신경모세포종에서 Alk 유전자의 변이
전 인 상
가천의과학대학교 의학전문대학원 소아과학교실
Alteration of Alk in Neuroblastoma
In-Sang Jeon, M.D.
Department of Pediatrics, Graduate School of Medicine, Gachon University of Medicine and Science, Incheon, Korea
Neuroblastoma, one of the common pediatric solid tumor, shows variable clinical out- come depending on the prognostic factors. Several molecular changes including N-myc amplification have been elucidated as one of the prognostic factor. However, their role in the pathogenesis of neuroblastoma is still not clearly documented. Recently, an alter- ation of Alk, a tyrosine kinase receptor gene preferentially expressed in the central and peripheral nervous systems, was observed in neuroblastoma. Germ line and somatic Alk amplification were found around 25% of familial and sporadic cases of neuroblastoma.
In addition, about 20 types of activating Alk mutations were reported, too. Most of the mutations were missense mutation and localized within the tyrosine kinase domain of the Alk. The most common types of mutation were F1174L and R1275Q, which occupy- ing about 60% of total cases. These alterations were mainly observed in the advanced cases of neuroblastoma or with N-myc amplification. This observable fact suggests that the Alk alteration could be used as an prognostic factor of neuroblastoma. It is interesting that the neuroblastoma is related to the activating Alk-driven pathway. Moreover, the understanding of the molecular oncogenesis of the neuroblastoma would be advanced through the study of the altered tyrosine kinase receptor gene, Alk. Besides, this finding could be applied to inhibit Alk signaling as an effective molecular therapy of neuroblastoma.
pISSN 2233-5250 / eISSN 2233-4580 Clin Pediatr Hematol Oncol 2011;18:1∼7
Received on March 29, 2011 Accepted on April 13, 2011
책임저자: 전 인 상 인천시 남동구 구월동 1198 가천의과학대학교 길병원 소아청소년과, 405-760 Tel: 032-460-8382 Fax: 032-460-3224
E-mail: [email protected]
Key Words: Neuroblastoma, Alk, Tyrosine kinase
서 론
신경모세포종은 교감신경계를 형성하는 원시신경능세포에 서 기원하는 소아 종양이다. 이 종양은 소아기에 발생하는 고 형종양 중 중추신경계 종양을 제외하면 가장 흔한 것으로 전 체 소아 종양의 약 7%를 차지하며 소아 악성 종양에 의한 사 망 원인의 15%를 차지한다[1].
이 종양은 임상 양상이 다양하여 특별한 치료 없이 스스로
위축되어 사라지기도 하고 일부에서는 강력한 복합 치료에도 불구하고 치료가 되지 않는다[2]. 이러한 현상은 신경모세포종 의 분자유전학적 변이와 일부 관련이 있을 것으로 생각된다.
실제로 신경모세포종에서는 여러 분자유전학적 변이들이 관 찰되고 있는데, 대표적인 것으로 N-myc 유전자의 증폭이 있 으며 그 이외에 염색체 1p 또는 11q 부위의 결손이 있다[3].
그러나 신경모세포종에서 N-myc 유전자 증폭의 분자생물학 적 이해는 미흡하다. 또한, 1p와 11q에 위치할 것으로 생각되 는 암억제유전자가 아직 알려져 있지 않아 이들을 통해 신경
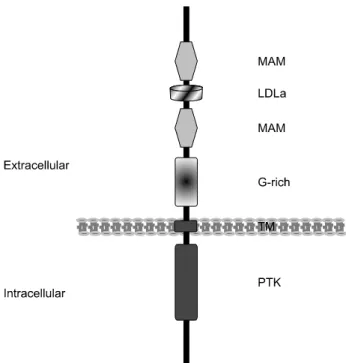
Fig. 1. Structure of human ALK. Alk has the typical feature of tyrosine kinase receptor. The N-terminal region comprises two MAM domains, one LDLa domain, and a glycine rich domain.
This region is suspicious to contribute to the binding of ligand.
Intracellular domain of protein tyrosine kinase (PTK) locates in the C-terminal region. MAM, meprin, A5 protein and receptor protein tyrosine phosphatase Mu; LDLa, low density lipoprotein class A; TM, transmembrane; G-rich, glycine rich.
모세포종을 이해하고 나아가 치료에 응용될 분자학적 표적을 아직 찾지 못하고 있다.
