DOI : 10.3341/jkos.2009.50.4.588
접수번호 : 08-16
각막윤부 줄기세포의 줄기세포성 유지 및 증식에 대한 Wnt 단백질의 효과
진현진⋅주천기
가톨릭대학교 의과대학 안과 및 시과학교실
목적: 각막윤부 줄기세포의 증식 및 줄기 세포성 유지에 대한 Wnt 단백질들의 효과에 대해 알아보고자 하였다.
대상과 방법: Wnt단백질의 발현은 western blot 분석을 이용하여 확인하였다. 각 Wnt에 의한 증식률은 콜로니 형성률로 확인하였고, Wnt에 의한 β-catenin활성은 면역형광 염색법으로 확인하였다. 각 Wnt에 의한 줄기세포성 유지 및 분화에 대한 효과는 p63과 connexin43에 대한 RT-PCR 이용하여 확인하였다.
결과: Wnt별 각막 윤부 줄기세포에 미치는 영향은 서로 상이하였으며, Wnt2와 Wnt4가 발현되는 배양액에서 성장한 각막윤부 줄기세 포가 대조군에 비해 유의적으로 높은 증식률을 보였다. Wnt2와 Wnt4에 영향을 받은 각막윤부 줄기세포에서 β-catenin이 핵에서 확인 되었다. p63은 Wnt2, Wnt4, Wnt5a, Wnt5b가 발현된 배양액에서 성장한 각막윤부 줄기세포에서 높게 발현되었다. 반면, Connexin43은 Wnt5a, Wnt5b, Wnt7b에서 높게 발현되는 것을 확인하였다.
결론: Wnt subtype 중 Wnt2와 Wnt4는 각막윤부 줄기세포의 증식과 줄기세포성 유지에 효과적일 것이라 판단된다.
<대한안과학회지 2009;50(4):588-593>
■ 접 수 일: 2008년 8월 25일 ■ 심사통과일: 2008년 11월 24일
■ 통 신 저 자: 주 천 기
서울시 서초구 반포동 505 가톨릭대학교 강남성모병원 안과 Tel: 02-2258-7620, Fax: 02-533-3801 E-mail: [email protected]
* 본 논문의 요지는 2008년 대한안과학회 제99회 춘계학술대회에서 구연으로 발표되었음.
* 이 논문은 2007년 가톨릭세포치료사업단 기초과학연구사업 연구비에 의하여 이루어졌음.
각막 상피세포는 안구표면의 투명성(integrity) 뿐만 아니 라 시각 기능에 매우 중요하다. 정상적인 상태에서 각막과 결막 상피세포 사이에 위치한 윤부의 기저막에 존재하고 있 는 줄기세포에 의해 유지되고 있다.1-3
화학 화상(Chemical injury) 또는 스티븐스-존슨 증후 군(Stevens-Johnson Syndrome)과 같은 병리적 또는 상 해로 부분적 또는 전체적인 윤부 줄기세포의 결핍을 가져 오며, 결과적으로 각막 상처 치유나 안구 표면 투명성에 대 한 심각한 결과를 초래 한다고 알려져 있다.4,5
병리적 또는 화학적 상해로 부분적 또는 전체적인 윤부 줄기세포의 결핍이 있을 때 이식될 수 있는 윤부 조직은 한 정되어 있고, 자가이식이 아닌 경우 면역 거부 등 많은 제 약이 따른다. 이러한 문제점을 극복하기 위하여 각막 윤부 줄기세포를 이용한 각막상피세포의 재건은 지난 수년간 많 은 연구자들에 의하여 연구되어 왔다.6-10
각막 윤부로부터 각막 윤부 줄기세포의 정확한 분리 및 추
