책임저자: 김창주, 서울시 동대문구 경희대로 26
130-701, 경희대학교 의과대학 생리학교실 Tel: 02-961-0407, E-mail: [email protected] 접수: 2012년 11월 15일, 심사: 2013년 5월 15일 게재승인: 2013년 6월 10일
이 논문은 2009년 정부(교육과학기술부)의 재원으로 한국연구재 단의 지원을 받아 수행된 연구임(NRF-2009-352-G00056).
트레드밀 운동이 외상성 뇌손상에 의하여 유발된 흰쥐의 단기기억 장애와 신경세포생성에 미치는 영향
*경희대학교 의과대학 생리학교실, †한남대학교 이과대학 생활체육학과
김 보 균 *ㆍ신 말 순 *ㆍ조 정 완 *ㆍ이 희 혁
†
ㆍ김 창 주 *Treadmill Exercise Alleviates Traumatic Brain Injury-Induced Memory Impairment by Enhancing Cell Proliferation in the Hippocampus of Rats
Bo-Kyun Kim*, Mal-Soon Shin*, Jung-Wan Cho*, Hee-Hyuk Lee
†, Chang-Ju Kim*
*Department of Physiology, College of Medicine, Kyung Hee University, Seoul, †Department of Sports Science, College of Nature Sciences, Han Nam University, Daejeon, Korea
Traumatic brain injury (TBI) induces many neuropsychiatric disorders, and memory loss is the most frequent symptom of TBI.
In the present study, we investigated the effects of treadmill exercise on the short-term memory in relation with cell proliferation in the hippocampus following TBI in rats. The animals were randomly divided into four groups (N=10 in each group):
sham-operation group, sham-operation and treadmill exercise group, TBI-induction group, and TBI-induction and treadmill exercise group. TBI was induced by an electromagnetic-controlled cortical impact. The animals in the treadmill exercise group were forced to run on the treadmill machine for 30 min once daily, for ten consecutive days. Induction of TBI deteriorated short-term memory and inhibited cell proliferation with suppressing brain-derived neurotrophic factor (BDNF) expression in the hippocampus.
Treadmill exercise alleviated TBI-induced short-term memory impairment and also increased cell proliferation with enhancing BDNF expression in the hippocampus. The present results revealed that treadmill exercise overcame the TBI-induced memory impairment through enhancing cell proliferation in the hippocampus. Herein, we suggest that treadmill exercise may be used as the useful strategy for the functional recovery following traumatic brain damage. (Korean J Str Res 2013;21:141∼149)
Key Words: Traumatic brain injury, Treadmill exercise, Short-term memory, Brain-derived neurotrophic factor, Rats
서 론
외상성 뇌손상(traumatic brain injury; TBI)은 퇴행성이 아 닌 외부 충격에 의해 뇌손상을 입는 것으로서 뇌손상의 가 장 흔한 원인중의 하나이다(Flanagan et al., 2006). 산업화, 기 계화, 도시화 그리고 개발을 추구해온 현대 문명의 발전으 로 인하여 외상성 뇌손상 환자는 증가되고 있다. 우리나라
의 경우에도 교통사고 발생율이 세계 1위라는 점을 감안해 볼 때, 외상성 뇌손상 환자 수는 급격히 증가할 것으로 예 측된다. 외상성 뇌손상은 대부분의 경우 진행되는 증상을 보이며, 짧은 기간 내에 뇌에 치명적인 손상을 입히게 된 다. 따라서 뇌경색보다 위험할 정도로 심각한 신경세포의 손상을 유발하여 인지적, 기능적, 형태적으로 회복이 어렵 게 되는 질환이다(Rudehill et al., 2006). 외상성 뇌손상 환자 들의 인지기능 장애 중에서 가장 많이 호소하는 어려움은 기억 장애이다. 기억 장애는 뇌손상의 심각도와 상관없이 환자들이 가장 빈번하게 호소하는 문제이며 다른 인지기 능에 장애에 비해 오랜 기간 지속되는 것으로 알려져 있다 (Ponsford et al., 2008).
포유동물의 내피 세포는 2주마다 새로운 세포로 대체되 고 피부의 상피세포는 몇 개월마다 새로 생성되며, 적혈구 의 수명은 120일로 새로운 적혈구가 계속 생성되지만, 뇌 신경세포의 수명은 생명체의 수명과 같다. 즉 정상 상태에 서 뇌는 완전히 분화된 신경세포들로 구성되어 대부분의 뇌 영역에서는 새로이 세포가 생성되지 않는다는 것이 일 반적이다. 그러나 외측부 뇌실의 뇌실하 영역(subventricular zone, SVZ)과 해마(hippocampus) 치상회(dentate gyrus)의 과립 하 영역(subgranular zone, SGZ) 등 특정 뇌 부위에는 일생동 안 신경 간세포가 존재하여 신경 전구세포(neuronal proge- nitor cells)가 증식(proliferation)되고 생존(survival)하여 신경세 포로 분화(differentiation)되는 신경세포생성이 일어난다(Fuch
et al., 2000).