최근에는 암유전자인 Alk (Anaplastic lymphoma kinase)가 신경모세포종의 발생에 관여하며 예후인자로도 관련이 있다 는 것이 알려졌다. 실제로 Alk 유전자의 생식선 돌연변이 (germline mutation)가 가족형 신경모세포종에서 발견되었고 또한 체세포 돌연변이(somatic mutation)가 산발형 신경모세 포종에서 발견되었다[4-8]. Alk 유전자는 역형성큰세포 림프 종(anaplastic large cell lymphoma)에서 관찰되는 염색체전 좌 t(2;5)(p23;q35)에 의해 NPM (nucleophosmin)과 융합되어 생성되는 Alk-NPM의 융합유전자 형태로 처음 알려졌다[9].
Alk는 세포막에 걸쳐있는 전형적인 타이로신 키나제 수용체 로 세포외 리간드 부착영역, 세포막횡단(transmembrane) 영 역, 세포내 타이로신 키나제 영역으로 구성되어 있다. 구조적 유사성을 기준으로 분류하면 LTK (leukocyte tyrosine kinase) 에 속하며 IR (insulin receptor) 가계군에 속한다[10]. 이 유전 자의 변이가 역형성큰세포 림프종 이외에 비소세포성 폐암과 다른 고형종형에서 발견되었다[11].
타이로신 키나제 수용체는 세포신호전달 과정에 관여해 세 포의 증식, 분화, 생존, 운동성 및 형질전환과 같은 다양한 세 포의 생명 현상에 관여를 한다. 또한 수용체의 변이에 의해 이러한 과정에 이상이 생겨 악성화가 되는 분자학적 기전은 잘 알려져 있다. 타이로신 키나제 수용체인 Alk의 변이에 의 해 신경모세포종이 발생한다는 사실은 이 종양을 더 이해하는 데 도움을 줄 것이다. 또한 치료 부분에도 응용할 수 있어 Alk 에 대한 연구를 통해 예후가 불량한 고위험군의 치료 성적을 향상시킬 수 있을 것이며 현재 실제로 Alk의 변이를 표적으로 한 치료 연구가 진행되고 있다.
이 논문에서는 Alk 유전자의 분자생물학적 행동 양식과 신 경모세포종에서 관찰되는 변이와 발병에 관여하는 것으로 생 각되는 분자학적 기전을 알아보고 향후 치료로 응용될 부분에 대해 알아보려고 한다.
타이로신 키나제 수용체인 Alk
1) Alk의 구조적 특징
인체 Alk 유전자에는 1,620개에 달하는 아미노산의 유전코 드가 입력되어 있으며 이 유전자에 의해 만들어지는 단백질의 크기는 약 180 kDa이다. 그러나 N-연결 당화(N-linked glyco- sylation) 등과 같은 번역후 변형(post-translational mod- ification)에 의해 실질적인 단백질의 크기는 220 kDa이다[12].
Alk의 세포외 영역은 여러 부위로 나눌 수 있다. 가장 말단
부위에는 N-말단 시그널 펩티드가 있다. 이어 MAM (Meprin, A5 protein and receptor protein tyrosine phosphatase Mu) 영역, LDLa (Low Density Lipoprotein class A) 영역이 있다.
막의 바로 바깥 부위에는 글라이신 풍부 구역(glysine rich domain)이 있다. MAM은 세포 사이의 상호작용에 관여하며, LDLa는 리간드 부착에 관여하는 것으로 생각되나 이들 영역 의 구체적인 역할은 아직 알려져 있지 않다(Fig. 1).
세포 내에는 타이로신 키나제가 있는데, 이 부위는 보존이 가장 잘 되어 있어 종마다 차이가 거의 없다. 실제로 마우스와 인간의 Alk는 단 4개의 아미노산만이 다르다. 키나제 영역의 활성화 고리(activation loop)에는 IR 가계군이 공통적으로 갖 고 있는 YXXXY (Y: 타이로신, X: 다른 아미노산) 모티프 (motif)가 있다. 이 부위는 타이로신 키나제가 자동인산화 (autophosphorylation)되는데 중요한 역할을 한다. Alk는 YRASYY (R:아르기닌, A: 알라닌, S: 세린) 모티프를 갖고 있 는데 첫 번째 Y (Tyr1278)가 자동인산화되는 과정의 열쇠를 쥐 고 있는 것으로 밝혀졌다. 또한 중간에 있는 RAS가 Tyr1278이 자동인산화를 시작하는데 영향을 미치는 것이 알려졌다[13].