출을 위하여 각막 윤부에서 줄기세포 표지자를 찾는 연구도 활발하다.11,12각막 상피 줄기세포를 이용한 치료는 시도되고 있으나5,13아직 임상에 적용하기에는 배양시간이 길고 준비 를 위한 세포의 수가 적다는 문제점이 있다. 또한 적은 수의 각막 윤부 줄기세포를 사용하기 위하여 줄기세포의 증식을 유도하는 방법으로 지속적인 계대 배양 방법을 사용하고 있 으나, 이러한 방법은 배양시간이 오래 걸리고, 증식하면서 분 화되는 줄기세포의 수도 증가한다.14,15
Wnt는 발생(embryonic development), 형태형성(morpho esis), 세포증식(cell proliferation), 분화(differentiation), 이 동(migration)에 중요한 역할을 담당하고 있다고 알려져 있다.16,17
Wnt protein은 세포막수용체인 Frizzled (fz)와 결합하여 Dsh (Disheveled)활성을 유도하고 이는 각각 Axin과 GSK-3 β를 비활성화 시킨다. 이로써 중간 전달자인 β-catenin은 세포질 내에서 안정화되고, 축적되어 핵 내로 이동하고 전 사인자(transcription factor)인 LEF-1/TCF family와 결 합하여 증식 또는 분화와 관련된 유전자의 발현을 활성화 시킨다. 최근 보고된 바에 의하면 Wnt는 줄기세포의 분열 과 분화에 영향을 주는 것이 관찰되었다.18-20
본 연구에서는 적은 수의 각막 윤부 줄기세포를 줄기세 포의 성격(stemness)을 유지시키면서 줄기세포의 수를 증 식시키기 위하여, Wnt 단백질을 이용하여 사람 각막 윤부 줄기세포의 증식을 유도하였고, Wnt단백질 중 효율적으로 증식을 유도하는 subtype의 종류를 찾고자 하였다.
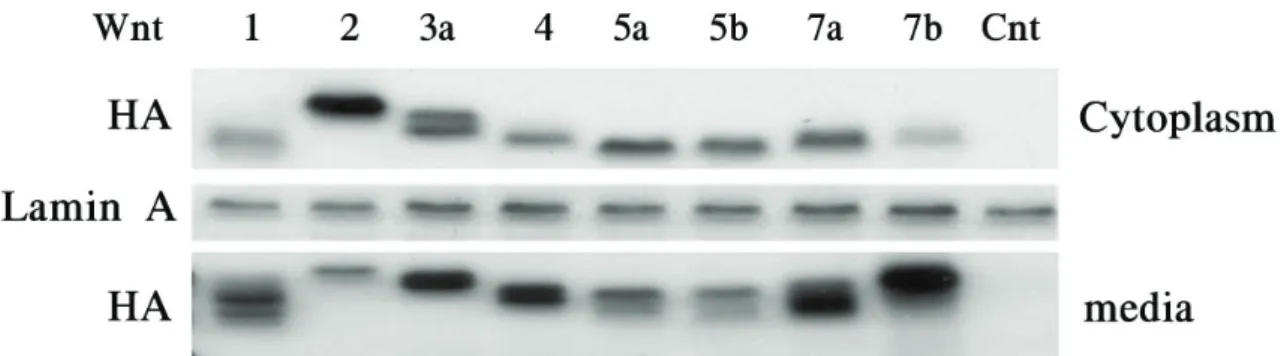
Figure 1. Wnt gene expression in NIH3T3 cells. HA‐Wnt protein expression was determined by western blot analysis. The protein was extracted from cytoplasm and media. HA was expressed Wnt trasnfected. Cnt as an empty vector‐transfected control.
대상과 방법
Wnt 단백질이 발현되는 NIH 3T3 Fibroblast feeder layer 준비
5% FBS (featal bovine serum albumin), 2 ng/ml hEGF, 5 ug/ml insulin, 5 ug/ml transferrin, 5 ng/ml sodiumselenite, 0.5 ug/ml hydrocortisone, 30 ng/ml cholera toxin, 0.5%
DMSO, 100 U/ml penicillin, 100 ug/ml streptomycin이 포 함된 DMEM/F12 (3:1)배지에 배양된 NIH3T3 fibroblast 지지세포(feeder cells)에 HA가 표지된 Wnt유전자를 가지는 pLNCX vector (Wnt1, 2, 3a, 4, 5a, 7a, 7b)와 대조군으로 pLNCX vector만을 각각의 배양 용기에 Lipofectamin-plus reagent (Invitrogen, Carlsbad, CA, USA)을 이용하여 3시 간 동안 전이(transfection) 시켰다. 24시간 동안 정상 배지 에서 배양한 후에, 세포의 증식을 억제하기 위하여 Mito- mycin C (MMC, 8 ug/ml)를 처리하고 37℃CO2배양기에 서 2시간 동안 반응시켰다.
