PREVENTION RESEARCH □ ORIGINAL ARTICLE □
265 책임저자:권현주, 614-714, 부산시 부산진구 가야동 산 24
동의대학교 자연과학대학 생명응용학과 Tel: 051-890-1519, Fax: 051-890-1532 E-mail: [email protected]
접수일:2009년 9월 8일, 게재승인일:2009년 9월 21일
Correspondence to:Hyun Ju Kwon
Department of Life Science and Biotechnology, College of Natural Science, Dongeui University, San 45, Gaya-dong, Busanjin-gu, Busan 614-714, Korea
Tel: +82-51-890-1519, Fax: +82-51-890-1532 E-mail: [email protected]
Widdrol의 인간 폐암 세포주에 대한 세포주기 정지 및 MCM 발현 억제 효과
1동의대학교 자연과학대학 생명응용학과, 2동의대학교 블루바이오 소재개발센터,
3동의대학교 대학원 바이오물질제어학과, 4동의대학교 생활과학대학 식품영양학과
홍윤경1ㆍ오유나2ㆍ현숙경2ㆍ윤희정3ㆍ황혜진2,4ㆍ이은우1,2ㆍ김병우1,2,3ㆍ권현주1,2,3
Widdrol Induces Cell Cycle Arrest and MCM Down Expression in Human Lung Carcinoma Cells
Yun Kyung Hong1, Yu Na Oh2, Sook Kyung Hyun2, Hee Jung Yun3, Hye Jin Hwang2,4, Eun Woo Lee1,2, Byung Woo Kim1,2,3 and Hyun Ju Kwon1,2,3
1Department of Life Science and Biotechnology, College of Natural Science, 2Blue-Bio Industry Regional Innovation Center,
3Department of Biomaterial Control (BK21 program), Graduate School, 4Department of Food and Nutrition, College of Human Ecology, Dongeui University, Busan 614-714, Korea
We report here the widdrol, an odorous compound extracted from Juniperus chinensis, induced cell cycle arrest in human lung carcinoma A549 cells. We found the widdrol significantly induced cell-cycle arrest in G1 phase. To investigate the possible mechanisms, G1-related proteins were assayed by western blot analysis. Widdrol treatment resulted in the up-regulation of tumor suppressor p53 and cyclin-dependent kinase (CDK) inhibitor p21. In contrast, the expressions of cyclin-dependent kinase (CDK2), cyclin E, retinoblastoma protein (pRB) and proliferating cell nuclear antigen (PCNA) levels were markedly decreased in the concentration-dependent manner. Moreover, the minichromosome maintenance (MCM) proteins, highly expressed in cancer cells, were significantly down-regulated in A549 cells treated with widdrol. Overall, these observations suggest widdrol induces antiproliferation and arrest G1 phase in A549 cells, possibly mediated by the inhibition of the MCM protein expression. Consequently, widdrol is a potential candidate for the anticancer drug development. (Cancer Prev Res 14, 265-273, 2009) Key Words: Widdrol, MCM protein, A549 cells, G1 arrest, CDK inhibitor
서 론
세포는 여러 가지 세포외 시그널을 받게 된다. 그 중요 한 반응 중 하나가 세포증식을 위한 것이며 또 다른 시 그널은 세포증식 정지 신호이다. 이런 시그널은 세포막 에서 세포질을 지나 핵으로 전달되며 세포주기 조절계
를 제어한다. G1, S, G2, M의 세포주기 중에서 세포 외 시그널이 전달되는 시기는 주로 G1, G2기로 DNA 손상 또는 복제 이상을 감지하여 이상이 있을 경우 G1/S 이행 시기와 G2/M 이행시기의 checkpoint에서 세포주기를 지 연시키고 수리기작을 작동한다.1) 암세포는 세포주기제 어 기작 어느 곳의 이상으로 인해 정지신호를 받지 않게 된 세포이다. 이미 알려진 많은 세포 독성 물질이나
DNA를 손상시키는 물질들은 세포주기를 정지시켜 apo- ptosis를 유도 한다.2,3) 사실상, 많은 항암제의 특성을 살 펴보면 세포주기 정지 유도 또는 세포의 자가사멸을 유 도하는 경우가 대부분이다.