2) Alk의 생리적인 역할
Alk 유전자는 여러 종(species)들에서 아주 잘 보존되어 있 다. Alk 유전자의 생리적인 기능은 주로 무척추동물에서 알려 져 있다. 초파리(Drosphila melanogaster) Alk는 몸중배엽 (somatic mesoderm)에서 분비되는 리간드인 Jelly belly (JEB)에 의해 활성화된다. JEB가 Alk 표면에 부착하게 되면 Alk는 활성화되어 Ras-mitogen-activated protein kinase (MAPK) 경로를 활성화하고 근육 융합에 관여하는 하위 유전 자인 DUF와 ORG-1를 발현시켜 창자 근육조직의 형성을 지휘 한다. JEB는 또한 광수용체 축색에서 표현되는데 이 JEB는 시각엽(optic lobe)의 표적 신경세포에 있는 Alk를 활성화시 키고 초파리의 시각계를 이루는 신경 회로 조합을 중재한다.
예쁜꼬마선충(Caenorhabditis elegans)에서는 Alk 상동체가 신경근육이음부(neuromuscular junction)의 시냅스전에 위치 하는데 여기서 Alk 상동체는 proteasome 매개 분해에 의해 정확하게 조절된다. 이러한 현상은 Alk가 시냅스형성을 안정 화하는데 필요한 것을 시사한다[14].
포유류 Alk는 주로 중추 및 말초신경계에서 발현된다 [10,12]. 인위적으로 Alk 유전자의 발현을 유도하는 융합 Alk 수용체는 여러 세포주에서 신경세포 분화를 유도하며 세포를 형질전환시키고 유사분열을 촉진시킨다. Alk 결핍 마우스는 외관상 이상이 없고 조직에 이상은 없으나 해마(hippocam- pus)의 원시세포가 증가되어 있고 비정상적인 행동을 한다.
이러한 현상은 Alk 유전자의 이상이 신경 기원 종양인 신경모 세포종의 발병과 관련이 있음을 시사하고 있다. 그러나 아직 Alk의 변이와 신경모세포종의 발병과의 상호 관련에 대한 연 구는 미흡하다.
무척추 동물과 달리 포유류 Alk 수용체는 JEB 리간드에 반 응을 보이지 않는다. 또한 JEB와 유사한 물질이 아직 확인되 지 않아 포유류 Alk는 다른 리간드와 반응하는 쪽으로 진화 한 것으로 추측된다. 포유류에서 Alk의 리간드로 생각되는 것 으로 pleiotrophin (PTN)과 midkine (MK)가 있다[15,16]. 이 두 리간드는 뉴론 발달에 관여하며 퇴행성 신경질환과도 관련 이 있다. 실제로 Alk는 척추 운동 뉴론에서 PTN의 활동을 매 개한다.
Alk는 리간드가 붙지 않고 비정상적으로 표현되면 아포프 토시스를 유도하며 리간드가 붙으면 아포프토시스를 방해한 다. 즉, 리간드에 따라 행동하는 의존성 수용체(dependence receptor)라고 할 수 있다. 그러나 PTN에 의한 Alk의 인산화 가 나타나지 않는 세포계도 있어 PTN이 엄밀한 의미의 리간 드인지 확실하지 않다[17]. 또한, PTN, MK 이외로 Alk를 활성
화시키는 다른 물질이 있는지 향후 더 많은 연구가 필요하다.