각막윤부 줄기세포 배양
각막이식 후 남은 사람 윤부 조직은 가톨릭 한국인 눈 조 직 및 실명 관련 유전자 은행에서 분양 받아 사용하였다.
100 U/ml penicillin과 100 ug/ml streptomycin이 포함된 1X PBS로 한 번 세척한 뒤, 현미경상(SZ-40, Olympus) 에서 각막 내피세포, 홍채, 공막과 결막하 조직을 제거하였 다. 분리된 윤부 조직은 4등분하여 1.2 U/ml dispaseⅡ에 담가 37℃ CO2 Incubator에서 30분 동안 처리하고, 각막 윤부 상피세포층을 scraper를 이용하여 막 상태로 분리하 였다. 막으로 분리된 상피세포층을 단일세포로 분리하기 위 해 0.25% Trypsin-EDTA를 담가 37℃ CO2 배양기에서 30분간 반응시켰다. 이렇게 준비된 각막 윤부 상피세포를
5% FBS, DMEM/F12 (3:1)배지에 배양된 NIH3T3 feeder layer에 동시배양(co-culture)하였다.
Colony Forming Efficiency (CFE) assay
Wnt가 발현되는 NIH3T3 fibroblast 지지층위에 각막 윤 부 줄기세포 (2×104cells/well)를 약 10~13일 배양한 후, 인산완충용액(1X PBS, phosphate buffered saline, pH 7.4)으로 1차례 세척한 후에 0.53 mM EDTA 1 ml를 각 well에 첨가하고 37℃CO2배양기에서 1분간 반응시켜 지 지세포를 제거하였다. 인산완충용액으로 2차례 세척하고 99.85% methanol로 상온에서 5분간 반응시켜 세포 고정하 였다. 다시 한 번 인산완충용액으로 2회 세척한 뒤 0.1%
Crystal violet으로 실온에서 30분간 염색하였다. 광학현미경 으로 colony를 관찰하였고, colony 형성률은(Colony-forming efficiency (CFE)=number of colonies/number of seeded cells×100%)으로 표현하였다.
mRNA 분리 및 역전사 중합 반응(reverse transcription PCR)
형성된 colony 주변의 NIH3T3 feeder layer제거를 위해 0.53 mM EDTA (Welgen, Seoul, Korea)를 37℃1분간 반 응시켰다. 지지세포를 제거한 뒤 1X PBS로 1차례 세척한 뒤, 1 ml의 TRIzol Reagent (Invitrogen, Carlsbad, CA, USA)를 이용하여 total mRNA를 분리하였다. cDNA는 AMV reverse transcriptase kit (Promega, Madison WI, USA)를 이용하 여 합성하였다. p63은 Forward (5’-cag act caa ttt age gag-3’)와 Reverse (5’-agc tca tgg ttg ggg cac-3’)로 5 8℃에서 440bp로 하였고, Connexin43은 Forward (5’-cct tct tgc tga tcc agt ggt ac-3’)와 Reverse (5’-acc aag gac acc acc agc at-3’) 58℃154b로 반응시켰다. PCR에 의한 생성물은 1.5% agarose gel에서 전기영동하여 분석하였다.
