저작자표시-비영리-변경금지 2.0 대한민국 이용자는 아래의 조건을 따르는 경우에 한하여 자유롭게
l 이 저작물을 복제, 배포, 전송, 전시, 공연 및 방송할 수 있습니다. 다음과 같은 조건을 따라야 합니다:
l 귀하는, 이 저작물의 재이용이나 배포의 경우, 이 저작물에 적용된 이용허락조건 을 명확하게 나타내어야 합니다.
l 저작권자로부터 별도의 허가를 받으면 이러한 조건들은 적용되지 않습니다.
저작권법에 따른 이용자의 권리는 위의 내용에 의하여 영향을 받지 않습니다. 이것은 이용허락규약(Legal Code)을 이해하기 쉽게 요약한 것입니다.
Disclaimer
저작자표시. 귀하는 원저작자를 표시하여야 합니다.
비영리. 귀하는 이 저작물을 영리 목적으로 이용할 수 없습니다.
변경금지. 귀하는 이 저작물을 개작, 변형 또는 가공할 수 없습니다.
의학 석사 학위논문
Analysis of Naturally Occurring Mutations in Reverse Transcriptase
Region of Hepatitis B Virus Genotype C2 Polymerase
유전자형 C2 Hepatitis B virus의 역전사효소 부위에서 자연적으로 발생한
유전자 변이 분석
2017년 02 월
서울대학교 대학원 의학과 미생물학전공
김 지 은
A thesis of the Master’s degree
Analysis of Naturally Occurring Mutations in Reverse Transcriptase
Region of Hepatitis B Virus Genotype C2 Polymerase
유전자형 C2 Hepatitis B virus의
역전사효소 부위에서 자연적으로 발생한 유전자 변이 분석
February 2017
The Department of Microbiology and Immunology,
Seoul National University College of Medicine
Ji-Eun Kim
Analysis of Naturally Occurring Mutations in Reverse Transcriptase Region of Hepatitis B Virus Genotype C2 Polymerase
지도교수 국 윤 호
이 논문을 의학 석사 학위논문으로 제출함 2017년 2월
서울대학교 대학원 의학과 미생물학 전공
김 지 은
김지은의 의학석사 학위논문을 인준함 2017년 2월
위 원 장 (인)
부위원장 (인)
위 원 (인)
Analysis of Naturally Occurring Mutations in Reverse Transcriptase Region of Hepatitis B Virus Genotype C2 Polymerase
by Ji-Eun Kim
A thesis submitted to the Department of medicine in partial fulfillment of the requirements for the Degree of
Master of Science in Medicine at Seoul National University College of Medicine
February 2017
Approved by Thesis Committee:
Professor Chairman
Professor Vice chairman
Professor
i
ABSTRACT
Hepatitis B virus (HBV) is a family of hepatotropic DNA viruses, and replicates its DNA genome through an RNA intermediate during the viral life cycle using reverse transcriptase (RT). Mutations within RT probably affect the replication capacity of HBV, which may in turn alter the antigenicity and generation of drug resistance. HBV is classified into eight genotypes, type A- H, according to the differences in the genome sequence. Patients infected with HBV genotype C are associated with more aggressive liver diseases and poorer response to antiviral therapy.
Full-length HBV RT sequences were analyzed from 131 treatment-naïve Korean patients infected with HBV genotype C2. The patients had two distinct clinical statuses: chronic hepatitis (CH) and hepatocellular carcinoma (HCC). To analyze the patterns and the frequencies of mutations in the RT region, a nested polymerase chain reaction (nested-PCR) based sequencing protocol was used.
The mutation frequency was significantly (p<0.05) higher in the HCC patients and the HBV envelope antigen (HBeAg) negative patients. Notably the A-B interdomain region had a significantly (p<0.05) higher mutation frequency than that of the other regions, and it overlaps the A-determinant of the HBV surface antigen (HBsAg) which is the region involved in vaccine escape. The patients with three type of mutations (rt80, rt139, and rt204) were significantly (p<0.05) relative to the HCC and the severe liver disease
ii
progression. In addition, among 71 out of 131 patients who had received the follow-up antiviral treatment, those with the previously unknown mutation (rt55) and the well known YMDD mutation (rt204) significantly (p<0.05) showed no response to the antiviral treatment.
In order to determine how the mutants affected the HBV replication, this study confirmed with in vitro transfection systems, using full-genome HBV constructs with the site-directed mutation (H55Q, L80I, N139K, and M204I), quantifying of HBV DNA and HBsAg levels, and treatment of antiviral agents.
In conclusion, naturally occurring RT mutations might be indicators for the prediction of the liver disease progression and antiviral treatment response of patients with HBV C2 infection.
--- Keywords: genotype C2 HBV infection, reverse transcriptase,
HBV surface antigen, chronic hepatitis, hepatocellular carcinoma, antiviral resistance Student number: 2015-21958
iii
Contents
Abstract ... i
Contents ...iii
List of Tables ... v
List of Figures ... vi
List of Abbreviations ... vii
Introduction ... 1
Materials and Methods ... 4
Results ... 11
Distribution of naturally occurring RT mutations ... 11
The mutation frequency according to liver disease and HBeAg seroconversion... 16
Discovery of naturally occurring HCC specific RT mutations in patients with genotype C2 ... 18
Clinical severity and liver function factors in patients with naturally occurring HCC specific RT mutations ... 23
Correlation with antiviral treatment response in patients with naturally occurring mutations ... 28
Discussion ... 34
iv
References ... 38
Abstract in Korean ... 48
v
LIST OF TABLES
Table 1. Definition of HBV mutations and polymorphism at RT based on database and cohort data. ... 10 Table 2. Mutation distribution according to functional region and frequency of
the HBV RT and the HBsAg ... 15 Table 3. Analysis of mutation number and frequency at 86 patients ... 17 Table 4. Comparison of clinical data between CH and HCC patients ... 20 Table 5. Statistical analysis of naturally occurring HCC specific RT mutations .... 22
vi
LIST OF FIGURES
Figure 1. Distribution of the naturally occurring mutations in the RT regions and HBsAg. ... 13 Figure2. Correlation between chronic hepatitis patients and hepatitis cellular
carcinoma patients according to 10 major mutations. ... 21 Figure 3. Analysis of the platelet level in patients with the rt55, rt80, rt139,
and rt204 mutations ... 24 Figure 4. Analysis of the HBV DNA and HBsAg levels for patients with the
rt55, rt80, rt139, and rt204 mutations ... 25 Figure 5. Naturally occurring HCC specific RT mutations related to the
extracellular HBV DNA and secreted HBsAg in hepatocellular carcinoma cell line. ... 26 Figure 6. Antiviral treatment response according to mutations ... 30 Figure 7. Naturally occurring RT mutations with antiviral treatment failure
increase antiviral resistance in hepatocellular carcinoma cell line. ... 32
vii
LIST OF ABBREVIATIONS
ADV Adefovir
AST Aspartate aminotransferase ALT Alanine aminotransferase CH Chronic hepatitis
ETV Entecavir
FIB-4 Fibrosis-4 HBV Hepatitis B virus HBsAg HBV surface antigen HBeAg HBV envelope antigen HCC Hepatocellular carcinoma
LMV Lamivudine
LdT Telbivudine
NAs Nucleos(t)ide analogues
NAr Nucleos(t)ide analogues resistance MHR Major histocompatibility region
MIF Melanocyte-stimulating hormone inhibiting factor ORF Open reading frames
PCR Polymerase chain reaction
PT Prothrombin time
RT Reverse transcriptase TNF-α Tumor Necrosis Factor-α
1
INTRODUCTION
Hepatitis B virus (HBV) infection is a major health problem worldwide.