3) Alk의 발암 과정
Alk의 발암과정은 주로 역형성큰세포 림프종에서 관찰되는 NPM/Alk의 융합유전자에 대한 연구로 알려져 있다. 유전자 융합에 의해 Alk가 활성화되면 Alk가 갖고 있는 타이로신 키 나아제는 비정상적으로 행동하여 암을 유발하게 된다. 즉, 비 정상적으로 활성화된 Alk는 세포의 증식을 유도하고 아포프 토시스를 방해해 세포가 사멸되지 않게 하며 세포뼈대를 재배 열시키며 세포 형태를 변형시킨다. Alk의 암유전자화는 Alk의 표적 물질인 하위 분자(downstream molecule)와의 상호작용 에 의해 이루어지는데, 하위 분자는 세포내 신호전달을 매개 하는 물질이다[18]. Alk는 정상이거나 암유전자화한 다른 타 이로신 키나제와 연결되어 상호작용을 하거나 여러 종류의 다 른 경로들을 활성화시킨다. 현재까지 알려져 있는 경로는 크 게 세 가지로 나눌 수 있다. 첫째 가장 잘 알려진 성장인자 신호 전달 체계인 Ras/MAPK (Ras-extracellular signal-regu- lated kinase, ERK) 경로이며 둘째는 Janus Kinase 3 (JAK3)- STAT3 경로이며 셋째는 phosphatidylinositol 3-kinase (PI3K)- Akt 경로이다. 이들은 서로 긴밀하게 상호작용을 하여 Alk의 작용을 매개한다. 여기서 Ras-ERK는 세포증식과 관련이 있으며 뒤의 두 경로는 세포 생존 및 표현형 변형과 관련이 있다[19].
신경모세포종에서 Alk 유전자의 증폭 및 돌연변이가 관찰 되는데 아직 이들이 신경모세포종의 발생을 일으키는 기전은 아직 확실히 알려져 있지 않다. 현재까지 알려진 바에 의하면 Ras-ERK 경로와 phosphatidylinositol 3-kinase (PI3K)-Akt 경 로와 관련이 있는 것으로 생각된다[19]. 신경모세포종과 신경 모세포종 세포주에서 활성화된 Alk 유전자는 Ras-ERK 경로에 서 하위 분자인 Shc (SH2 domain-containing transforming protein)의 활성화를 유도한다[20]. Shc는 도킹(docking) 단백 질로 활성화된 타이로신 키나제 수용체의 phosphotyrosine 잔유기에 부착하여 세포내 신호를 전달하는 역할을 한다. Shc 단백질은 N-말단 부위에는 phosphotyrosine binding (PTB) 구역, 중앙에는 glycine/proline 풍부 구역(CH1), C-말단 부위 에는 Src homology 2(SH2) 구역으로 이루어져 있다[21]. Shc 는 PTB 구역에 의해 활성화된 Alk의 타이로신 키나제에 부착 하게 되고 C-말단 부위에 있는 SH2 구역을 통해 Grb2, SRC, Abl, PI3-K 같은 물질을 불러 모은다. Grb2는 SH2-containing adaptor 분자를 통해 Shc에 부착하게 되면 인산화되어 활성화 되어 자신의 다른 부분인 SH3 구역을 통해 교환 단백질인 SOS (son of sevenless)을 불러 모은다. SOS는 GDP가 GTP로 교환되는 과정에서 촉매 역할을 한다. Ras는 GTP와 붙게 되
Fig. 2. The signal transduction of Alk via Ras/MAPK (Ras-extracellular signal-regulated kinase, ERK) path- way. The altered Alk activate the Shc aberrantly. Subsequently the pathway is activated and the target gene is expressed aberrantly.
면 RAF를 활성화시키고 이어서 활성화된 RAF는 하부경로 실 행자인 MAPK (mitogen-activated protein kinase)의 연쇄 반 응을 활성화하여 세포외 신호를 세포 내로 전달한다(Fig. 2).
Shc 단백질에는 ShcA, ShcB, SchC 3 종류가 있다[22]. ShcA 는 전 기관에서 발현되며, ShcB도 여러 기관에서 발현되나 특히 뇌와 척수에서 특이하게 발현된다. 그에 반해 ShcC는 마우스의 신경계통에서만 발현되며 특이하게 배아 뇌에선 낮 게 발현되다가 출생 후 발현이 급격히 증가한다. ShcB와 ShcC이 부족한 마우스에선 정상적인 뉴론의 형성이 되지 않 아 이 두 분자는 신경성장 인자 수용체인 TrkA 신호전달에서 중요한 역할을 하는 것으로 생각된다. 대부분의 신경모세포종 에서는 ShcC의 타이로신 인산화가 관찰되나 ShcB는 낮게 발 현된다. 이러한 상황을 보면 Alk의 변이에 의해 신경세포 형 성에 관여하는 Shc에 이상이 오게 되어 신경능 세포에서 기원 하는 신경모세포의 발병과 관련이 있을 것으로 유추할 수 있다.