A
B
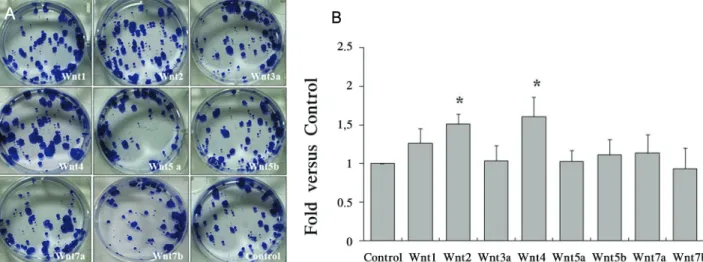
Figure 2. Proliferation of limbal stem cells by Wnt. (A) Proliferation capacity of Wnt was evaluated by 0.1% crystal violet staining at 13 days. (B) The colony formed in Wnt2 and Wnt4 higher in treated Wnt2 and Wnt4 compared with control. In addition, CFE was calculated as the number of colonies/number of inoculated cells. Colony counting (holoclone): diameter of colony>1 mm. Values represent the mean±S.E.M of three independent experiments. Significant difference (*p<0.05) compared with vector‐transfected control are represented by* for Wnt-treated. Control as a vector-transfected control. Abbreviation: CFE, colony‐forming efficiency.
면역형광 염색(Immunofluorescence staining)
각막 윤부 줄기세포을 배양하여 형성된 colony들을 인산 완충용액(1X PBS, phosphate buffered saline, pH 7.4)으로 1회 세척한 뒤 4% paraformaldehyde로 고정하였다. 인산 완충용액으로 2회 세척 후에 항체의 비특이적 결합을 억제 하기 위해 2.5% horse serum (Vector Lab, Burlingameb, CA, USA)로 10분간 상온에서 반응시켰다. Horse serum 을 제거한 후 1% horse serum이 포함된 인산완충용액에 일차 항체 anti-β-catenin을 2,000배 희석하여 처리한 뒤 4℃에서 12시간 반응시켰다. 인산완충용액으로 2차례 세척 하고, 2차 항체는 alexa 546으로 표지된 Goat-anti-rabbit (Santa Cruz, Santa Curz, CA, USA)을 1% horse serum이 포함된 인산완충용액에 400배 희석하여 상온에서 반응시 켰다. 인산완충용액으로 2회 세척한 뒤, 핵을 염색하기 위 해 Hoechst 33342 (Sigma, Louis, Missouri, USA)를 인산 완충용액에 500배 희석하여 상온에서 10분간 반응시킨 후, Flourescence Microscope (Axiovert 200, Zeiss)을 이용 하여 관찰하였다.
결 과
지지세포에서 각 Wnt subtype의 발현
NIH3T3 fibroblast 지지세포에 Wnt 유전자를 전이시킨 후, Wnt 단백질이 발현 되는지를 western blot분석법을 이용
하여 확인하였다. Vector만 전이시킨 대조군에서는 HA가 발 현되지 않았으며, 각각의 Wnt를 전이시킨 군에서는 세포질 (cytoplasm) 및 배양액(media)에서 HA가 발현되는 것을 확인함으로써 Wnt가 발현되는 것을 확인하였다(Fig. 1).
Lamin A는 단백질 발현의 대조군으로 사용하였다.
Wnt 단백질에 의한 각막 윤부 줄기세포의 증식 유도
각 Wnt1, 2, 3a, 4, 5a, 5b, 7a, 7b가 발현되는 지지세포와 동시 배양된 각막 윤부 줄기세포에서 콜로니 형성률(colony forming efficiency)을 확인하였다. 그 결과 Wnt2와 Wnt4 가 발현되는 지지세포와 배양된 군에서 다른 Wnt에 비해 콜 로니 형성이 많은 것이 관찰되었다. 또한 직경 1 mm 이상 되는 콜로니들은 pLNCX-vector만을 처리한 대조군에 비 해 Wnt2와 Wnt4에서 유의적으로 높은 콜로니 형성률을 보였다(Fig. 2).
배양된 각막윤부 줄기세포에서 ß-catenin 면역염색
Wnt에 의한 β-cantenin의 활성을 확인하기 위하여, 각막 윤부 줄기세포의 증식에 효과를 보인 Wnt2와 Wnt4에서 β -cantenin에 대한 면역염색을 실행하였다(Fig. 3). Wnt 단백 질이 없는 배지에서 증식된 각막윤부 줄기세포에서는 β -cantenin이 세포질과 세포막에 위치되어 있는 것을 관찰 할 수 있었다. 그러나 Wnt2와 Wnt4가 있는 배지에서 성장한 각 막윤부 줄기세포에서는 β-cantenin이 핵 내에서 관찰되었다.