The serious consequences of disease include chronic hepatitis (CH), liver cirrhosis (LC) and hepatocellular carcinoma (HCC) [1]. The annual number of deaths caused by HBV-related diseases including LC and HCC estimated to be approximately 786,000 worldwide [2].
HBV is the member of the hepadnaviridae, a family of hepatotropic DNA viruses, and classified into eight Genotypes, type A-H, based on inter-group divergence in the entire genomic nucleotide sequences of 8% or greater [3].
HBV genotypes have a distinct geographical distribution worldwide.
Genotypes A to D are the most common worldwide, with genotype A predominant in western Europe, North America, genotype C prevalence with Asia, genotype D present in Mediterranean countries. It has become increasingly evident that HBV genotypes have an impact on the natural course of chronic HBV infection. Patients infected with HBV genotype A also respond better to interferon-based treatment compared to genotype D infected patients. Furthermore genotype A is associated with earlier HBV envelope antigen (HBeAg) seroconversion compared with genotype C [4]. Numerous studies have shown that genotype C is associated with more aggressive liver disease and poorer response to antiviral therapy compared to genotype B [5].
The sub-genotypes occurred by co-infections and recombination. It is
2
leading to the emergence of hybrid strains that can become the dominant sub- genotype prevailing in certain geographical regions [6]. The sub-genotype C1 is prevalent in Vietnam and Thailand and related to high rates of the HCC.
Sub-genotype C2 in Korea, China, and Japan increases the risk of HCC and cirrhosis in comparison with genotype B. Other C3 in Pacific Islands but liver disease association is not studied [7].
The HBV has an incomplete double-stranded DNA genome that is approximately 3.2 kb in length and contains 4 overlapping open reading frames (ORFs) encoding the polymerase (P), core (C), surface antigen (S), and X protein [8]. The HBV is highly prone to error and lacks proofreading activity, which can lead to HBV mutations that occur at a 10-fold higher frequency compared to other DNA viruses [9]. The HBV replicates its DNA genome through an RNA intermediate during the viral life cycle using reverse transcriptase (RT) [10]. The RT of the HBV polymerase consists of 344 amino acids (aa) and completely overlaps with the HBV surface antigen (HBsAg) (Figure 1A). The HBsAg region of the HBV plays an essential role in the interaction with immune response, encoding several T- and B-cell epitopes [11]. Mutations within RT probably affect the replication capacity of HBV, which may in turn alter the antigenicity, encapsidation and virulence of the virus as well as the generation of drug resistance [12, 13].
Previously, several studies have been found drug resistant mutations in RT during nucleos(t)ide analogues (NAs) therapy such as lamivudine (LMV) [14],
3
adefovir (ADV) [15], entecavir (ETV) [16], telbivudine (LdT) [17] and tenofovir (TNF) [18]. Potential nucleos(t)ide analogues resistance (NAr) mutations are grouped into 4 categories(Ⅰ-Ⅳ) in accordance with clinical antiviral treatment resistance: primary antiviral resistance mutation (categoryⅠ), secondary/compensatory antiviral resistance mutations (categoryⅡ), putative antiviral resistance mutations (categoryⅢ), and pre- treatment antiviral resistance mutations (categoryⅣ) [19]. It has been found that NAr mutations are pre-existed which are not induced by antiviral agents in treatment-naïve patients with chronic hepatitis, from Europe, Asia, and the Middle East, but the prevalence vary from 0% to 57% [20-23] .
The prevalence of infection with HBV genotype C2 in Korea may influence the liver disease progression; based on the Korean National Health and Nutrition Survey for the year 2011, the prevalence of HBsAg positivity was 3.4% among men and 2.6% among women [24, 25]. In addition, several unique types of naturally occurring HBV mutations related to the progression of liver disease (particularly HCC) that are rarely, if ever, encountered in other areas have been found in South Korea [26-30]. The purpose of this study was to analyze naturally occurring HBV RT mutations, and find HCC-specific and antiviral resistance mutations in 131 Korean treatment-naïve patients infected with HBV genotype C2.
4
MATERIALS AND METHODS 1. Treatment-naïve patient samples
Treatment-naïve patient serum samples were collected from the 131 chronic hepatitis patients who visited Konkuk University Hospital, met the inclusion criteria of HBsAg positivity and HBV DNA positivity and were LMV, ADV, ETV, LdT and TNF treatment-naïve. All patients had negative tests for hepatitis C virus, human immune-deficiency virus and markers for co-existing autoimmune liver disease.
The Institutional Review Boards (IRB) of Seoul National University Hospital (IRB-1605-065-761) and the Konkuk University Hospital (KUH- 1010544) approved this study. The experiments based on extracted virion DNA from isolates; hence, the study did not require informed consent and the waiver of informed consent agreed upon by the IRBs.
2. HBV DNA extraction and PCR amplification
HBV DNA was extracted from the secured 200㎕ of serum samples using the QIAamp DNA Blood Mini Kit (QIAGEN Inc, Hilden, Germany). To analyze the mutation patterns and the frequencies of mutations in the RT region, a nested Polymerase chain reaction (PCR) based sequencing protocol was used as previously described [31]. The first-round PCR was performed using the sense primer Pol-RT-F1 (5'-CAG CCT ACT CCC ATC TCT CCA CCT CTA AG -3') and the antisense primer Pol-RT-R1 (5'-GCT CCA GAC
5
CGG CTG CGA GC -3') to yield a 1,375-bp amplicon between nucleotide 3,157 and 1,316 of the HBV genome. The second-round PCR was performed using the sense primer Pol-RT-F2 (5'-CCT CAG GCC ATG CAG TGG AA -3') and the antisense primer Pol-RT-R2 (5'-GTA TGG ATC GGC AGA GGA GC- 3') to yield a 1,291-bp amplicon between nucleotide 3,196 and 1,271 of the HBV genome. The PCR was initiated in a 20㎕ PCR mixture containing 1.5 mM MgCl2, 250μM dNTPs and 1.0 U of the ProFi Taq DNA polymerase (Bioneer). For both rounds, the protocol was as follows: 95 °C for 10 min, followed by 15 cycles at 94 °C (15 sec), 55 °C (15 sec) and 68 °C (3 min).
Final extension step was 72 °C for 5 min. The second-round PCR protocol used 2㎕ of the product from the first-round PCR and was identical to the conditions described above except that 30 cycles were performed. The PCR products analyzed by electrophoresis on 1.0% agarose gels, stained with ethidium bromide, and visualized on a UV transilluminator.