한편 ShcC 단백질이 신경모세포종이 진행되거나 N-myc 증 폭이 있는 예후 불량군에서 높게 나타나서 ShcC는 신경모세 포종의 예후인자가 될 수 있음을 시사한다[23]. 또한 ShcC의 발현을 방해하면 정상적인 신경 분화가 일어나, ShcC는 신경 모세포종의 정상 분화를 방해하는 것으로 생각된다. 이러한 상황을 고려해 보면 ShcC는 신경모세포종 치료의 표적이 될 수 있음을 시사하고 있다.
활성화된 Alk는 phosphatidylinosito 3-kinase (PI3K)-Akt 경로를 통해 Akt를 활성화시키는데 신경모세포종에서 Akt가 활성화되어 아포프토시스를 방해하는 것으로 알려져 있다[23].
Alk의 변이
신경모세포종에서 정상 Alk가 표현되는 것은 2000년도에 처음 알려졌다[24]. 그러나 구체적으로 신경모세포종 검체나 신경모세포종 세포주에서 Alk가 증폭되어 있고 돌연변이가 있다는 것은 2008년도에 처음 보고되었다[5-8]. 현재까지 알려 진 바에 의하면 신경모세포종에서 Alk 유전자의 증폭이 약 25%에서 나타난다. 또한 가족형과 산발형 신경모세포종의 10% 내외에서 Alk의 계속적인 활성화를 유도하는 돌연변이가 관찰된다. 특히 돌연변이는 Alk의 키나제 영역에서 주로 일어 난다[25].
먼저 Mosse 등은 491개의 신경모세포종 검체에서 약 26%
의 Alk 유전자 증폭이 있다고 보고했다[5]. 또한 이 경우는 진 단 시 전이가 되어 예후가 불량한 환아에서 주로 나타났으며 사망률도 높았다. 이러한 현상은 Alk 유전자의 증폭이 예후가 불량한 환아군에서 많았다는 Passon 등의 보고와 일치하여 Alk 유전자의 증폭은 신경모세포종의 예후 인자의 하나가 될 수 있다는 것을 시사한다. 이들은 가족형 신경모세포종 8 가 계에서 Alk의 생식선돌연변이(germline mutation) 3 종류를 보고하였는데 돌연변이는 missense mutation이었다. 또한, 고 위험군 환아의 신경모세포종 검체 194개를 분석하여 그 중 24개(12.4%)에서 체세포 돌연변이(somatic mutation)가 있다 고 보고하였다. 이들 돌연변이는 9 종류였는데 그 중 8 종류는 키나제 영역에서 돌연변이가 있었다. 한편, 이들 돌연변이에 의해 Alk는 계속 인산화가 되었으며. 활성화된 Alk m-RNA를
Nucleotide changes Amino acid mutation Targeted region Incidence (n=72) 3260 C → T
3271 G → A 3383 G → C 3452 C → T 3497 T → G 3512 T → A 3520 T → A 3521 T → G 3520 T → G 3522 C → A/G 3575 G → C 3700 G → A 3733 T → G 3733 T → A 3735 C → A/G 3734 T → G 3749 T → C 3824 G → T 3824 G → A 3833 A → C
K1062M T1087I D1091N G1128A T1151M M1166R I1171N F1174I F1174C F1174V F1174L R1192P A1234T F1245V F1245L F1245C I1250T R1245L R1275Q Y1278S
Juxtamembrane domain Juxtamembrane domain P loop
Kinase domain C helix C helix End of C helix End of C helix End of C helix End of C helix β4 strand Catalytic loop?
Catalytic loop Catalytic loop Catalytic loop Catalytic loop Catalytic loop Activation loop Activation loop Activation loop?
1 1 1 1 2 1 2 1 1 1 25 2 2 1 1 3 1 1 23 1 Table 1. A class of mutations in Alk in neuroblastoma
녹다운(knockdown)시키면 신경모세포의 증식이 억제되었다.
두 번째 연구로 Janoueix-Lerosey 등은 592 신경모세포종 검체를 조사하여 약 25%에서 Alk 유전자의 증폭이 있다고 보 고하였다[6]. 두 가계에서 R1275Q 이형접합 돌연변이와 R1192P 이형접합 돌연변이를 발견하였다. 또한, 신경모세포 종 검체 143개를 분석하여 그 중 16개(12.4%)에서 돌연변이 가 있다고 보고하였다.