A
B
C
Control β-catenin Merged
Wnt2 β-catenin Merged
Wnt4 β-catenin Merged
Figure 3.Nuclear translocation of β‐catenin in the colony induced by Wnt2 and Wnt4. The localization of β‐catenin was examined by immunocytochemistry at 11 days. (A) β‐catenin is observed in the cytoplasm and plasma membrane.
(B)(C) Wnt2 and Wnt4 treated cells show nuclear accumulation of β‐catenin. β‐catenin proteins were detected as a red color using anti‐rabbit alexa 546. Cell nuclei were stained with Hoechst 33342 Scale bar=10 μm.
Wnt에 의한 줄기세포성과 분화 관련 유전자의 발현
각 Wnt1, 2, 3a, 4, 5a, 5b, 7a, 7b가 발현되는 지지세포 에서 증식한 각막 윤부 줄기세포에서 분화된 각막 상피 세 포 표지인자로 알려진 Connexin43 (Cx43)과 각막상피 줄 기세포의 표지인자인 p63에 대한 유전자 발현의 차이를 RT-PCR법을 이용하여 관찰하였다(Fig. 4). 대조군(control) 과 Wnt 단백질이 포함된 배양액에서 증식한 각막 윤부 줄기 세포에 Cx43과 p63의 유전적 발현을 비교해 볼 때 각막 윤부 줄기세포 표지인자인 p63의 발현은 Wn2, Wnt4, Wnt5a Wnt5b에서 높았으며, 그 중 Wnt4와 Wnt5a에서 가장 높은
발현을 보였고, 분화 표지인자인 Cx43은 Wnt5a, Wnt5b, Wnt7b에서 가장 높은 것을 확인하였다.
고 찰
현재 각막상피줄기세포를 이식을 위하여 사용하기에는 줄기세포의 분리, 증식과 이식을 위한 준비 기간이 길고, 각 막상피 줄기세포를 많이 얻을 수 없다는 문제점이 있다.8,9 이러한 문제점을 해결하기 위한 한 가지 방법으로 줄기세 포의 성격을 유지하면서 증식을 유도하는 방법이 많은 연 구자들에 의하여 연구되어지고 있다.10,14
p63 Connexin43 GAPDH
Wnt1 Wnt2 Wnt3a Wnt4 Wnt5a Wnt5b Wnt7a Wnt7b Cut
Figure 4.Expression of limbal stem cells and differentiation marker in cultured limbal stem cells. RT‐PCR profiles showing mRNA expression of Connexin43 and p63 with GAPDH as an internal control. Limbal stem cell marker p63 mRNA was strongly expressed in Wnt4 and Wnt5a.
Otherwise, Differentiation marker Connexin43 mRNA strongly expressed in Wnt5a, Wnt5b, Wnt7b.
본 연구에서 사람의 각막 윤부로부터 분리된 각막 윤부 줄기세포(Limbal stem cells, LSCs)를 증식시키는 방법으 로 현재 줄기세포의 조절인자로 알려진 Wnt 단백질을 사용 하였고, Wnt 단백질의 종류 중 Wnt2와 Wnt4가 각막 줄기 세포의 증식에 영향을 미치는 것으로 관찰되었다.
Wnt는 canonical과 noncanonical pathway의 2가지 신호 전달 기전에 의하여 세포의 분화 및 증식을 조절하는 데, 줄기세포에 있어서는 β-cantenin이 관여하는 canonical pathway가 잘 알려져 있다.19,20본 연구에서도 Wnt가 포함 된 배양액에서 자란 각막 윤부 줄기세포에서 β-cantenin의 활성을 확인할 수 있었다.