3. HBV genotyping
For genotyping, a phylogenetic analysis based on entire sequences of the RT region (1,032 bp) was PCR-positive 131 HBV strains. The 1,032-bp RT sequences of the 131 HBV strains were compared with the sequences of eight reference strains representing each of the genotypes (A-F including the C2 strains) obtained from GenBank [accession numbers M57663 (A), AB100695 (B), AB074755 (C1), AY247032 (C2), AY641559 (C2), X02496 (D),
6
AB106564 (E), and X75663 (F)]. Phylogenetic trees were inferred using the neighbor-joining method in MEGA version 7.0.14 [32]. The phylogenetic analysis based on 1,032-bp sequences covering the entire RT region showed that the 131 HBV strains were genotype C2. The sequences used for mutation analysis were registered at GenBank. [CH patients (GenBank Nos: KX264864 -KX 264922) and HCC patients (GenBank Nos: KX264792-KX264863)]
4. Mutation analysis and definitions
The aa mutations were identified by comparing the HBV RT sequences obtained in this study and the Stanford HBV RT database [33]. The mutation type was defined as change in a sequence deviating from normal, and polymorphism type is the DNA sequence variation that common in population [34].
According to the Stanford database (Table 1), the genotypes divided into eight types (A-H) and each position has the consensus types (common type at each genotype) and the mutation types. However, the consensus types were not existed only one type, variable with genotype. These defined as polymorphism and it is not included mutation. Along with rt269, rt333, rt337 position had additional polymorphism in this study.
5. Statistical analyses
Differences between categorical variables analyzed using Fisher’s exact test
7
or the Chi-square test. For continuous variables, Student’s t-test and the Mann-Whitney U test was used. The multiple tests via Bonferroni correction adjusted level of significance of each test. The SPSS version 18.0 software (Professional Statistic, Chicago, IL, USA). Statistical consultation from the Medical Research Collaborating Center at the Seoul National University Hospital and the Seoul National University College of Medicine
6. Site-directed mutagenesis
The HBV full-length genome construct with rt55, rt80, rt139, and rt204 mutation was generated by site-directed mutagenesis of the wild type pHBV- 1.2x vector which was provided by Prof. Jung G, et al [35]. The mutagenesis was performed using the primers: H55Q (F: TCT AGG GGG AGC ACC CAA GTG TCC TGG CCA AAA, R: TTT TGG CCA GGA CAC TTG GGT GCT CCC CCT AGA), L80I (F: TCC AAT TTG TCC TGG ATA TCG CTG GAT GTG TCT, R: AGA CAC ATC CAG CGA TAT CCA GGA CAA ATT GGA), N139K (GAT TCC TGC TCA AGG AAA CTC TAT GTT TCC CTC, R: GAG GGA AAC ATA GAG TTT CCT TGA GCA GGA ATC) and M204I (F: TTG GCT TTC AGT TAT ATT GAT GAT GTG GTA TTG, R: CAA TAC CAC ATC ATC AAT ATA ACT GAA AGC CAA). These constructs with site- directed mutation (H55Q, L80I, N139K, and M204I) used for in vitro systems.
8
7. Cell culture and transfection
The HepG2 cells were grown in Dulbecco’s modified Eagle’s medium (Welgene, South Korea) supplemented with 10% fetal bovine serum (FBS) and 100ug/ml of penicillin-streptomycin (Welgene, South Korea) at 37℃ in a humidified environment containing 5% CO2. For transient transfection, HepG2 cells seeded into 60 mm dishes and allowed to adhere overnight. On the following day, when the cells were 60-80% confluent, transiently transfected using lipofectamine and harvested 24hr, 48hr, 72hr. Antiviral agents, entecavir (ETV, 30µM) and lamivudine (LAM, 500µM) were treated with 5 days with media.
8. Quantitative HBV DNA replication and HBsAg ELISA assay
Precipitation of HBV DNA in cell culture supernatant with 50% PEG solution conducted using the viral gene-spin kit (Intron, South Korea). HBV DNA was measured using a quantitative real-time PCR (Q-PCR) targeting the HBV DNA. The PCR amplification was performed with a set of real-time PCR primers targeting the small S gene designed to amplifying with primer sequences as follows: sense primer Real-SF (TTG ACA AGA ATC CTC ACA ATA CC) and antisense primer Real-SR (GGA GGT TGG GGA CTG CGA AT). The quantitative PCR assay was conducted using the commercial Sensi FAST SYBR Lo-ROX kit (BIOLINE, London, UK) and primers
9
specific to the S gene with an ABI-7500 system (Applied Biosystems, CA, USA). To compare the HBsAg secretion between wild type and mutant type at the supernatant, quantification of the HBsAg using the HBsAg ELISA kit (BIOKIT. Barcelona, Spain).
10
For example, at the rt7 position, the consensus type is D, and the mutation type is A, V, and T in genotype A; however, the consensus type is T, and the mutation type is A and I in genotype C. Therefore, for the various genotypes, T, Y, A, and D are the polymorphisms at the rt7 position.
Table1. Definition of HBV mutations and polymorphisms in RT based on database and cohort data.
11
RESULTS
Distribution of naturally occurring RT mutations
Overall, 10 major mutations (rt40, rt55, rt80, rt128, rt134, rt139, rt191, rt204, rt325, and rt332) were detected, and among them, 6 were potential NAr mutations which are grouped into 4 categories (Ⅰ-Ⅳ). The detected NAr mutations were rt204 (6.8%, Ⅰ), rt80 (3.8%, Ⅱ), rt128 (12.2%, Ⅲ), rt191 (3.8%, Ⅲ), rt134 (7.6%, Ⅳ), and rt139 (3.8%, Ⅳ). The remaining mutations at rt40 (4.5%), rt55 (7.6%), rt325 (4.5%), and rt332 (5.4%) were not included in the NAr 4 categories (Figure 1B). Additionally, the 4 major mutations detected in HBsAg region (overlapped with RT region), the s126 (6.8%, rt134, Ⅳ) and s196 (6.1%, rt204, Ⅰ) mutations were affected by the RT 10 major mutations, but the s47 (7.6%) and s160 (8.3%) mutations were not affected (Figure 1C).
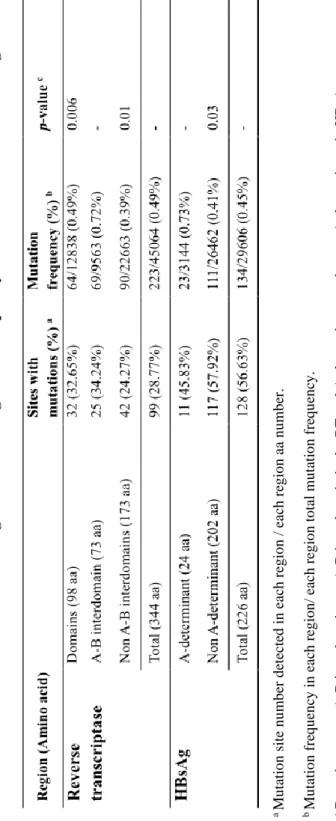
The HBV RT region consists of 7 functional domains (G, F, A, B, C, D and E) and 6 interdomains (G-F, F–A, A–B, B–C, C–D, and D–E). The distribution analysis of naturally occurring RT mutations revealed that there were 99 positions with 223 mutation frequency; especially, the mutation frequency of the A–B interdomain (0.72%) was significantly higher than those of the domain regions (0.49%, p=0.006) and non A-B interdomain regions (0.39%, p=0.01, Table 2). The RT region includes the complete HBsAg region. Thus, RT mutations, particularly in the A-B interdomain of
12
the RT section that overlaps the A-determinant of HBsAg, can result in HBsAg mutations and subsequently, vaccine escape mutants. This A- determinant region (0.73%) had a significantly higher mutation frequency compared to the non A-determinant regions (0.41%, p=0.03, Table 2).