세포주기 진행을 담당하고 있는 주요 단백질로는 cyclin dependent kinase (CDK)와 이들의 활성을 조절하는 cyclin이 heterodimer 형태로 결합하여 holoenzyme 기능을 하며 G1, S, G2, M기에 작용한다.4,5) Cyclin-CDK 복합체가 세포주기를 정방향으로 진행시키는 역할을 담당한다면 여러 가지 외부, 내부 요인의 증식억제 신호에 반응하여 세포주기를 정지시키는 역할은 cyclin dependent kinase inhibitor (CKIs)가 담당한다. 포유동물에서는 지금까지 7 종류의 CKI를 코드하는 유전자가 밝혀져 있으며 이들 유전자의 구조상 Cip/Kip family와 Ink4 family 2가지로 구 분된다.6,7) p21, p27 및 p57이 속하는 Cip/Kip family는 cyclin-CDK 복합체에 결합하여 kinase활성을 저해하고 이 들 단백질의 발현이 증가되면 배양세포에서 G1기 정지 가 잘 관찰된다. Ink4 family에는 p16, p15, p18 및 p19가 있으며 CDK4/6에 대해 cyclin D와 경쟁적으로 결합함으 로써 G1기 정지를 유도시킨다.8) p21은 DNA 손상 시그 널을 받은 암억제 유전자 p53에 의해 전사 유도되어 발 현되며 cyclin-CDK 복합체에 대해 저해작용을 가지고 있 다. 또한 DNA 복제개시에 있어 필수적인 PCNA 단백질 에 결합하여 S기 개시를 방해 하는 등 G1기 이외에도 여러 지점에서 세포주기 제어에 관여하고 있다.9) Minichromosome maintenance (MCM) 단백질은 효모에서 복제분기점 활성화와 세포 분화 과정에 필수 단백질로 처음 동정 되었으며 진핵세포 내에 공통으로 존재하는 단백질이다. 또한 MCM 단백질은 복제기점에서 helicase 활성으로 DNA 복제를 개시하는 중요 단백질이며 세포 주기 당 1회의 DNA가 복제되도록 하는 DNA replication licensing factor이다.10∼12) 뿐만 아니라, 여러 암세포에서 MCM 단백질의 과발현이 발견되었으며 이것은 복제 개 시점의 계속적인 활성화로 over-replication으로 이어질 가 능성이 높으며 게놈 DNA의 불안정화를 초래할 수 있
다.13,14) 그 외에도 각각의 MCM 단백질은 전사, chromatin
의 형성, 세포주기 checkpoint 등의 역할도 담당하고 있 다.15) 이와 같이 MCM 단백질은 매우 다양한 세포내 기 능을 가지고 있을 뿐만 아니라, 최근에는 MCM 단백질이 종양형성에도 관련이 있는 것으로 시사되어 많은 연구 들이 여기에 초점을 맞추고 있다. 실제로 여러 종류의 암세포에서 MCM 단백질이 정상세포에 비해 약 10∼20 배 정도 과발현 되는 현상이 관찰되었으며, 이러한 MCM 단백질의 과발현이나 기능 이상은 복제조절 실패로 이
어져 세포는 암화 되는 것으로 보고되어 있다.16∼20) 본 연구팀은 MCM 단백질 발현 저하 물질을 스크리닝 하는 중 Juniperus chinensis로부터 단일 성분 widdrol을 분리 하였다. Widdrol은 Juniperus chinensis을 포함하는 Juniperus sp.
와 같은 다양한 식물에 함유 되어 있는 자연 재료로 결 정화 되어진 물질이며 Botrytis cinerea에 대한 항진균성 활 성이 보고된바 있다.17∼19) 본 연구에서는 MCM 단백질의 발현 저하 기작을 연구 하던 중 인간 폐암 세포 A549에 서 MCM 단백질의 발현 저하와 함께 항암활성인 세포주 기 정지 효과를 관찰하였으므로 그 결과를 보고하는 바 이다.
재료 및 방법 1. 세포배양
인간 폐암세포인 A549는 한국 세포주 은행에서 구입 하여, RPMI 1640배지에 10% (v/v)의 FBS와 0.1% genta- mycine을 첨가한 배지를 사용하였으며 37oC, 5% CO2 조 건하에서 배양 하였다.
2. Widdrol 처리
Juniperus chinensis로 부터 분리되어진 widdrol은 1H NMR,
13C NMR 그리고 GC-MS를 통해 확인하였다. Widdrol은 dimethyl sulfoxide (DMSO)에 용해하였으며 −20oC에서 보 관하여 사용하였고 widdrol의 희석은 culture medium에서 수행하였다.
3. 세포성장 억제
A549 세포를 35 mm dish에 1×104 cells/ml이 되도록 분 주하여 24시간 후 10 μg/ml widdrol을 처리하였다.
Widdrol 처리 후 12, 24, 36 및 72시간 별로 cell을 회수하 여 trypan blue (Trypan blue solution 0.4%, SIGMA)시약으로 1:1 염색하였고 hemocytometer를 이용하여 생세포 수를 측정하였다.
4. 세포주기 분석
Widdrol 처리에 따른 A549의 세포주기는 flow cyto- metry를 사용하여 분석하였다. 35 mm dish에 5×105 cells 의 농도로 분주하여 widdrol을 농도별(0, 5, 10, 15, 20 μg/
ml)로 처리하고 대조군은 0.1%의 DMSO를 첨가하여 48 시간 배양하였다. 배양한 세포를 회수 한 후 PBS로 세척 하고 100% EtOH을 이용하여 4oC에서 고정하였다. 고정 된 세포는 분석 전에 PBS로 세척하고 ribonuclease A를 최 종농도가 1 μg/ml이 되도록 가해 실온에서 30분간 반응
시켰다. PI/RNase staining buffer (BD PharmingenTM)용액을 500 μl 첨가한 후 유세포 분석기(FACS, Becton Dickinson, San Jose, CA)로 측정하였으며 결과는 Cell Quest program 을 이용하여 분석하였다.