Wnt는 조직에서 줄기세포의 증식을 조절하거나 분화를 유도하는 것으로 알려져 있고, 최근 신경, 근육, 피부 등의 줄기세포에서 발현이 관찰되고 있고,18,21또한 β-cantenin 의 활성을 유도하여 줄기세포의 분열을 촉진한 연구가 여 러 성체 줄기세포 연구에서 보고되고 있어 각막상피줄기 세포의 증식을 유도하고 조절하는 데 유용한 조절인자로 판단된다.
본 연구에서는 각 Wnt를 3T3 feeder 세포에 전이(trans- fection)시켜 배지에 분비되게 하는 방법으로 Wnt를 각 막상피 줄기세포에 공급하였다. 현재 많은 연구에서 β- cantenin을 줄기세포에 전이시키거나 활성 시켜 줄기세포 의 증식과 분화를 유도하는 데, 이것은 Wnt 신호전달기전 중 β-cantenin활성에 중점을 두고 이루어진 방법으로 Wnt 에 의한 줄기세포의 증식이라고는 할 수 없다.
본 연구에서 확인한 결과 모든 Wnt 단백질이 증식을 유 도하지 않았고, Wnt subtype별로 각막상피줄기세포에 대한 역할이 각각 상이하였다. 즉, Wnt2와 Wnt4는 줄기세포의 증식유도에 효과를 나타내었으며, 줄기세포성(stemness) 유지에서는 증식유도에 효과를 보인 Wnt2와 Wnt4 뿐만 아니라, Wnt5a, Wnt5b에서 대조군에 비해 효과를 보였다.
반면, Wnt5a, Wnt5b, Wnt7b는 분화를 유도하였다.
줄기세포에서 Wnt의 역할을 연구한 여러 논문에서와 같 이 본 연구에서도 Wnt가 줄기세포의 증식과 줄기세포의 성격을 유지한다는 결과를 얻었지만, Wnt의 종류에 따라 그 기능이 상이하다는 결론을 본 연구를 통하여 얻을 수 있었다.
현재 Wnt 단백질은 19개가 알려져 있지만, 본 연구에서 저자들은 모든 Wnt subtype별로 각막상피줄기세포에 대한 영향을 관찰하지는 못하였다. 따라서 이 연구에서 확인되지 않은 다른 Wnt 단백질 중에서도 줄기세포의 증식과 분화에 대한 연구가 필요할 것으로 판단된다.
결론적으로, Wnt 단백질을 각막 윤부 줄기세포의 성장 인자로써 사용 가능하고, 각 Wnt subtype의 특성을 이용하 여 증식과 분화를 조절할 수 있을 것으로 판단된다. 특히, Wnt2와 Wnt4는 각막윤부 줄기 세포의 증식과 줄기세포성 유지에 효과적일 것이라 판단된다.
참고문헌
1) Davanger M, Evensen A. Role of the pericorneal papillary structure in renewal of corneal epithelium. Nature 1971;229:
560-1.
2) Daniels JT, Dart JK, Tuft SJ, et al. Corneal stem cells in review.
Wound Repair Regen 2001;9:483-94.
3) Revoltella RP, Papini S, Rosellini A, et al. Epithelial stem cells of the eye surface. Cell Prolif 2007;40:445-61.
4) Kenyon KR, Tseng SC. Limbal autograft transplantation for ocular surface disorders. Ophthalmology 1989;96:709-22.
5) Frucht-Pery J, Siganos CS, Solomon A. Limbal cell autograft transplantation for severe ocular surface disorders. Graefes Arch Clin Exp Ophthalmol 1998;236:582-7.
6) Kim MK, Lee JL, Shin KS, et al. Isolation of putative corneal epithelial stem cells from cultured limbal tissue. Korean J Ophthalmol 2006;20:55-61.
7) Park KS, Lim CH, Min BM, et al. The side population cells in the rabbit limbus sensitively increased in response to the central cornea wounding. Invest Ophthalmol Vis Sci 2006;47:892-900.
8) Dogru M, Tsubota K. Current concepts in ocular surface recon- struction. Semin Ophthalmol 2005;20:75-93.
9) Shortt AJ, Secker GA, Notara MD, et al. Transplantation of ex vivo cultured limbal epithelial stem cells: a review of techniques and clinical results. Surv Ophthalmol 2007;52:483-502.