13
14
Figure 1. Distribution of the naturally occurring mutations in the RT regions and HBsAg.
(A) HBV RT includes HBsAg, and it overlaps from aa position 9-234 in RT, and shifted downstream by 1 nucleotide.
(B) The mutations in this study identified for the full-length sequences of HBV RT from 131 patients. Potential NAr mutations are grouped into 4 categories:
primary drug resistance mutations (categoryⅠ), secondary/ compensatory mutations (category Ⅱ), putative antiviral resistance mutations (category Ⅲ), and pre-treatment mutations (category Ⅳ).
(C) The mutations of MHR (100-160 aa) and the A-determinant (124-147 aa) in the HBsAg region (1-226 aa).
15
a Mutation site number detected in each region / each region aa number. bMutation frequency in each region/ each region total mutation frequency. cDomains or non A-B interdomains vs A-B interdomain in the RT and A-determinant vs the non A-determinant in HBsAg.
Table 2. Mutation distribution according to functional region and frequency of the HBV RT and the HBsAg.
16
The mutation frequency according to liver disease and HBeAg seroconversion
With the sequence analysis data of the full-length HBV RT from 131 treatment-naïve Korean patients, this study identified positions of 59 polymorphisms and 99 positions in the RT region. The analysis showed that the mutations were present in 86 of the 131 patients (65.64%), and among them 39 (66.10%) and 47 (65.27%) patients belonged to the CH and HCC groups, respectively. The mutation frequency in the HCC patients (0.58%) was significantly higher than in the CH patients (0.38%, p=0.002, Table 3).
Additionally, 37 (61.66%) and 49 (69.01%) patients were HBeAg negative and positive, respectively, and the HBeAg negative group had a significantly higher mutation frequency (0.56%) than that of the HBeAg positive group (0.43%, p=0.04, Table 3).
17
Mutation number and frequency were divided with liver disease progression and HBeAg.
Table3.Analysis of mutation number and frequency at 86 patients.
18
Discovery of naturally occurring HCC specific RT mutations in patients with genotype C2
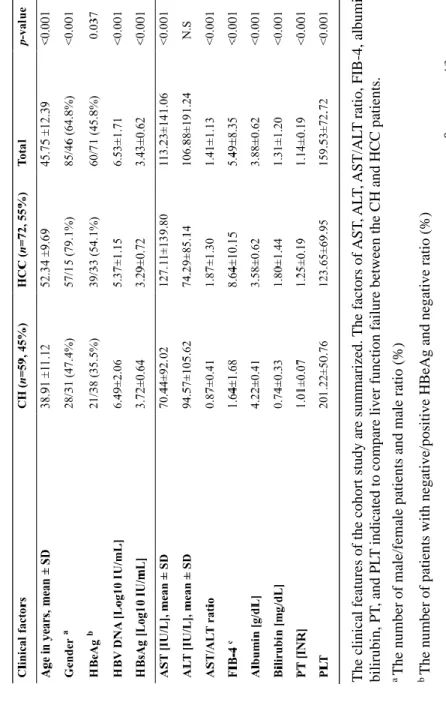
The patient cohort consisted of CH (n=59, 45%) and HCC (n=72, 55%) patients which included 85 males (64.8%) and 46 females (35.1%) with a median age of 45 years (range 22–73 years). The main characteristics of the CH and HCC patients compared in Table 4. The HCC patients were significantly older (p<0.001) and had a significantly higher number of male patients (p<0.001) and HBeAg-negative patients (p=0.037). Additionally, the HBV DNA and HBsAg levels in the HCC patients were significantly lower compared to the CH patients (p<0.001, p<0.001).
The risk factors of the liver function failure: aspartate aminotransferase (AST), alanine aminotransferase (ALT), AST/ALT ratio, fibrosis (FIB-4), albumin, bilirubin, prothrombin time (PT), and platelets (PLT) were compared using the patients clinical data. This cohort did not have any difference in ALT; however, the AST (70.44: 127.11, p<0.001), AST/ALT ratio (0.87: 1.87, p<0.001), FIB-4 (1.64: 8.64, p<0.001), bilirubin (0.74: 1.80, p<0.001), PT
(1.01: 1.25, p<0.001) were significantly higher in the HCC group.
Additionally, albumin (4.22: 3.58, p<0.001) and PLT (201.22: 123.65, p<0.001) were lower in the HCC group.
Overall, 10 major naturally occurring mutation sites, among them, 4 sites, rt55, rt80, rt139, and rt204 mostly detected in the HCC patients. Moreover, the rt80, rt139, and rt204 mutations were significant in the HCC patients
19
(p=0.036, p=0.036, p=0.004); however, the rt55 mutations were not significantly higher in HCC patients (Figure 2).
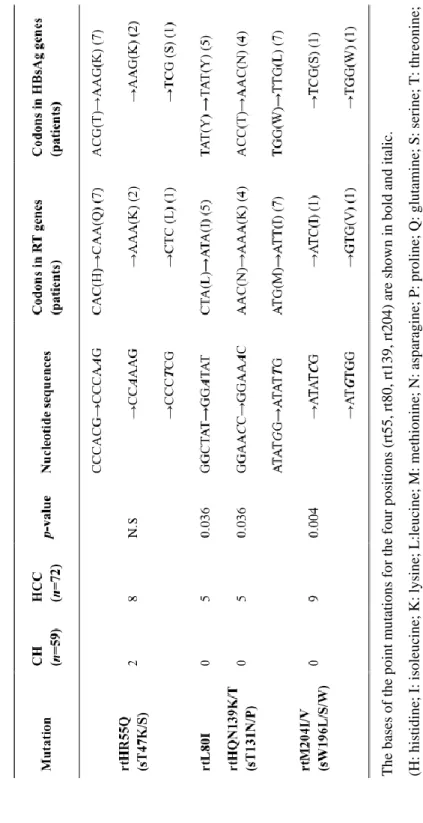
These mutations except for rt80 led to simultaneous HCC related mutations at s47, s131, and s196 in HBsAg, which overlaps the RT region (Table 5).
The only type mutation, L80I, was found in the HCC patients (n=5, p=0.036) and not in the CH patients. Of note, the 5 HCC patients with the L80I mutation also had a M204I mutation. The second mutation types, N139K (n=4) and N139T (n=1) at the rt139 mutation site were found in 5 HCC patients but not in the CH patients (p=0.036), which also led to simultaneous T131N (n=4) and T131P (n=1) HBsAg mutations in the 5 HCC patients (Table 5). For the third mutation type, M204I (n=8) and M204V (n=1), at rt204 leading to YMDD motif mutations were found in 9 HCC patients (p=0.004), which also led to simultaneous W196L (n=7) and W196S (n=1) HBsAg mutations in 8 HCC patients (Table 5).
20
The clinical features of the cohort study are summarized. The factors of AST, ALT, AST/ALT ratio, FIB-4, albumin, bilirubin, PT, and PLT indicated to compare liver function failure between the CH and HCC patients. a The number of male/female patients and male ratio (%) bThe number of patients with negative/positive HBeAg and negative ratio (%) c FIB-4 is calculated using Sterling’s formula, as follows; age[years]× AST[IU/L] /PLT[10⁹/L]× ALT1/2[U/L].