5. Microarray 분석
Widdrol을 처리한 A549세포와 처리하지 않은 대조군 A549세포의 유전자발현의 차이를 (주)지노첵에서 제작 된 operon human whole 37K oligo chip을 사용하여 확인하 였다. 총 RNA 표본은 역전사 과정을 통하여 특정한 primer가 부착된 cDNA를 합성하였으며 이것을 platinum oligo chip에 1차 융합을 실시하였다. 그리고 2차 융합 과 정 시 labeling하였다. 본 실험에서 사용된 지노첵 plati- num biochip human cancer 3.0K oligo chip은 Qiagen Operon 에서 제공하는 human cancer oligo set으로, 이미 알려진 유전자 2,959개, 기능이 알려지지 않은 EST 유전자 81개, housekeeping gene 및 대조군 유전자로 arabidopsis DNA를 점적한 총 3096개의 스폿이 포함되어 있으며 이들은 1개 의 block에 23개의 column과 23개의 row로 구성되어 있 다. 각 oligo nucleotide는 (주)지노첵에서 제공하는 spotting 용액 15 μl에 용해시켜 pixsys 5500 arrayer (Cartesian Technologies, CA)를 사용하여 24 Stealth Micro spotting pins 으로 CMT-GAPS II silane slide glass (Corning, NY)에 점적 하였다. 점적한 슬라이드는 1×SSC 용액으로 1분간 재수 화 하고 UV crosslinker (Stratagene, CA)로 DNA를 슬라이 드에 링크시켰다. 다음은 슬라이드를 succinic anhyd- ride/sodium borate 용액으로 적당히 교반하면서 15분간 적신 다음 95oC, water bath에 2분간 방치한다. 그 후 슬라 이드를 재빨리 95% ethanol에 1분간 옮기고 3,000 rpm에 서 20초간 원심분리 하여 건조시킨다. DNA 칩 분석을 위한 총 RNA의 추출은 widdrol을 48시간 처리한 A549 세 포와 처리하지 않은 대조군 A549 세포를 TRI REAGENT (MRC, OH)를 사용하여 추출하였다. 형광 label cDNA probe는 40 μg의 총 RNA를 SuperScript II reverse trans- criptase (Invitrogen, NY)를 사용하여 oligo (dT)18-일차 중합 반응으로 제작하였다. 역전사 반응액은 400 U SuperScript RNase H-reverse transcriptase (Invitrogen), 0.5 mM dATP, dTTP와 dGTP, 0.2 mM dCTP와 0.1 mM Cy3 또는 Cy5가 label된 dCTP (NEN Life Science Product Inc.)를 포함하고 있다. 역전사 반응 후 표본 RNA는 5 μl의 stop solution (0.5 M NaOH/50 M EDTA)을 가하여 65oC에서 10분간 배 양함으로써 분해시킨다. Label된 cDNA 혼합액은 ethanol 침전법으로 농축한다. 농축된 Cy3와 Cy5로 label된 cDNAs 20 μl hybridization solution (GenoCheck, KR)에 재용해시킨
다. 두 cDNA는 혼합하고 95oC에서 2분간 변성시킨 후 45oC water chamber에서 20분간 반응시킨다. 그 후 cDNA 혼합액을 spotted slide에 놓고 커버슬립(22×22 mm)으로 덮는다. 슬라이드를 62oC에서 12시간 hybridization cham- ber에서 혼성화시킨다. 혼성화된 슬라이드는 2×SSC와 0.1% SDS 용액으로 2분간 세척 후 상온에서 1×SSC 용액 으로 3분, 0.2×SSC 용액으로 2분 세척한다. 그 다음 슬라 이드를 3,000 rpm에서 20초간 원심분리하여 건조한다.
혼성화 슬라이드는 Axon Instruments GenePix 4000B 스캐 너로 스캔하여 이미지를 GenePix Pro 5.1 (Axon, CA)과 GeneSpring 6.1 (Sillicongenetics, CA), R 패키지 프로그램을 사용하여 분석하였다. DNA 칩 결과의 분석 과정은 스캐 닝을 통하여 얻어진 스폿의 signal intensity 수치 데이터를 사용하여 global normalization과 intensity dependent normalization를 실시하였다. 또한 칩의 각 블럭에 대한 차 이를 최소화하기 위하여 block-wise 표준화를 실시하였 다. 실험 결과로 얻어진 모든 유전자들의 분포를 히스토 그램으로 표시하여 표준화가 끝난 데이터의 분포도와 비교 검토하여 실험 결과의 정확성을 확인하였다.
6. Western blot analysis
Widdrol이 처리된 A549 세포를 CSK buffer (10 mM Pipes, pH 6.8, 100 mM NaCl, 1 mM MgCl2, 1 mM EGTA, 1 mM dithiothreitol, 1 mM phenyl methane sulfonyl fluoride) 에 0.1% triton X-100, 1 mM ATP와 protease inhibitor (Phar- minogen A)가 첨가된 용액을 사용하여 현탁한 후 soni- cation을 행하여 세포를 파쇄하였다. 파쇄된 세포는 20,000× g로 30분간 원심 분리하여 상층액을 획득하였으 며, 상층액의 단백질 농도는 BCA protein assay kit (Bio- Rad)를 사용하여 측정하였다. 동일 량의 단백질(40 μg) 을 사용하였고, actin은 대조군으로 사용하여, SDS-PAGE 를 행하였다. p21, PCNA와 actin은 15%, PARP는 8%의 분 리 겔을 사용하였으며 p53, CDK2와 cyclin E는 12%, MCM-2, -3, -4, -6, -7, pRB와 p-pRB는 10% 분리 겔을 사용 하였다. 단백질 전기영동 후 겔 내의 단백질은 PVDF 막 에 전사시키고 blocking solution (BlockaceTM, Dai-Nippon)을 사용하여 실온에서 1시간 blocking 과정을 거쳤다. 1차 항 체는 4oC에서 16시간 반응시켰으며 TBS (50 mM Tris/HCl, pH 7.5, 0.15 M NaCl)에 0.1% Triton X-100이 첨가된 용액 을 사용하여 PVDF 막을 세척하였다. Peroxidase-conjugated 이차 항체(Pierce)를 사용하여 4oC에서 16시간 반응시키 고 chemilluminescence system (Super- Signal West Femto Maximum sensitivity Substrate, Pierce)으로 검출하였다. 반 응의 정도는 FluorchemTM5500 (AlphaInnotech)을 사용하여
Fig. 1. Growth inhibition of widdrol on A549 cells. 10 μg/ml (IC50) of widdrol were treated on A549 cells and cells were collected in the indicated time periods. Viable cells were determined by trypan blue exclusion assay and calculated to the percentage of cell growth inhibition.
