cell function from damaged protein. These data provide new insights into regulatory mechanism defin- ing pharmacological effects of HA.
REFERENCES
11) Adams ME, Atkinson MH, Lussier AJ, et al.:
The role of viscosupplementation with hylan G-F 20 (Synvisc) in the treatment of osteoarthritis of the knee: a Canadian multicenter trial comparing hylan G-F 20 alone, hylan G-F 20 with non- steroidal anti-inflammatory drugs (NSAIDs) and NSAIDs alone. Osteoarthritis Cartilage. 3:213-225, 1995.
12) Altman RD, Moskowitz R, Hyalgan Study Group. Intraarticular sodium hyaluronate (Hyal- gan) in the treatment of patients with osteoarthri- tis of the knee: a randomized clinical trial. J Rheumatol. 25:2203-2212, 1998.
13) Ballock RT, Reddi AH: Thyroxine is the serum factor that regulates morphogenesis of columnar cartilage from isolated chondrocytes in chemical- ly defined medium. J Cell Biol. 126(5):1311-8, 1994.
14) Bradford MM: A rapid and sensitive method for the quantitation of microgram quantities of pro- tein utilizing the principle of protein-dye bind- ing. Anal Biochem. 72:248-54, 1976.
15) Brown IR, Sharp FR.: The cellular stress gene response in brain. In: Latchman DS, editor.
Stress proteins. Berlin: Springer-Verlag. p. 243- 263, 1999.
16) Carroll R, Yellon DM: Heat stress proteins and their relationship to myocardial protection. In:
Latchman DS, editor. Stress proteins. Berlin:
Springer-Verlag. p. 265-279, 1999.
17) Chappell TG, Welch WJ, Schlossman DM, Palter KB, Schlesinger MJ, Rothman JE.:
Uncoating ATPase is a member of the 70 kilo- dalton family of stress proteins. Cell. 45(1):3-13, 1986.
18) Dougados M, Nguyen M, Listrat V, Amor B.:
High molecular weight sodium hyaluronate (hyalectin) in osteoarthritis of the knee: a 1 year placebo-controlled trial. Osteoarthritis Cartilage.
1:97-103, 1993.
19) Fraser JR, Laurent TC, Laurent UB: Hyaluro- nan: its nature, distribution, functions and turnover. J Intern Med. 242(1):27-33, 1997.
10) Frean SP, Abraham LA, Lees P: In vitro stimu- lation of equine articular cartilage proteoglycan synthesis by hyaluronan and carprofen. Res Vet Sci . 67:183-190, 1999.
11) Frydman J, Hartl FU: Principles of chaperone- assisted protein folding: differences between in vitro and in vivo mechanisms. Science. 272(5267):1497- 502, 1996.
12) Frydman J, Nimmesgern E, Ohtsuka K, Hartl FU: Folding of nascent polypeptide chains in a high molecular mass assembly with molecular chaperones. Nature. 370(6485):111-7, 1994.
13) Hall BK, Miyake T: Divide, accumulate, differ- entiate: Cell condensation in skeletal develop- ment revisited. Int J Dev Biol. 39:881-893, 1995.
14) Han F, Ishiguro N, Ito T, Sakai T, Iwata H:
Effects of sodium hyaluronate on experimental osteoarthritis in rabbit knee joints. Nagoya J Med Sci . 62:115-126, 1999.
15) Hartl FU: Molecular chaperones in cellular pro- tein folding. Nature. 381(6583):571-9, 1996.
16) Hendrick JP, Hartl FU: Molecular chaperone functions of heat-shock proteins. Annu Rev Biochem. 62:349-84, 1993.
17) Jazwinski SM: Longevity, genes, and aging.
Science. 273(5271):54-9, 1996.
18) Kang Y, Eger W, Koepp H, Williams JM, Kuettner KE, Homandberg GA: Hyaluronan supresses fibronectin fragment-mediated damage to human cartilage explant cultures by enhancing proteoglycan synthesis. J Orthop Res 17: 858- 869, 1999.
19) Kawasaki K, Ochi M, Uchio Y, Adachi N, Matsusaki M: Hyaluronic acid enhances prolif-
eration and chondroitin sulfate synthesis in cul- tured chondrocytes embedded in collagen gels. J Cell Physiol. 179:142-148, 1999.
20) Laurent TC, Fraser JRE: Hyaluronan. FASEB J. 6:2397-2404, 1999.
21) Liu X, Engelman RM, Moraru II, Rousou JA, Flack JE 3rd, Deaton DW, et al: Heat shock. A new approach for myocardial preservation in car- diac surgery. Circulation. 86 Suppl 5:II358-63, 1992.
22) Laemmli UK: Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature. 227(259):680-5, 1970.
23) Marshall KW: The current status of hylan thera- py for the treatment of osteoarthritis. Today’s Therapeutic Trends. 15:99-108, 1997.
24) Minami Y, Hohfeld J, Ohtsuka K, Hartl FU:
Regulation of the heatshock protein 70 reaction cycle by the mammalian DnaJ homolog, Hsp40.
J Biol Chem. 271(32):19617-24, 1996.
25) Scale D, Wobig M, Wolpert W: Viscosupple- mentation of osteoarthritic knees with hylan: a treatment schedule study. Curr Ther Res. 55:220- 232, 1994.
26) Shukunami C, Shigeno C, Atsumi T, Ishizeki K, Suzuki F, Hiraki Y: Chondrogenic differen-
tiation of clonal mouse embryonic cell line ATDC5 in vitro: Differentiation-dependent gene expression of parathyroid hormone (PTH)/PTH- related peptide receptor. J Cell Biol. 133(2):457- 68. 1996.
27) Sohal RS, Weindruch R: Oxidative stress, caloric restriction, and aging. Science. 273(5271):59- 63, 1996.
28) Sonoda M, Harwood FL, Amiel ME, Moriya H, Temple M, Chang DG, et al: The effects of hylauronan in tissue healing after meniscus injury and repair in a rabbit model. Am J Sports Med. 28:90-97, 2000.
29) Tatar M, Khazaeli AA, Curtsinger JW: Chaper- oning extended life. Nature. 390(6655):30, 1997.
30) Toole BP: Hyaluronan is not just a goo! J Clin Invest. 106(3):335-6, 2000.
31) Ungewickell E, Ungewickell H, Holstein SE, et al: Role of auxilin in uncoating clathrin-coated vesicles. Nature. 378(6557):632-5, 1995.
32) Wobig M, Dickhut A, Maier R, et al: Visco- supplementation with hylan G-F 20: A 26-week controlled trial of efficacy and safety in the osteoarthritic knee. Clin Ther. 20:410-423, 1998.