Table 4. Comparison of clinical data between CH and HCC patients.
21
Figure2. Correlation between chronichepatitispatientsandhepatitis cellular carcinoma patients according to 10 major mutations. Of the 10 identified major mutations, mutations at three sites, rt80, rt139, and rt204, only found in the HCC patients and not in the CH patients.
22
The bases of the point mutations for the four positions (rt55, rt80, rt139, rt204) are shown in bold and italic. (H: histidine; I: isoleucine; K: lysine; L:leucine; M: methionine; N: asparagine; P: proline; Q: glutamine; S: serine; T: threonine; V: valine; W: tryptophan; Y: tyrosine).
Table 5. Statistical analysis of naturally occurring HCC specific RT mutations.
23
Clinical severity and liver function factors in patients with naturally occurring HCC specific RT mutations
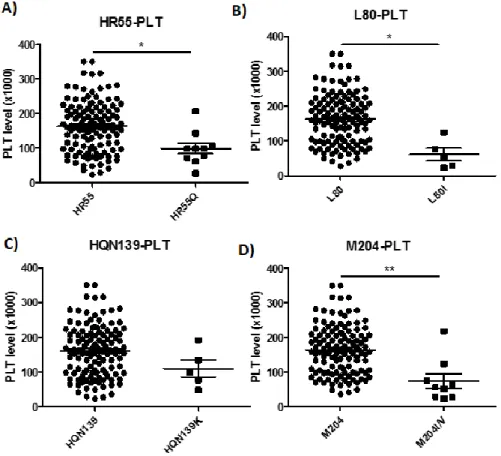
To identify any correlations between clinical factors and naturally occurring mutations, this cohort study investigated with age, gender, HBeAg, HBV DNA, AST, ALT, albumin, bilirubin, PT, and PLT and then antiviral drug treatment. The patients with the naturally occurring rt55, rt80, rt139, and rt204 mutants had a lower PLT level. The PLT level was low in patients with the rt55 (p=0.029), rt80 (p=0.011), rt139 (NS), and rt204 (p=0.002) mutants (Figure 3).
In addition, the patients with the naturally occurring rt55, rt80, rt139, and rt204 mutants had a lower HBV DNA level and HBsAg level than without mutants (NS, Figure 4A, 4B).
To determine if these mutations related to HBV DNA replication and HBsAg, using an in vitro transfection system. The rt55 (H55Q), rt80 (L80I), rt139 (N139K), and rt204 (M204I) mutants had significantly (p<0.005) lower extracellular HBV DNA replication compared to the wild type (Figure 5A). In addition, the intracellular HBV DNA and the secreted HBsAg level were relatively lower for the mutants compared to the wild type (Figure 5B, 5C).
24
Figure 3. Analysis of the platelet level in patients with the rt55, rt80, rt139, and rt204 mutations.
(A) The patients were divided into without (n=121) and with (n=10) rt55 mutation, and the with rt55 mutation group had a significantly lower platelet level (p=0.029).
(B) The patients with rt80 mutation (n=5) had a significantly lower platelet level (p=0.011).
(C) The patients with rt139 mutation (n=5) had a lower platelet level (NS)
(D) The patients with rt204 mutation (n=9) had a significantly lower PLT level (p=0.002).
25
Figure 4. Analysis of the HBV DNA and HBsAg levels for patients with the rt55, rt80, rt139, and rt204 mutations.
(A) The patients with the rt55, rt80, rt139, and rt204 mutations had a relatively lower HBV DNA level than without mutations (NS).
(B) The same patients also had a relatively lower HBsAg level than the patients without mutations (NS).
26
27
Figure 5. Naturally occurring HCC specific RT mutations related to the extracellular and intracellular HBV DNA and secreted HBsAg in hepatocellular carcinoma cell line.
HepG2 cells were transiently transfected with mutants (H55R, L80I, N139K, M204I, and L80I+M204I), and the extracellular and intracellular HBV DNA and secreted HBsAg level were quantified at 24, 48, and 72 hr.
(A) Mutants had a significantly (p<0.005) lower extracellular HBV DNA replication level than the wild type. The double mutation (L80I+M204I) is a complementary mutation, which restores HBV replication; it could be as an in vitro control for comparison with other suppressed HBV DNA mutants.
(B) The mutants had a relatively lower (NS) intracellular HBV DNA replication than that of the wild type.
(C) The mutants had a relatively lower (NS) secreted HBsAg level than that of the wild type.
28
Correlation with antiviral treatment response in patients with naturally occurring mutations
To identify naturally occurring NAr mutations, the antiviral treatment response reported retrospectively from the patients with HBV genotype C2 infections. The 71 patients were investigated for an antiviral treatment, and of these, antiviral treatment failure occurred in 21patiens (30%). The rt55 (n=5, p=0.007), rt80 (n=3, NS), and rt204 (n=6, p<0.001) mutants were frequently
found in patients with antiviral treatment failure.
The antiviral treatment investigation was done for 6 of the 10 patients who had the rt55 mutant, and 5 of them showed antiviral treatment failure. The antiviral treatment failure ratio was compared between the groups without (24.61%) and with (83.33%) mutants. It was significantly higher in the mutant group (p=0.007, Figure 6A). The antiviral treatment investigation was done for 3 out of 5 who had the rt80 mutants, and all of 3 patients showed antiviral treatment failure but not significantly (NS, Figure 6B). In addition to, the antiviral treatment investigation was done for 6 of the 9 patients who had the rt204 mutant, and all of 6 patients showed antiviral treatment failure. The antiviral treatment failure ratio was higher compared between the groups without (23.07%) and with (100%) mutants, and it was higher than in the mutant group (p<0.001, Figure 6C).
The antiviral treatment responses confirmed with HepG2 transfection system in vitro and treatment of antiviral agents (ETV and LAM). These antiviral
29
agents effectively reduced extracellular HBV DNA replication in the wild type; however, the construct with the rt55 (H55Q), rt204 (M204I), and rt80+rt204 (L80I+M204I) mutation were not suppressed, and even worse, extracellular HBV DNA replication significantly increased despite antiviral treatments (p<0.005, Figure 7A). The antiviral treatment in the mutant group could not suppressed the intracellular HBV DNA and HBsAg (Figure 7B, 7C).
30
31
Figure 6. Antiviral treatment response according to mutation.
(A) Overall, 71 patients received antiviral treatment, and the patients without rt55 mutation had an antiviral treatment failure of 24.61% (16/65); however, most of the patients with the rt55 mutation showed antiviral treatment failure (83%, 5/6, p=0.007).
(B) Patients without rt80 mutation had an antiviral treatment ratio of 26.47% (18/68);
however, all of the 3 patients with the rt80 mutation showed antiviral treatment failure (p=NS).
(C) Patients without rt204 mutation had an antiviral treatment ratio of 23.07%
(15/65); however, all of the 6 patients with the rt204 mutation showed antiviral treatment failure (p<0.001).
32
33
Figure 7. Naturally occurring RT mutations with antiviral treatment failure increase antiviral resistance in the hepatocellular carcinoma cell line.
HepG2 cells were transiently transfected with mutants (H55R, L80I, N139K, M204I, and L80I+M204I) and treated with antiviral agents (ETV and LAM) for 5 days.