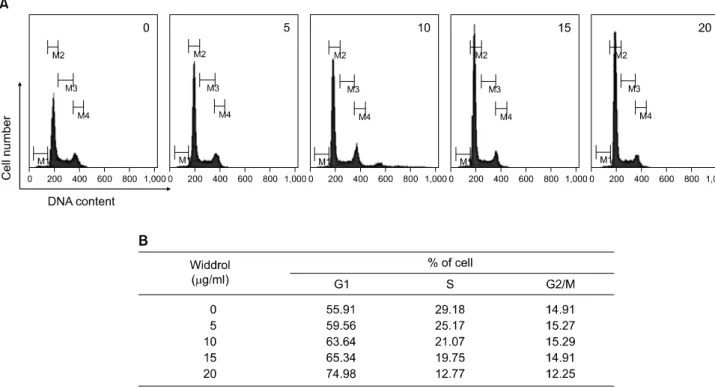
Fig. 2. Induction of G1 cell cycle arrest by widdrol in A549 cells. A549 cells treated by widdrol for 48 h were collected, stained by PI, and analysed by flow cytometry. (A) G1 areas of cells treated with widdrol increased in the concentration-dependent manner compare to the control cells (no treatment of widdrol). (B) The changes of G1, S, and G2/M were calculated by Cell Quest program.
Data are representative results from three independent experiments.
정량 하였다. Actin, p53, p21, CDK2, cyclinE, pRB, MCM-2, -3, -6, -7, PCNA 및 E2F1 항체는 Santa Cruz Biotechnology, Inc. (Santa Cruz, CA)에서 MCM-4단백질의 항체는 BD Bio- sciences Pharminogen (BD PharmingenTM, CA)에서 구입하였
으며 p-pRB는 Cell Signaling Technology (Bererly, MA)에서 구입하였다.
7. Immunoprecipitation analysis
Widdrol을 처리한 암세포를 CSK buffer에 0.1% TritonX- 100, 1 mM ATP와 Protease inhibitor (Pharminogen A)가 첨 가된 용액을 사용하여 lysis 시킨 후 BCA protein assay kit (Bio-Rad)를 사용하여 500 μg의 단백질을 검출하였다. 1 μg 항체와 단백질을 함께 4oC에서 1시간 처리한 후 immune complexes는 protein A-agarose (Santa Cruz Biothech- nology Inc.)와의 혼합을 통해 면역침전으로 획득하였다.
면역침전을 통해 분리된 복합체는 western blot analysis를 시행함으로써 각 복합체의 존재 정도를 확인하였다.
결과 및 고찰 1. Widdrol의 세포 성장 억제 효과
Widdrol의 A549 세포에 대한 성장억제효과를 측정하 기 위하여 IC50 10 μg/ml을 처리 한 후 3일 동안 배양하 여 변화를 확인하였다. 12시간 별로 cell을 회수하여 trypan blue 염색한 후 hemocytometer에서 생세포수를 측
Table 1. Expression pattern of the cell cycle related genes in A549 cell after treated with widdrol Gene
symbol Description Expression
fold
Gene bank no.
TP73 Tumor suppressor p73 +1.04 NM_005427
AbL2 v-abl Abelson murine leukemia viral oncogene homolog 2 (Abelson-related gene) +0.55 BX538317
CDKN1A Cyclin-dependent kinase inhibitor 1A (p21, Cip1) +1.13 NM_078467
CCND3 Cyclin D3 −0.59 AK096276
CDK6 Cyclin-dependent kinase 6 −0.34 NM_001259
E2F1 E2F transcription factor 1 +1.47 BC050369
FOS v-fos FBJ murine osteosarcoma viral oncogene homolog −0.67 BX647104
JUN v-jun sarcoma virus 17 oncogene homolog (avian) −0.44 NM_002228
MYCN v-myc myelocytomatosis viral related oncogene, neuroblastoma derived (avian) −0.53 NM_005378
PIAS2 Protein inhibitor of activated STAT, 2 +1.08 NM_004671
GADD45A Growth arrest and DNA-damage-inducible, alpha +0.67 CR612719
PCNA Proliferating cell nuclear antigen −0.27 NM_182649
TOP2B Topoisomerase (DNA) II beta 180 kDa −0.40 NM_001068
정하였다. Fig. 1에서 보는 바와 같이 widdrol을 처리한 세 포는 12시간째부터 대조군 100%에 비해 48.5%의 생존율 을 나타내며 처리 시간에 따라 생존율이 감소하여 48시 간 이상 처리하면 20%이하의 생존율을 보였다. 따라서 폐암세포에 대한 widdrol의 성장 억제 활성을 확인할 수 있었다.
2. Widdrol에 의한 세포주기 변화
Widdrol이 세포주기에 어떠한 영향을 나타내는지 조 사하기 위하여 A549 세포에 widdrol을 농도 별로 48시간 처리한 후 세포를 회수하였다. PI염색을 한 후 flow cytometry를 사용하여 각 주기별로 정량 분석하였다. 그 결과 Fig. 2에서 보는 바와 같이 5 μg/ml:59.56%, 10 μg/
ml:63.64%, 15 μg/ml:65.34%, 20 μg/ml:74.98%로 G1 개체군이 대조군과 비교하여 농도 의존적으로 증가 하는 것을 확인할 수 있었다. 따라서 widdrol은 폐암세포 주기에 영향을 주어 모든 주기를 G1기에 정지시키는 효 과를 나타내는 것으로 사료된다.
