21(4) : 240-247 (2015)
http://dx.doi.org/10.20307/nps.2015.21.4.240
240
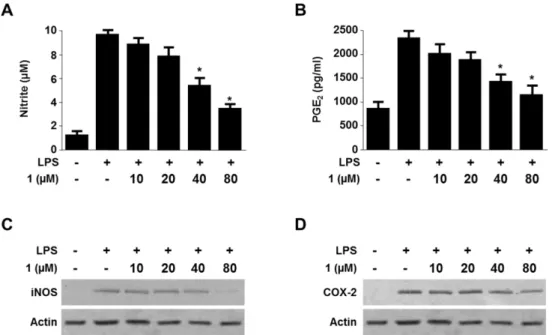
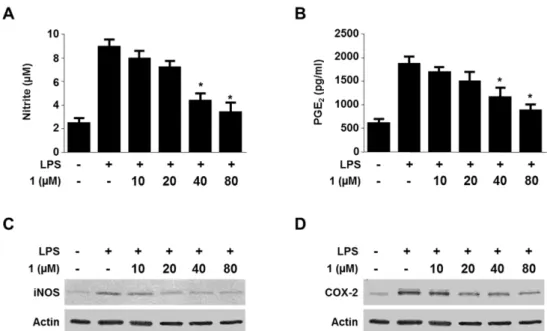
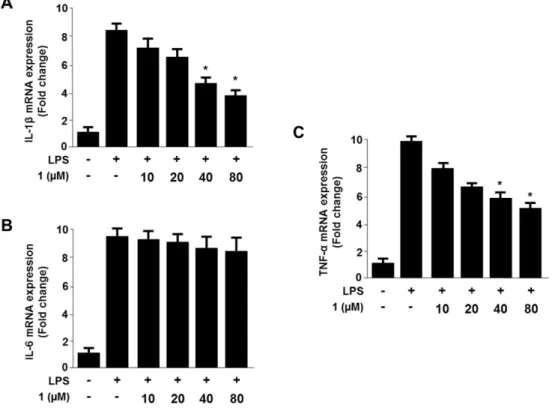
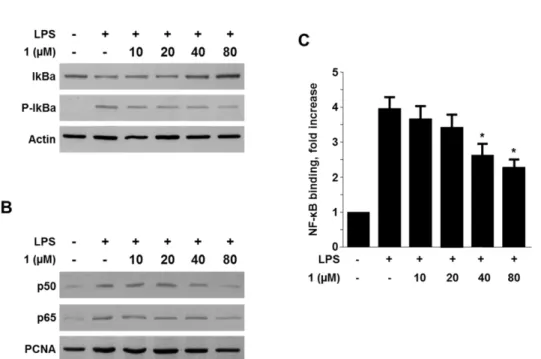
Viridicatol from Marine-derived Fungal Strain Penicillium sp. SF-5295 Exerts Anti-inflammatory Effects through Inhibiting NF- κB Signaling Pathway
on Lipopolysaccharide-induced RAW264.7 and BV2 Cells
Wonmin Ko
1, Jae Hak Sohn
2, Youn-Chul Kim
1,*, and Hyuncheol Oh
1,*
1
College of Pharmacy, Wonkwang University, Iksan 570-749, Korea
2