(A) Treatment of the ETV and LAM suppressed extracellular HBV DNA replication in the wild type; however, the mutants did not show a decrease, even worse, the levels increased. Especially, the H55Q, M204I, and L80I+M204I mutants showed significantly (p<0.005) increased HBV DNA replication compared to the ones not treated with the antiviral agents.
(B) For quantitative intracellular HBV DNA replication, the antiviral agents suppressed the HBV DNA in the wild type strain; however, it increased in the mutants despite the ETV and LAM treatments.
(C) The ETV and LAM treatment not suppressed the secreted HBsAg in the H55Q, M204I, and L80I+M204I mutants.
34
DISCUSSION
Naturally occurring RT mutations associated with HBV drug resistance have been reported in treatment-naïve chronic patients from several countries [23, 36-38]. In South Korea, higher mutation rates and unique mutation patterns related to clinical implications in several HBV ORFs (Polymerase, HBsAg, X, and pre C/C regions) compared to other countries have been reported to date, and these mutations promote liver disease progression [39]. Furthermore, higher relapse rates after antiviral therapy in chronic Korean patients have also been reported [40]. However, there are no reports regarding the distribution of HCC specific and antiviral resistance mutations from treatment-naïve Korean patients to date. This study analyzed potential HCC specific and antiviral resistance mutations from 131 treatment-naïve Korean patients with HBV genotype C2 infections using direct sequencing protocols.
The results of this study offer several implications. First, this cohort showed the prevalence of patients with mutations was 65.64% (86/131), which was around two times higher than the prevalence of mutants (59/192, 30.73%) in a treatment-naïve Chinese cohort using the same direct sequencing protocols [39]. In addition to, this study had significantly (p<0.005) higher mutation frequency in the HCC patients (0.58%) than the CH patients (0.38%).
Mutations frequently found in the A-B interdomain overlapping the A- determinant region of HBsAg (0.73%) compared to the other regions (0.41%).
This result was in agreement with a previous report that potential NAr
35
positions within this region might be hotspots for naturally occurring mutations in this treatment-naïve population [39] and naturally occurring RT mutations can affect HBsAg secretion and lead to vaccine escape mutants [41]
Second, naturally occurring HCC specific RT mutations (rt80, rt139, and rt204) seemed to be an indicator of liver severity and progression (hepatocarcinogenesis) in Korean patients with genotype C2. The clinical data of the patients with these mutations showed significantly (p<0.005) lower PLT levels,and it indicates that decreased liver function, severe liver damage, and cirrhosis progression [42]. In addition, the patients who had these mutants had relatively lower (NS) HBV DNA replication as well as lower (NS) HBsAg level. It is suggested that these mutations (rt80, rt139, and rt204) may contribute to HCC by possibly suppressing HBV replication and inducing immune escape and persistence infections [43]. Third, after tracing the antiviral treatment response, potential NAr mutations at rt55 and rt204 detected in patients. The patients with the rt55, rt204 mutants had significantly (p<0.005) higher antiviral treatment failure, and these mutants induced extracellular HBV DNA replication despite the antiviral treatment in the in vitro systems. These findings may partially provide a likely explanation for why relapse after antiviral therapy is so prevalent in Korean patients and suggest that these patients should be treated with acyclic phosphonates, such as tenofovir (TDF), which is very potent and has a high genetic barrier to antiviral response [44].
36
Furthermore, the YMDD-motif mutation (M204I/V) naturally occurred in chronic HBV patients without antiviral treatment, such as lamivudine therapy, reported in several studies [45, 46] and L80I first reported as a mutation associated with LMV resistance [47]. They found that L80I and M204I mutants suppressed HBV DNA replication compared to the wild type;
however, the L80I+M204I mutation restored viral loads in the case of the LMV treatment [47]. In this study, 5 patients who had L80I combined with the M204I/V mutation suggested that L80I might have a role in compensating for the defective replication of M204I/V.
It is uncertain if the HBV mutation occurs naturally, or if it occurs due to the infection of a mutated HBV strain. There are three possibilities of the mutation occurrence. First, in horizontal infection, a mutated HBV strain could be transmitted to another individual and another infection route during the perinatal period, persistence infection influenced by the HBV status of the mother [48]. Second, HBV YMDD mutation frequently naturally occur in liver disease patients from China, Korea, Japan and other countries. This distribution is affected by infection with mutated HBV strain and immune pressure from the host system [20]. Especially, distinct immune pressure at the T cell level in Korean patients may also lead to a high mutation frequency [49]. Third, most of HBV mutations occurred after antiviral treatment. Some patients had naturally occurring YMDD mutations before treatment; however, all the patients had YMDD mutation after LAM treatment [50]. This cohort
37
study included the naturally occurring factors (mutated HBV strain infection and host immune pressure), but excluded the environmental factors (antiviral treatment).
In conclusion, this study showed the distribution of naturally occurring genotype C2 HBV RT mutations, and found potential HCC specific and antiviral resistance mutations. The naturally occurring mutations at rt80, rt139, and rt204 may contribute to liver disease progression (particularly HCC generation) and rt55 and rt204 may enhance antiviral treatment failure in chronic Korean patients with genotype C2 infections. It could be a likely explanation for why patients with advanced liver disease are difficult to treat with NAs. Of note, this study has found, for the first time, the importance of the rt55 mutation in antiviral treatment failure.
38
References
1. Beasley RP, Hwang LY, Lin CC, Chien CS. Hepatocellular carcinoma and hepatitis B virus. A prospective study of 22 707 men in Taiwan. Lancet (London, England). 1981;2(8256):1129-33. PubMed PMID: 6118576.
2. Black RE, Cousens S, Johnson HL, Lawn JE, Rudan I, Bassani DG, et al.
Global, regional, and national causes of child mortality in 2008: a systematic analysis. Lancet (London, England). 2010;375(9730):1969-87. doi:
10.1016/S0140-6736(10)60549-1. PubMed PMID: 20466419.
3. Kramvis A, Arakawa K, Yu MC, Nogueira R, Stram DO, Kew MC.
Relationship of serological subtype, basic core promoter and precore mutations to genotypes/subgenotypes of hepatitis B virus. Journal of medical virology. 2008;80(1):27-46. doi: 10.1002/jmv.21049. PubMed PMID:
18041043.
4. Chu CJ, Hussain M, Lok AS. Hepatitis B virus genotype B is associated with earlier HBeAg seroconversion compared with hepatitis B virus genotype C. Gastroenterology. 2002;122(7):1756-62. PubMed PMID:
12055581.
5. Chan HL, Wong ML, Hui AY, Hung LC, Chan FK, Sung JJ. Hepatitis B virus genotype C takes a more aggressive disease course than hepatitis B virus genotype B in hepatitis B e antigen-positive patients. Journal of clinical microbiology. 2003;41(3):1277-9. Epub 2003/03/08. PubMed PMID:
12624067; PubMed Central PMCID: PMCPMC150268.
39
6. Jayalakshmi MK, Kalyanaraman N, Pitchappan R. Hepatitis B Virus Genetic Diversity: Disease Pathogenesis2013 2013-02-27.
7. McMahon BJ. The influence of hepatitis B virus genotype and subgenotype on the natural history of chronic hepatitis B. Hepatology international.