3. Widdrol이 세포주기 관련 유전자의 발현에 미치는 영향
Fig. 1, 2의 결과에 의하면 widdrol 처리로 A549 세포의 성장 억제 및 G1 세포주기 정지 효과를 확인할 수 있었 다. 이와 같은 효과는 widdrol에 의해 세포주기 관련 유전 자의 변화를 초래하여 나타나는 결과로 해석할 수 있다.
따라서 DNA microarray 분석을 통해 세포주기 관련 유전 자의 변화를 확인하기로 하였다.
Widdrol 처리군과 비처리군에서 세포주기 관련 유전 자의 발현 변화를 확인한 결과 다양한 세포주기 관련 유
전자의 변화를 관찰할 수 있었다(Table 1). 대조군에 비해 p73은 +1.04, c-abl은 +0.55 배 증가하는 것을 알 수 있었 다. p73은 p53과 비슷한 암 억제유전자로 ATM (ataxia- telangiectasia-mutated), c-abl에 의해 활성화 되고, G1기에 서 S기로 가는 세포주기 과정을 제어한다.24) CDK inhi- bitor로써 CDK-cyclin complex 활성을 감소시키는 p21은 +1.13배 증가 하였다. 세포주기 엔진으로 알려져 있는 cyclin D3와 CDK6도 −0.59, −0.34배 감소하였다. Widdrol 처리에 의해 p21의 증가하고, 이것이 CDK의 활성 저하 로 연결되는 것으로 보인다.
E2F1은 +1.47배 증가하였으며 이 단백질은 과발현 되 면 apoptosis를 유도하는 것으로 알려져 있다. fos와 jun 그 리고 myc는 −0.67, −0.44, −0.53배 감소하였다. c-myc는 암유전자로 이들은 여러 인체 암 조직이나 암 세포주에 서 유전자가 증폭되어 있음이 알려져 있다. c-myc단백질 은 핵에 존재하며, 이중 또는 단일 DNA와 결합하는 능 력을 가지고 있으며 유전자의 전사를 촉진하는 것으로 알려져 있다.25) 전사 조절 인자 중 일부는 성장인자 등의 자극에 의해 유도되는데 c-fos나 c-jun 등이 이에 속한다.26) 이 유전자들이 감소함으로써 전사가 억제되었음을 확인 할 수 있었다. PIAS는 miz라는 단백질로 CDK6을 억제하 는 p15의 단백질을 활성화시킨다.27) 이 유전자의 발현이 +1.08배 증가함으로써 CDK6을 억제하고 RB 단백질을 인산화 시키지 못해 G1기 초기에 전사를 억제함을 추측 할 수 있었다. GADD45A는 p53에 의해 활성화 되는 단백 질로 DNA 복제를 억제한다.28) 이는 +0.67배 증가함으로 써 DNA 복제 제어 효과를 추측해 볼 수 있다. PCNA는
−0.27배 감소하였으며 이는 polymerase 보조인자로 발현 량이 감소함으로써 DNA 복제를 활성화시키지 못함을
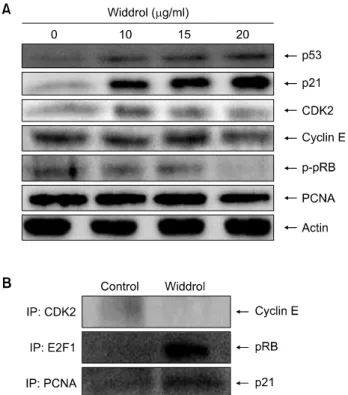
Fig. 3. Altered expression of cell cycle regulatory proteins by widdrol. (A) A549 cells were treated with the indicated concentrations of widdrol for 48 h. The total cell lysates were resolved by SDS-PAGE and the expression levels of p53, p21, CDK2, cyclin E, p-pRB, and PCNA were detected by western blot analysis. Actin was used as a loading control. (B) A549 cells were harvested at the 48 h after incubation with widdrol (20 μg/ml). Total lysates were immunoprecipitated with anti- CDK2, anti-E2F1 and anti-PCNA antibody. Bound cyclin E, pRB and p21 in each immunocomplex were determined by western blot analysis. The immunoreacted proteins were detected using a chemiluminescence system (SuperSignal West Femto Maximum Sensitivity Substrate, Pierce), an the level of reactivity was quantified (FluorChemTM 5500, Alpha Innotech).
확인 할 수 있었다. DNA 이중나선을 꼬거나 풀어주는 topoisomerease는 −0.4배 감소하였다. 이와 같은 DNA microarray 결과를 분석해 보니 widdrol처리에 의해 세포 내에서는 세포주기 정지를 일으키는 움직임이 일어나 며, 또한 DNA 복제를 원활하게 하지 못한다는 사실을 알 수 있었다(Table 1).
DNA microarray의 결과를 정리해 보면 다음과 같다. 세 포주기의 G1기에서 S기로의 진행은 D형 cyclin과 CDK4 또는 6의 complex에 의해 조절되어 진다. 세포주기를 원 활하게 진행시키기 위해서는 세포주기 엔진에 해당하는 D형 cyclin과 CDK4 또는 6의 complex의 활성화가 필요하 며 또한 세포주기가 S기로 진입하면서 DNA 복제에 필 요한 여러 가지 S기 필수 단백질들의 발현 유도가 필요 하게 된다. 하지만 widdrol을 폐암세포 A549에 처리하게 되면 엔진에 해당하는 cyclin D3와 CDK6의 감소와 CDK inhibitor p21의 증가가 나타나게 된다. p21의 증가는 cyclin-CDK 복합체에 결합하여 kinase활성을 저해하여 A549세포가 G1기 정지를 나타내는 것으로 보이며 이와 연동되어 전사인자 E2F 유전자의 변화도 보인다.