10) Liu J, Song G, Wang Z, et al. Establishment of a corneal epithelial cell line spontaneously derived from human limbal cells. Exp Eye Res 2007;84:599-609.
11) Chee KY, Kicic A, Wiffen SJ. Limbal stem cells: the search for a marker. Clin Experiment Ophthalmol 2006;34:64-73.
12) Schlötzer-Schrehardt U, Kruse FE. Identification and characteri- zation of limbal stem cells. Exp Eye Res 2005;81:247-64.
13) Ivekovic R, Tedeschi-Reiner E, Novak-Laus K, et al. Limbal graft and/or amniotic membrane transplantation in the treatment of ocular burns. Ophthalmologica 2005;219:297-302.
14) Vossmerbaeumer U, Kuehl S, Bieback K, et al. Cultivation and
=ABSTRACT=
The Effects of Wnt Protein on Proliferation and Stemness Maintenance of Corneal Limbal Stem Cells (CLSCs)
Hyun-Jin Jin, BS, Choun-Ki Joo, MD, PhD
Department of Ophthalmology and Visual Science, College of Medicine and Laboratory of Ophthalmology and Visual Science, Catholic Research Institutes of Medical Science, The Catholic University of Korea, Seoul, Korea
Purpose: To evaluate the effects of the Wnt protein on proliferation and stemness maintenance of cultured corneal limbal stem cells.
Methods: We examined the expression of Wnt proteins by Western blot analysis. We then evaluated the effects of Wnt on cell proliferation by colony forming efficiency. β-catenin activation using Wnt proteins was examined by immunocytochemistry. We also examined the effects of Wnt on proliferation and stemness maintenance by reverse transcriptase polymerase chain reaction of p63 and connexin43.
Results: Wnt has a different effect on corneal epithelial stem cells. Colony forming efficiency was also significantly higher in treated Wnt2 and Wnt4 cells compared with controls. The Wnt2 and Wnt4 treated cells showed nuclear accumulation of β -catenin. In addition, the limbal stem cell marker p63 was strongly expressed in Wnt2, Wnt4 Wnt5a, and Wnt5b. Connexin43 mRNA was also strongly expressed in Wnt5a, Wnt5b and Wnt7b cells.
Conclusions: We suggest that Wnt2 and Wnt4 could lead to more effective proliferation and stemness maintenance for human corneal epithelial stem cells.
J Korean Ophthalmol Soc 2009;50(4):588-593
Key Words: Limbal stem cell, Proliferation, Stemness, Wnt protein
Address reprint requests to Choun-Ki Joo, MD, PhD
Department of Ophthalmology and Visual Science, St. Mary’s Hospital, College of Medicine, The Catholic University of Korea
#505 Banpo-dong, Seocho-gu, Seoul 137-040, Korea
Tel: 82-2-2258-7620, Fax: 82-2-533-3801, E-mail: [email protected] differentiation characteristics of human limbal progenitor cells.
Tissue Cell 2008;40:83-8.
15) Zhao B, Allinson SL, Ma A, et al. Targeted cornea limbal stem/
progenitor cell transfection in an organ culture model. Invest Ophthalmol Vis Sci 2008;49:3395-401.
16) Moon RT, Brown JD, Torres M. Wnts modulate cell fate and behaviour during vertebrate development. Trends Genet 1997;13:
157-62.
17) Logan CY, Nusse R. The Wnt signaling pathway in development and disease. Annu Rev Cell Dev Biol 2004;20:781-810.
18) Nusse R. Wnt signaling and stem cell control. Cell Res 2008;
18:523-7.
19) Willert K, Nusse R. Beta-catenin: a key mediator of Wnt signaling.
Curr Opin Genet Dev 1998;8:95-102.
20) Gordon MD, Nusse R. Wnt signaling: multiple pathways, multiple receptors, and multiple transcription factors. J Biol Chem 2006;
281:22429-33.
21) Hoppler S, Kavanagh CL. Wnt signalling: variety at the core. J Cell Sci 2007;120:385-93.