2009;3(2):334-42. doi: 10.1007/s12072-008-9112-z. PubMed PMID:
19669359; PubMed Central PMCID: PMCPMC2716762.
8. Liang TJ. Hepatitis B: the virus and disease. Hepatology (Baltimore, Md).
2009;49(5 Suppl):S13-21. Epub 2009/04/29. doi: 10.1002/hep.22881.
PubMed PMID: 19399811; PubMed Central PMCID: PMCPMC2809016.
9. Nowak MA, Bonhoeffer S, Hill AM, Boehme R, Thomas HC, McDade H.
Viral dynamics in hepatitis B virus infection. Proceedings of the National Academy of Sciences of the United States of America. 1996;93(9):4398-402.
Epub 1996/04/30. PubMed PMID: 8633078; PubMed Central PMCID:
PMCPMC39549.
10. Nassal M. Hepatitis B viruses: reverse transcription a different way. Virus research. 2008;134(1-2):235-49. Epub 2008/03/15. doi: 10.1016/j.virusres.
2007.12.024. PubMed PMID: 18339439.
11. Park J, Zellers ET. Determination of solvents permeating through chemical protective clothing with a microsensor array. J Environ Monit.
2000;2(4):300-6. PubMed PMID: 11249783.
12. Sheldon J, Rodes B, Zoulim F, Bartholomeusz A, Soriano V. Mutations affecting the replication capacity of the hepatitis B virus. Journal of viral
40
hepatitis. 2006;13(7):427-34. Epub 2006/06/24. doi: 10.1111/j.1365- 2893.2005.00713.x. PubMed PMID: 16792535.
13. Torresi J. The virological and clinical significance of mutations in the overlapping envelope and polymerase genes of hepatitis B virus. Journal of clinical virology : the official publication of the Pan American Society for Clinical Virology. 2002;25(2):97-106. Epub 2002/10/09. PubMed PMID:
12367644.
14. Selabe SG, Lukhwareni A, Song E, Leeuw YG, Burnett RJ, Mphahlele MJ.
Mutations associated with lamivudine-resistance in therapy-naive hepatitis B virus (HBV) infected patients with and without HIV co-infection:
implications for antiretroviral therapy in HBV and HIV co-infected South African patients. Journal of medical virology. 2007;79(11):1650-4. Epub 2007/09/15. doi: 10.1002/jmv.20974. PubMed PMID: 17854040.
15. Rodriguez C, Chevaliez S, Bensadoun P, Pawlotsky JM. Characterization of the dynamics of hepatitis B virus resistance to adefovir by ultra-deep pyrosequencing. Hepatology (Baltimore, Md). 2013;58(3):890-901. Epub 2013/03/19. doi: 10.1002/hep.26383. PubMed PMID: 23505208.
16. Tenney DJ, Rose RE, Baldick CJ, Pokornowski KA, Eggers BJ, Fang J, et al. Long-term monitoring shows hepatitis B virus resistance to entecavir in nucleoside-naive patients is rare through 5 years of therapy. Hepatology (Baltimore, Md). 2009;49(5):1503-14. Epub 2009/03/13. doi:
10.1002/hep.22841. PubMed PMID: 19280622.
41
17. Zhang Y, Lian JQ, Li Y, Wang JP, Huang CX, Bai XF, et al. Telbivudine plus adefovir therapy for chronic hepatitis B patients with virological breakthrough or genotypic resistance to telbivudine. European journal of gastroenterology & hepatology. 2013;25(7):814-9. Epub 2013/02/15. doi:
10.1097/MEG.0b013e32835ee516. PubMed PMID: 23406845.
18. Liu Y, Miller MD, Kitrinos KM. HBV clinical isolates expressing adefovir resistance mutations show similar tenofovir susceptibilities across genotypes B, C and D. Liver international : official journal of the International Association for the Study of the Liver. 2014;34(7):1025-32. doi:
10.1111/liv.12343. PubMed PMID: 24118725.
19. Ciftci S, Keskin F, Cakiris A, Akyuz F, Pinarbasi B, Abaci N, et al.
Analysis of potential antiviral resistance mutation profiles within the HBV reverse transcriptase in untreated chronic hepatitis B patients using an ultra- deep pyrosequencing method. Diagn Microbiol Infect Dis. 2014;79(1):25-30.
doi: 10.1016/j.diagmicrobio.2014.01.005. PubMed PMID: 24630522.
20. Tan Y, Ding K, Su J, Trinh X, Peng Z, Gong Y, et al. The naturally occurring YMDD mutation among patients chronically infected HBV and untreated with lamivudine: a systematic review and meta-analysis. PLoS One. 2012;7(3):e32789. doi: 10.1371/journal.pone.0032789. PubMed PMID:
22479339; PubMed Central PMCID: PMCPMC3314000.
21. Tan YW, Ge GH, Zhao W, Gan JH, Zhao Y, Niu ZL, et al. YMDD motif mutations in chronic hepatitis B antiviral treatment naive patients: a multi-
42
center study. Braz J Infect Dis. 2012;16(3):250-5. PubMed PMID: 22729192.
22. Mahabadi M, Norouzi M, Alavian SM, Samimirad K, Azad TM, Saberfar E, et al. Drug-related mutational patterns in hepatitis B virus (HBV) reverse transcriptase proteins from Iranian treatment-naive chronic HBV patients.
Hepatitis monthly. 2013;13(1):e6712. doi: 10.5812/hepatmon.6712. PubMed PMID: 23596461; PubMed Central PMCID: PMCPMC3626233.
23. Zhang Q, Liao Y, Cai B, Li Y, Li L, Zhang J, et al. Incidence of natural resistance mutations in naive chronic hepatitis B patients: a systematic review and meta-analysis. Journal of gastroenterology and hepatology.
2015;30(2):252-61. Epub 2014/10/17. doi: 10.1111/jgh.12831. PubMed PMID: 25318660.
24. Kweon S, Kim Y, Jang MJ, Kim Y, Kim K, Choi S, et al. Data resource profile: the Korea National Health and Nutrition Examination Survey (KNHANES). Int J Epidemiol. 2014;43(1):69-77. doi: 10.1093/ije/dyt228.
PubMed PMID: 24585853; PubMed Central PMCID: PMCPMC3937975.
25. Song BC, Cui XJ, Kim H. Hepatitis B virus genotypes in Korea: an endemic area of hepatitis B virus infection. Intervirology. 2005;48(2-3):133- 7. doi: 10.1159/000081740. PubMed PMID: 15812186.
26. Kim H, Lee SA, Kim DW, Lee SH, Kim BJ. Naturally occurring mutations in large surface genes related to occult infection of hepatitis B virus genotype C. PLoS One. 2013;8(1):e54486. Epub 2013/01/26. doi:
10.1371/journal.pone.0054486. PubMed PMID: 23349904; PubMed Central
43 PMCID: PMCPMC3548799.
27. Lee SA, Kim KJ, Kim DW, Kim BJ. Male-specific W4P/R mutation in the pre-S1 region of hepatitis B virus, increasing the risk of progression of liver diseases in chronic patients. Journal of clinical microbiology.
2013;51(12):3928-36. Epub 2013/09/13. doi: 10.1128/jcm.01505-13.
PubMed PMID: 24025913; PubMed Central PMCID: PMCPMC3838094.