E2F (E2 promoter binding factor)는 많은 S기 관련 표적 유전자의 전사인자로 전사를 유도할 뿐만 아니라, pRB, cyclin A, Cdk2 등의 G1기에 있어 중요한 기능을 가진 인 자와 복합체를 형성한다고 알려져 있다. E2F의 전사활성 은 이 전사인자와 복합체를 형성하고 있는 pRB의 인산 화에 의해 유도된다. E2F와 결합하여 전사를 억제하고 있던 pRB가 G1기 후기에서 G1/S기에 걸쳐 cyclin/CDK complex에 의해 인산화 되면서 E2F는 pRB 결합형에서 유 리형 E2F가 되고 proliferating cell nuclear antigen (PCNA), DNA polymerase α, MCM, ORC 등 DNA 복제에 관여하는 많은 유전자의 promoter 영역에 결합하여 전사를 활성화
시킨다.29,30) Table 1의 결과에 의하면 E2F 유전자의 감소
로 인해 복제단백질 PCNA, topoisomerase의 감소도 함께 관찰되는 것으로 사료된다.
4. 세포주기 조절 단백질들의 발현 양상
Widdrol처리에 의해 세포 주기 조절 단백질들의 발현 양상을 검토하기 위해 western blot analysis를 수행하였다.
Fig. 3A에 나타난 결과와 같이 A549 cell내에서 widdrol 처 리에 따라 농도 의존적으로 항암단백질로 잘 알려진 p53 의 발현이 증가하였으며, CDK inhibitor인 p21의 발현도 현저히 증가함을 확인할 수 있었다. 그리고 CDK2와 cyclin E가 감소하였으며 pRB의 인산화 정도를 검토한 p-pRB가 감소함으로써, 이는 DNA microarray의 결과와 동일한 결과로 세포주기의 진행이 원활하지 못하다는
것을 알 수 있었다.
Cyclin E 발현은 cyclin D보다 약간 늦은 G1기부터 증가 하여 G1-S 경계에서 정점이 된다. Cyclin E-CDK2는 S기로 진입하는데 역할을 한다.31) 그러므로 Cyclin E-CDK 2의 kinase 활성 및 발현에 영향을 미치는 어떠한 인자는 정 상적인 pRB의 활성을 막고 G1기에서 S기로의 진입을 방 해하여 세포의 축적을 유도한다.
전체적인 단백질들의 발현 양상의 변화는 확인하였으 나, widdrol처리에 의해 complex의 형성에도 영향을 미치 는지 알아보기 위하여 immunoprecipitation 분석을 행하였 다(Fig. 3B).
Immunoprecipitation 분석에서 widdrol을 A549 세포에
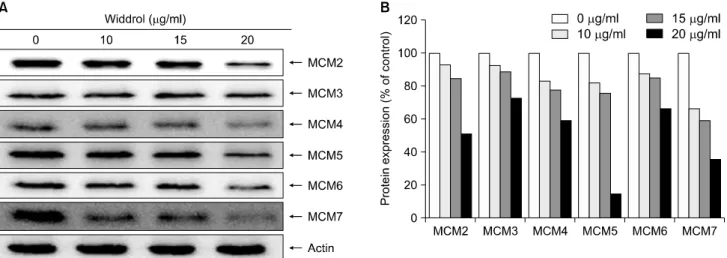
Fig. 4. The inhibitory effect of widdrol on the expression of MCM proteins in A549 cells. The cells were treated with various concentrations of widdrol for 48 h, and (A) the expression of MCM 2, 3, 4, 5, 6, and 7 were detected by western blot analysis.
(B) The protein expression levels were shown as bars from the results of (A). Actin was used as a loading control.
처리하였을 경우 대조군에 비해 CDK2에 결합하는 CDK2/
cyclin E complex가 31% 감소함을 알 수 있었다. 이로써 CDK2/cyclin E complex의 감소로 세포주기를 진행하는 동 력 장치 역할이 제어되어 세포주기 진행이 원활하지 못 함을 알 수 있다. 게다가 E2F1과 pRB의 결합능을 확인하 기 위하여 IP는 E2F1으로 행하고 western blot은 pRB로 확 인한 결과 대조군과 비교하여 widdrol을 처리한 세포에 서 E2F1과 pRB 결합형이 116.5% 증가하였다. 이는 widdrol이 pRB의 인산화 반응을 방해하여 E2F1과 pRB의 결합형의 형태로 유지하고 E2F1의 방출을 저해함을 알 수 있다. 따라서 방출된 E2F1이 줄어들어 인해 S기에 필 요로 하는 복제 단백질들의 전사를 차단시키고 있음을 시사하고 있다. 이와 동시에 widdrol에 의한 A549 세포에 서 PCNA와 p21의 결합능을 확인하기 위하여 immuno- precipitation을 시행한 결과 대조군과 비교하여 widdrol을 처리한 세포에서 PCNA와 p21의 결합형이 30.4% 증가하 였다. 이로써 PCNA와 p21 결합이 증가함으로써 p21이 PCNA을 억제하여 G1기에서 S기로의 진행을 억제하는 데 영향을 주었다고 판단되어진다.