George 등은 93 개의 신경모세포종 환아 검체에서 Alk 유 전자의 염기서열을 분석하여 8명(8.6%)에서 다섯 종류의 mis- sense 돌연변이를 발견하였다. 돌연변이는 모두 Alk의 타이로 신 키나제 구역에서 있었다[7]. 이들 돌연변이 중 두 개는 생식 선 돌연변이였으며, 여섯 개는 체세포 돌연변이였다. 네 검체 에서 발견된 흔한 돌연변이형은 엑손 23에서 시토신이 아데닌 으로 바뀌어 페닐알라닌이 루이신으로 변형된 F1174L이었다.
특히 F1174L과 R1275Q는 인터루킨-3에 의존해 성장하는 murine hematopoietic Ba/F3 세포를 시토카인과 상관없이 성 장하는 세포로 형질전환시켜 이 두 돌연변이형은 타이로신 키 나제를 계속적으로 활성화시키는 것을 시사하였다.
마지막으로 일본에서 Chen 등은 215개의 신경모세포종 환 아 검체와 24개의 신경모세포종 세포주를 대상으로 Alk 유전 자의 변이를 조사하여 각각 13 (6.1%), 8 (33.3%) 개에서 돌연 변이를 발견하였다[8]. 그런데, Alk 돌연변이가 있는 13예에서 12예는 병기 3이나 4의 고위험군이었다. 또한, Alk 유전자의
돌연변이와 N-myc 유전자 증폭은 상관관계가 있었다. 또한 151예의 고위험군 환아를 대상으로 조사한 바에 의하면 16 (11%)에서 Alk 유전자의 증폭 등 비정상적인 신호전달이 관 찰되었다.
이들 네 보고를 종합하면 20 종류의 돌연변이가 생식선세 포 및 체세포에서 관찰되었다. 가장 흔한 형은 F1174L R1275Q로 각각 약 30% 합해 60%를 차지했다. 한편 이 두 돌연변이형은 배양세포를 형질전환시킨다(Table 1).
표적치료로의 응용
종양형성에 관여하는 분자생물학적 지식이 늘어남에 따라 이러한 지식을 응용해 종양 치료에 적용하는 표적치료가 현재 여러 종양에서 시행되고 있다. 가장 대표적이라고 할 수 있는 것은 imatinib (GleevecⓇ)이다. 이 약물은 만성골수성백혈병 에서 관찰되는 필라델피아 염색체에서 염색체 전좌에 의해 형 성되는 Bcr-Abl 융합유전자의 작용을 억제하여 항암 작용을 한다. 타이로신 키나제 방해제인 소분자 물질 imatinib는 만성 골수성백혈병 환자에 투여 시 만족할 만한 성과를 거두고 있 어 다른 타이로신 키나제 방해제 약물을 개발하는 새로운 장 을 열어주었다. 그 후 타이로신 키나제 방해제로 항암 효과를 나타내는 약물로는 비소세포성 폐암 치료제로 사용되는 EGFR (epidermal growth factor receptor) 타이로신 키나제
방해제인 gefitinib (IressaⓇ)와 erlotinib (TarcevaⓇ)가 있다 [26]. 그 외 sorafenib와 sunitinib도 타이로신 키나제 방해제로 신세포암종 치료제로 사용된다[27].
타이로신 키나제 유전자인 Alk의 활성화가 인체 여러 종양 에서 발견되어 이 유전자도 표적치료의 목표물이 될 수 있다.