28. Kim H, Gong JR, Lee SA, Kim BJ. Discovery of a Novel Mutation (X8Del) Resulting in an 8-bp Deletion in the Hepatitis B Virus X Gene Associated with Occult Infection in Korean Vaccinated Individuals. PLoS One. 2015;10(10):e0139551. Epub 2015/10/06. doi:
10.1371/journal.pone.0139551. PubMed PMID: 26437447; PubMed Central PMCID: PMCPMC4593592.
29. Lee SA, Kim H, Won YS, Seok SH, Na Y, Shin HB, et al. Male-specific hepatitis B virus large surface protein variant W4P potentiates tumorigenicity and induces gender disparity. Molecular cancer. 2015;14:23.
Epub 2015/02/04. doi: 10.1186/s12943-015-0303-7. PubMed PMID:
25645622; PubMed Central PMCID: PMCPMC4326317.
30. Lee SA, Kim KJ, Kim H, Choi WH, Won YS, Kim BJ. Hepatitis B virus preS1 deletion is related to viral replication increase and disease progression.
World journal of gastroenterology. 2015;21(16):5039-48. Epub 2015/05/07.
doi: 10.3748/wjg.v21.i16.5039. PubMed PMID: 25945020; PubMed Central PMCID: PMCPMC4408479.
44
31. Kramvis A, Bukofzer S, Kew MC. Comparison of hepatitis B virus DNA extractions from serum by the QIAamp blood kit, GeneReleaser, and the phenol-chloroform method. Journal of clinical microbiology.
1996;34(11):2731-3. Epub 1996/11/01. PubMed PMID: 8897174; PubMed Central PMCID: PMCPMC229395.
32. Kumar S, Tamura K, Jakobsen IB, Nei M. MEGA2: molecular evolutionary genetics analysis software. Bioinformatics (Oxford, England).
2001;17(12):1244-5. Epub 2001/12/26. PubMed PMID: 11751241.
33. Rhee SY, Margeridon-Thermet S, Nguyen MH, Liu TF, Kagan RM, Beggel B, et al. Hepatitis B virus reverse transcriptase sequence variant database for sequence analysis and mutation discovery. Antiviral research.
2010;88(3):269-75. Epub 2010/09/30. doi: 10.1016/j.antiviral.2010.09.012.
PubMed PMID: 20875460; PubMed Central PMCID: PMCPMC4374605.
34. Karki R, Pandya D, Elston RC, Ferlini C. Defining "mutation" and
"polymorphism" in the era of personal genomics. BMC Med Genomics.
2015;8:37. doi: 10.1186/s12920-015-0115-z. PubMed PMID: 26173390;
PubMed Central PMCID: PMCPMC4502642.
35. Park SG, Lee SM, Jung G. Antisense oligodeoxynucleotides targeted against molecular chaperonin Hsp60 block human hepatitis B virus replication. J Biol Chem. 2003;278(41):39851-7. doi:
10.1074/jbc.M301618200. PubMed PMID: 12869561.
36. Masaadeh HA, Hayajneh WA, Alqudah EA. Hepatitis B virus genotypes
45
and lamivudine resistance mutations in Jordan. World journal of gastroenterology. 2008;14(47):7231-4. Epub 2008/12/17. PubMed PMID:
19084939; PubMed Central PMCID: PMCPMC2776882.
37. Vutien P, Trinh HN, Garcia RT, Nguyen HA, Levitt BS, Nguyen K, et al.
Mutations in HBV DNA polymerase associated with nucleos(t)ide resistance are rare in treatment-naive patients. Clinical gastroenterology and hepatology : the official clinical practice journal of the American Gastroenterological Association. 2014;12(8):1363-70. Epub 2013/12/18. doi:
10.1016/j.cgh.2013.11.036. PubMed PMID: 24342744.
38. Xu J, Wu B, Wang JH, Huang L, Wang DY, Zhao L, et al. Pre-existing mutations in reverse transcriptase of hepatitis B virus in treatment-naive Chinese patients with chronic hepatitis B. PLoS One. 2015;10(3):e0117429.
Epub 2015/03/31. doi: 10.1371/journal.pone.0117429. PubMed PMID:
25821965; PubMed Central PMCID: PMCPMC4379075.
39. Liu BM, Li T, Xu J, Li XG, Dong JP, Yan P, et al. Characterization of potential antiviral resistance mutations in hepatitis B virus reverse transcriptase sequences in treatment-naive Chinese patients. Antiviral research. 2010;85(3):512-9. Epub 2009/12/26. doi:
10.1016/j.antiviral.2009.12.006. PubMed PMID: 20034521.
40. Song BC, Suh DJ, Lee HC, Chung YH, Lee YS. Hepatitis B e antigen seroconversion after lamivudine therapy is not durable in patients with chronic hepatitis B in Korea. Hepatology (Baltimore, Md). 2000;32(4 Pt
46
1):803-6. doi: 10.1053/jhep.2000.16665. PubMed PMID: 11003626.
41. Carman WF, Zanetti AR, Karayiannis P, Waters J, Manzillo G, Tanzi E, et al. Vaccine-induced escape mutant of hepatitis B virus. Lancet (London, England). 1990;336(8711):325-9. PubMed PMID: 1697396.
42. Lisman T, Porte RJ. Mechanisms of platelet-mediated liver regeneration.
Blood. 2016;128(5):625-9. doi: 10.1182/blood-2016-04-692665. PubMed PMID: 27297793.
43. Zoulim F, Locarnini S. Hepatitis B virus resistance to nucleos(t)ide analogues. Gastroenterology. 2009;137(5):1593-608.e1-2. Epub 2009/09/10.
doi: 10.1053/j.gastro.2009.08.063. PubMed PMID: 19737565.
44. De Clercq E. Clinical potential of the acyclic nucleoside phosphonates cidofovir, adefovir, and tenofovir in treatment of DNA virus and retrovirus infections. Clin Microbiol Rev. 2003;16(4):569-96. PubMed PMID:
14557287; PubMed Central PMCID: PMCPMC207110.
45. Mahmoud Ali M. Mutation Patterns at Codons Rt204 And Rt180 of the HBV Polymerase Gene Associated with Lamivudine Resistance in Treated and Untreated Chronic HBV Patients in Kuwait: A Case Series. Journal of Clinical Case Reports. 2013;03(06). doi: 10.4172/2165-7920.1000276.
46. Lee SH, Kim HS, Byun IS, Jeong SW, Kim SG, Jang JY, et al. Pre- existing YMDD mutants in treatment-naive patients with chronic hepatitis B are not selected during lamivudine therapy. Journal of medical virology.
2012;84(2):217-22. doi: 10.1002/jmv.23191. PubMed PMID: 22170540.
47
47. Warner N, Locarnini S, Kuiper M, Bartholomeusz A, Ayres A, Yuen L, et al. The L80I substitution in the reverse transcriptase domain of the hepatitis B virus polymerase is associated with lamivudine resistance and enhanced viral replication in vitro. Antimicrob Agents Chemother. 2007;51(7):2285- 92. doi: 10.1128/AAC.01499-06. PubMed PMID: 17438047; PubMed Central PMCID: PMCPMC1913255.
48. Chang MH. Natural history and clinical management of chronic hepatitis B virus infection in children. Hepatology international. 2008;2(Supplement 1):28-36. doi: 10.100