5. Widdrol에 의한 MCM 단백질의 발현 저하 확인
Widdrol 처리로 topoisomerase의 전사량 변화와 PCNA 의 발현량 변화를 확인할 수 있었다. MCM 단백질은 DNA 복제 개시인자로 helicase 활성을 가지고 있으며 복 제의 licensing factor로써 진핵세포 내에 중요한 역할을 담 당하고 있다. 또한 여러 암세포에서 MCM 발현이 정상세 포와 비교해 과발현 되고 있다는 사실이 보고되어, MCM 이 새로운 항암제 개발의 표적 분자로 주목 받고 있다.14)
MCM 단백질 발현량 변화를 관찰하기 위하여 western blot analysis를 시행하여 확인하였다. Widdrol을 A549 세 포에 처리하였을 경우 Fig. 4에서 보는 바와 같이 대조군 과 비교하여 MCM 2, 3, 4, 5, 6, 7의 모든 MCM 단백질이 발현이 농도 의존적으로 감소함을 확인할 수 있었다. 그 중 MCM7의 경우 widdrol 농도 20 μg/ml일 때 64.6% 감 소를 보여 다른 MCM 단백질 보다 현저한 변화를 보였 다. 따라서 widdrol을 A549에 처리하였을 경우 암세포에 서 과발현되는 MCM 단백질 발현을 억제하고 있음을 알 수 있었다.
폐암세포 A549에 widdrol을 처리하면 세포 증식억제 및 세포주기 정지효과를 나타낸다. 이와 같은 효과는 여 러 항암제가 가지는 효능 중에 한가지라고 생각된다. 그 러나 widdrol의 경우 항암단백질로 알려져 있는 p53과 CDK inhibitor p21의 발현량 증가를 동반하면서 현재 새 로운 항암 타겟으로 알려진 MCM 단백질의 발현량을 제 어하고 있다.
MCM 단백질은 대부분의 암세포 및 종양세포에서 발 현량이 증가되어 있으며 MCM단백질의 발현량 제어는 세포의 정상화에 기여할 수 있다. 현재까지 많은 세포주 기 정지 효과를 가진 항암 후보 물질들이 보고되었으나, MCM 단백질의 발현 제어를 함께 유도하는 물질은 widdrol이 첫 보고이다. 항암활성과 MCM 단백질의 발현 저하의 연관성을 확실하게 규명하기 위해서 widdrol 처 리에 따른 p21의 발현 증가와 MCM 단백질의 발현저하 의 보다 구체적인 메카니즘 연구가 필요하며, 세포주기 정지와 checkpoint의 움직임에 대한 추가적인 연구가 필 요할 것으로 사료된다.
결 론
본 연구에서는 Juniperus chinensis로부터 분리되어진 widdrol을 A549 폐암세포에 처리하여 일어나는 여러 가 지 항암활성에 대해 확인하였으며, 새로운 항암 타겟 단 백질로 떠오르는 MCM 복제단백질의 발현저하 활성을 확인하였다.
Widdrol은 IC50 10 μg/ml 농도로 A549 폐암세포에 시 간별로 처리하였을 때 세포성장을 억제 하였고, 농도별 로 처리해 본 결과 농도 의존적으로 세포주기가 G1기로 모이는 것을 확인하였다. 유전자 변화 양상을 DNA Microarray 법으로 확인하고, 단백질 발현 양상을 western blot analysis로 확인해 보면 특이적으로 항암 단백질 p53 과 그 하류 단백질인 CDK inhibitor p21의 발현 증가를 확인할 수 있었다. 이들 단백질의 발현 증가로 인해 cyclin-CDK 복합체의 저하와 이 복합체의 기질이며 전사 인자를 담고 있는 pRB (pocket protein)의 인산화가 저하되 는 것을 확인할 수 있었다. pRB는 전사인자 E2F를 결합 하고 있다가 세포주기가 진행되면서 cyclin-CDK 복합체 에 의해 인산화되면서 E2F를 방출시키고 방출된 E2F는 S기에 필요한 복제단백질 등의 전사인자로 작용하여 전 사 및 발현을 유도하게 된다. 하지만 widdrol을 처리한 세포에서는 이와 같은 진행이 방해 받아 세포주기의 G1 기 정지 및 S기에 필요한 단백질들의 발현 저하가 확인 되었다. 본 연구에서는 widdrol이 S기의 필수단백질로 helicase활성을 가지고 있는 MCM 단백질의 발현을 현저 하게 저하시키고 동시에 암세포 성장억제 등의 항암활 성을 가지고 있는 특이적인 성분으로 다양한 인간 암에 사용가능한 항암제로써의 개발 가능성을 제시하였다.
감사의 글
본 연구는 2008년 동의대학교 교내 연구비(2008AA108) 지원에 의해 수행되었으며, 이에 감사드립니다.
참 고 문 헌
1) Houtgraaf JH, Versmissen J, Van der Giessen WJ. A concise review of DNA damage checkpoints and repair in mammalian cells. Cardiovasc Revasc Med 7, 165-172, 2006.
2) Kaina B. DNA damage-triggered apoptosis: critical role of DNA repair, double-strand breaks, cell proliferation and signaling. Biochem Pharmacol 66, 1547-1554, 2003.
3) Nakanishi M, Shimada M, Niida H. Genetic instability in
cancer cells by impaired cell cycle checkpoints. Cancer Sci 97, 984-989, 2006.
4) Morgan DO. Principles of CDK regulation. Nature 374, 131- 134, 1995.
5) Lees E. Cyclin dependent kinase regulation. Curr Opin Cell Biol 7, 773-780, 1995.
6) Sherr CJ, Roberts JM. Inhibitors of mammalian G1 cyclin- dependent kinases. Genes Dev 9, 1149-1163, 1995.