현재는 Alk의 키나제의 활성화 영역에 대한 결정 구조가 아직 알려져 있지 않고, Alk 유전자가 관여하는 종양의 발생빈도가 높지 않아 시장성이 낮아 이 유전자를 표적으로 하는 치료제 개발은 어려움이 있다. 또한 치료제 개발이 연구가 많이 되어 있는 역형성큰세포 림프종에서 관찰되는 Alk-NPM를 대상으 로 하고 있다. 그러나 신경모세포종에서 관찰되는 Alk 유전자 의 활성화도 이들 약물을 치료제로 사용하는 데는 큰 문제는 없을 것으로 생각된다. 현재 임상 1 상중인 약물로는 ATP 경 쟁성 c-Met/HGFR와 Alk 타이로신 키나제 방해제인 소분자 PF-02341066d 있다. 이 약물은 경구용 약물로 역형성큰세포 림프종 실험동물에서 종양의 쇠퇴가 있었다[28]. 그러나 현재 임상 시험 중으로 더 많은 자료가 필요하다. 그 외에 개발된 약물로 NVP-TAE684가 있다. 이 약물은 역형성큰세포 림프종 세포주 이외에 신경모세포종 세포주에서도 종양의 크기를 감 소시키는 효과가 있었다[29]. 그 외에 현재 개발 중인 약물로 CRL151104A와 WZ-5-126이 있는데 아직 약리기전과 효과를 알려면 시간이 더 필요하다[30].
결 론
전이가 많이 되어 있거나 N-myc 유전자의 증폭이 있는 등 예후 불량 신경모세포종은 강력한 치료에도 불구하고 치료가 어렵다. 최근에는 조혈모세포 이식을 포함한 강력한 항암치료 및 방사선 동위원소를 이용한 MIBG 치료 등이 시도되고 있으 나 여전히 치료 성적은 만족스럽지 못하다. 비호지킨 림프종 의 한 아형인 역형성큰세포 림프종에서 처음 알려진 Alk 유전 자는 전형적인 타이로신 키나제를 갖고 있는 수용체이다. 신 호전달 경로에서 중요한 역할을 하는 타이로신 키나제 수용체 는 비교적 연구가 많이 되어 있으며 이들 수용체의 변이가 각종 종양을 일으키는 기전도 잘 알려져 있다. 타이로신 키나 제 수용체인 Alk의 변이가 일부 신경모세포종에서 발견되어 여러 연구가 진행 중이다. 임상적으로는 특히 Alk 유전자의 변이가 있는 경우 병기가 3,4이거나 N-myc 증폭이 동반되어 이 유전자가 예후인자가 될 수 있음을 시사하고 있다. 또한 Alk에 대한 표적치료제가 개발 중으로 향후 신경모세포종 환 아에서 응용되어 개선된 치료 성적을 얻을 수 있을 것으로 기대된다.
참 고 문 헌
1. Maris JM, Hogarty MD, Bagatell R, Cohn SL. Neuroblastoma.
Lancet 2007;369:2106-20.
2. Sung KW, Chung EH, Yoo KH, et al. Risk-based grouping of patients and risk-directed treatment in neuroblastoma. Korean J Pediatr Hematol Oncol 2001;8:238-49.
3. Brodeur GM. Neuroblastoma: biological insights into a clinical enigma. Nat Rev Cancer 2003;3:203-16.
4. Caren H, Abel F, Kogner P, Martinsson T. High incidence of DNA mutations and gene amplifications of the Alk gene in advanced sporadic neuroblastoma tumours. Biochem J 2008;
416:153-9.
5. Mosse YP, Laudenslager M, Longo L, et al. Identification of Alk as a major familial neuroblastoma predisposition gene.
Nature 2008;455:930-5.
6. Janoueix-Lerosey I, Lequin D, Brugieres L, et al. Somatic and germline activating mutations of the Alk kinase receptor in neuroblastoma. Nature 2008;455:967-70.
7. George RE, Sanda T, Hanna M, et al. Activating mutations in Alk provide a therapeutic target in neuroblastoma. Nature 2008;455:975-8.
8. Chen Y, Takita J, Choi YL, et al. Oncogenic mutations of Alk kinase in neuroblastoma. Nature 2008;455:971-4.
9. Morris SW, Kirstein MN, Valentine MB, et al. Fusion of a kin- ase gene, Alk, to a nucleolar protein gene, NPM, in non- Hodgkin's lymphoma. Science 1994;263:1281-4.
10. Morris SW, Naeve C, Mathew P, et al. Alk, the chromosome 2 gene locus altered by the t(2;5) in non-Hodgkin's lympho- ma, encodes a novel neural receptor tyrosine kinase that is highly related to leukocyte tyrosine kinase (LTK). Oncogene 1997;14:2175-88.
11. Li XQ, Hisaoka M, Shi DR, Zhu XZ, Hashimoto H. Expression of anaplastic lymphoma kinase in soft tissue tumors: an im- munohistochemical and molecular study of 249 cases. Hum Pathol 2004;35:711-21.