7) Peter M, Herskowitz I. Joining the complex: cyclin-dependent kinase inhibitory proteins and cell cycle. Cell 79, 181-184, 1994.
8) Quelle DE, Ashmun RA. Hannon GJ, Rehberger PA, Trono D, Richter KH, Walker C, Beach D, Sherr CJ, Serrano M.
Cloning and characterization of murine p16INK4a and p15INK4b genes. Oncogene 11, 635-645, 1995.
9) Rousseau D, Cannella D, Boulaire J, Fitzgerald P, Fotedar A, Fotedar R. Growth inhibition by CDK-cyclin and PCNA binding domains of p21 occurs by distinct mechanisms and is regulated by ubiquitin-proteasome pathway. Oncogene 18, 4313-4325, 1999.
10) Chong JP, Mahbubani HM, Khoo CY, Blow JJ. Purification of MCM-containing complex as a component of the DNA replication licensing system. Nature 375, 418-421, 1995.
11) Kubota Y, Mimura S, Nishimoto S, Takisawa H, Nojima H.
Identification of the yeast MCM3-related protein as a component of xenopus DNA replication licensing factor. Cell 81, 601-609, 1995.
12) Madine MA, Khoo CY, Mills AD, Laskey RA. MCM3 complex required for cell cycle regulation of DNA replication in vertebrate cells. Nature 375, 421-424, 1995.
13) Lei M. The MCM complex: its role in DNA replication and implications for cancer therapy. Curr Cancer Drug Targets 5, 365-380, 2005.
14) Stark GR, Debatisse M, Giulotto E, Wahl GM. Recent progress in understanding mechanisms of mammalian DNA amplification. Cell 57, 901-908, 1989.
15) Forsburg SL. Eukaryotic MCM proteins: beyond replication initiation. Microbiol Mol Bio Rev 68, 109-131, 2004.
16) Brodeur GM, Hogarty MD. Gene amplification in human cancers: biological and clinical significance. In: eds, by Vogelstein B and Kinzler KW, The Genetic Basis of Human Cancer, New York, McGraw Hill, pp 161-172, 1998.
17) Freeman A, Morris LS, Mills AD, Stoeber K, Laskey RA, Williams GH, Coleman N. Minichromosome maintenance proteins as biological markers of dysplasia and malignancy.
Clin Cancer Res 5, 2121-2132, 1999.
18) Hiraiwa A, Fujita M, Nagasaka T, Adachi A, Ohashi M, Ishibashi M. Immunolocalization of hCDC47 protein in normal and neoplastic human tissues and its relation to growth. Int J Cancer 74, 180-184, 1997.
19) Ishimi Y, Okayasu I, Kato C, Kwon HJ, Kimura H, Yamada K, Song SY. Enhanced expression of Mcm proteins in cancer cells derived from uterine cervix. Eur J Biochem 270, 1089-
1101, 2003.
20) Todorov IT, Werness BA, Wang HQ, Buddharaju LN, Todorova PD, Slocum HK, Brooks JS, Huberman JA.
HsMCM2/BM28: a novel proliferation marker for human tumors and normal tissues. Lab Invest 78, 73-78, 1998.
21) Lee CK, Fang JM, Cheng YS. Norditerpenes from Juniperus chinensis. Phytochemistry 39, 391-394, 1995.
22) Yarelis ON, Iraida SS, Isidro GC, Rosario HG. Sesquiterpenes from the wood of Juniperus lucayana. Phytochemistry 68, 2409- 2414, 2007.
23) Nunez YO, Salabarria IS, Collado IG, Hernandez-Galan R.
The antifungal activity of widdrol and its biotransformation by Colletotrichum gloeosporioides (penz.) Penz. & Sacc. and Botrytis cinerea Pers.: Fr. J Agric Food Chem 54, 7517-7521, 2006.
24) Wang JY. Regulation of cell death by the Abl tyrosine kinase.
Oncogene 19, 5643-5650, 2000.
25) Zhang P, Li H, Wu ML, Chen XY, Kong QY, Wang XW, Sun Y, Wen S, Liu J. c-Myc downregulation: a critical molecular event in resveratrol-induced cell cycle arrest and apoptosis of human medulloblastoma cells. J Neurooncol 80, 123-131, 2006.
26) Murakami Y, Satake M, Yamaguchi-Iwai Y, Sakai M,
Muramatsu M, Ito Y. The nuclear protooncogenes c-jun and c-fos as regulators of DNA replication. Proc Natl Acad Sci USA 88, 3947-3951, 1991.
27) Zhao N, Wang J, Cui Y, Guo L, Lu SH. Induction of G1 cell cycle arrest and P15INK4b expression by ECRG1 through interaction with Miz-1. J Cell Biochem 92, 65-76, 2004.
28) Yamasawa K, Nio Y, Dong M, Yamaguchi K, Itakura M.
Clinicopathological significance of abnormalities in Gadd45 expression and its relationship to p53 in human pancreatic cancer. Clin Cancer Res 8, 2563-2469, 2002.
29) Suzuki S, Adachi A, Hiraiwa A, Ohashi M, Ishibashi M, Kiyono T. Cloning and characterization of human MCM7 promoter. Gene 216, 85-91, 1998.
30) Yamaguchi M, Hayashi Y, Matsukage A. Essential role of E2F recognition sites in regulation of the proliferating cell nuclear antigen gene promoter during Drosophila development. J Biol Chem 270, 25159-25165, 1995.
31) Ohtsubo M, Theodoras AM, Schumacher J, Roberts JM, Pagano M. Human cyclin E, a nuclear protein essential for the G1-to-S phase transition. Mol Cell Biol 15, 2612-2624, 1995.