12. Iwahara T, Fujimoto J, Wen D, et al. Molecular character- ization of Alk, a receptor tyrosine kinase expressed specifi- cally in the nervous system. Oncogene 1997;14:439-49.
13. Tartari CJ, Gunby RH, Coluccia AM, et al. Characterization of some molecular mechanisms governing autoactivation of the catalytic domain of the anaplastic lymphoma kinase. J Biol Chem 2008;283:3743-50.
14. Liao EH, Hung W, Abrams B, Zhen M. An SCF-like ubiquitin ligase complex that controls presynaptic differentiation. Nature 2004;430:345-50.
15. Stoica GE, Kuo A, Aigner A, et al. Identification of anaplastic lymphoma kinase as a receptor for the growth factor pleiotrophin. J Biol Chem 2001;276:16772-9.
16. Stoica GE, Kuo A, Powers C, et al. Midkine binds to anaplastic lymphoma kinase (Alk) and acts as a growth factor for differ-
ent cell types. J Biol Chem 2002;277:35990-8.
17. Moog-Lutz C, Degoutin J, Gouzi JY, et al. Activation and in- hibition of anaplastic lymphoma kinase receptor tyrosine kin- ase by monoclonal antibodies and absence of agonist activity of pleiotrophin. J Biol Chem 2005;280:26039-48.
18. Bai RY, Ouyang T, Miething C, Morris SW, Peschel C, Duyster J. Nucleophosmin-anaplastic lymphoma kinase associated with anaplastic large-cell lymphoma activates the phosphati- dylinositol 3-kinase/Akt antiapoptotic signaling pathway.
Blood 2000;96:4319-27.
19. Chiarle R, Voena C, Ambrogio C, Piva R, Inghirami G. The anaplastic lymphoma kinase in the pathogenesis of cancer.
Nat Rev Cancer 2008;8:11-23.
20. Miyake I, Hakomori Y, Shinohara A, et al. Activation of ana- plastic lymphoma kinase is responsible for hyperphosphory- lation of ShcC in neuroblastoma cell lines. Oncogene 2002;21:
5823-34.
21. Pelicci G, Dente L, De Giuseppe A, et al. A family of Shc related proteins with conserved PTB, CH1 and SH2 regions.
Oncogene 1996;13:633-41.
22. Nakamura T, Muraoka S, Sanokawa R, Mori N. N-Shc and Sck, two neuronally expressed Shc adapter homologs. Their differ- ential regional expression in the brain and roles in neuro- trophin and Src signaling. J Biol Chem 1998;273:6960-7.
23. Miyake I, Ohira M, Nakagawara A, Sakai R. Distinct role of
ShcC docking protein in the differentiation of neuroblastoma.
Oncogene 2009;28:662-73.
24. Lamant L, Pulford K, Bischof D, et al. Expression of the Alk tyrosine kinase gene in neuroblastoma. Am J Pathol 2000;156:
1711-21.
25. George RE, Attiyeh EF, Li S, et al. Genome-wide analysis of neuroblastomas using high-density single nucleotide poly- morphism arrays. PLoS One 2007;2:e255.
26. Kumar A, Petri ET, Halmos B, Boggon TJ. Structure and clin- ical relevance of the epidermal growth factor receptor in hu- man cancer. J Clin Oncol 2008;26:1742-51.
27. Wilhelm S, Carter C, Lynch M, et al. Discovery and develop- ment of sorafenib: a multikinase inhibitor for treating cancer.
Nat Rev Drug Discov 2006;5:835-44.
28. Christensen JG, Zou HY, Arango ME, et al. Cytoreductive anti- tumor activity of PF-2341066, a novel inhibitor of anaplastic lymphoma kinase and c-Met, in experimental models of ana- plastic large-cell lymphoma. Mol Cancer Ther 2007;6:3314-22.
29. Kelleher FC, McDermott R. The emerging pathogenic and therapeutic importance of the anaplastic lymphoma kinase gene. Eur J Cancer 2010;46:2357-68.
30. Webb TR, Slavish J, George RE, et al. Anaplastic lymphoma kinase: role in cancer pathogenesis and small-molecule in- hibitor development for therapy. Expert Rev Anticancer Ther 2009;9:331-56.