외상성 뇌손상으로 인한 인지기능의 손상은 주로 뇌의 해마(hippocampus) 영역의 손상 때문이다(Clausen et al., 2005;
Scheff et al., 2005). 해마는 학습과 기억의 형성과정에 관여 하는 중요한 뇌 영역이며, 신경의 발생기뿐만 아니라 성장 후에도 신경세포생성이 활발하게 일어나는 부위이다. 또 한 알츠하이머병 및 파킨슨병과 같은 퇴행성 뇌질환과 (Lopez-Toledano et al., 2004; Junque et al., 2005) 허혈이나 산 화스트레스로부터 가장 손상받기 쉬운 부위이이며, 많은 뇌 영역 가운데 신경가소성(neural plasticity)이 가장 큰 부위 로 알려져 있다(Fuchs et al., 2000; Lie et al., 2004). 이런 이유 로 많은 연구자들은 외상성 뇌손상과 관련하여 일차적으 로 해마부위의 신경세포생성(neurogenesis)과 신경세포사멸 (apoptosis)에 초점을 두고 연구를 꾸준히 진행해 왔으며, 따 라서 외상성 뇌손상은 해마와 시상(thalamus)에 신경세포의 소실(loss)을 야기할 것이라고 제시하였다(Hall et al., 2005).
뇌에서의 인지형성과정에 깊이 관여하는 것으로 알려진
여러 종류의 신경전달물질들은 운동의 영향을 받는다. 운 동은 뇌에서 세로토닌계의 신경조절을 통하여 신경세포 생성을 촉진하는 중요한 인자임이 알려져 있다(Weicker et
al., 2001). 신체운동은 뇌에서의 신경영양인자의 발현을 증
가시키고(Neeper et al., 1996; Trejo et al., 2001), 흥분성 뇌 손 상 기전에 관여하는 glutamate 수용체의 발현을 하향 조절 하는 것으로 보고되었다(Guezennec et al., 1998). 다양한 신 경성장인자들 중, 뇌유래 신경영양인자(brain-derived neuro- trophic factor, BDNF)는 뇌허혈이나 파킨슨 질환과 같은 뇌 질환 동물들에 기능적 회복과 인지기능 발달을 유도하고 (Almli et al., 2000, Griesbach et al., 2004), 외상성 뇌손상 후에 도 해마의 신경세포 생성과 신경생리학적 기능을 향상시 키는 것으로 알려져 있다(Hicks et al., 1999; Kim BK et al., 2011). Lee HH et al.(2009)은 14일간 트레드밀 달리기를 한 쥐 해마에서 BDNF mRNA의 발현이 증가되었음을 보고하 였다.이러한 연구들은 운동이 정상적인 뇌에서 뿐 아니라 손 상된 뇌에서도 새로운 뇌신경세포의 생성을 증가시키거나 또는 뇌신경세포의 손상을 억제함으로써 뇌기능의 회복에 도움이 될 수 있다는 사실을 보여주며, 이를 근거로 현재 퇴행성 뇌질환 환자에게 행동학적ㆍ신경학적 장애를 완화 시키는데 효과적인 방법으로 재활운동이 권장되고 있다.
그러나 재활운동이 외상성 뇌손상 환자의 회복에도 도움 이 되는지에 대한 연구는 국내ㆍ외적으로 보고가 미흡한 실정으로, 이러한 이유로 일부 연구자들은 외상성 뇌손상 환자에 대한 재활운동의 효과성에 의문을 제기하고 있다.
따라서 본 연구는 트레드밀 운동이 외상성 뇌손상 유발 흰쥐에서의 단기기억능력에 미치는 영향을 알아보기 위하 여 passive avoidance task를 실시하였고, 해마 부위에서 새로 운 신경세포의 생성과 BDNF의 발현에 미치는 영향은 5-bromo-2'-deoxyuridine(BrdU) 면역조직화학법(immunohistoc- hemistry)과 BDNF에 대한 western blot으로 규명하였다.
재료 및 방법
1. 실험동물
본 실험에 사용된 동물은 7주령의 Sprague-Dawley 계 암 컷 흰쥐(210±10 g)를 국가공인 동물취급업체(Orient Co., Chungbuk, Korea)로부터 공급받았다. 실험동물을 3일간의 환경적응 기간을 거치게 한 후, 대조군(N=10), 대조 트레 드밀 달리기군(N=10), 외상성 뇌손상군(N=10), 외상성 뇌
손상 후 트레드밀 달리기군(N=10)의 4군으로 분류하였다.
흰쥐들은 사육케이스(30×20 cm)에 넣고 항온(20±2oC), 항습 (60%)이 유지되며 12시간 간격으로 낮과 밤을 교대되는 동 일한 실험실 환경에서 사육하였다.
2. 외상성 뇌손상 유발
외상성 뇌손상 유발을 위하여 흰쥐들에게 Zoletil 50Ⓡ (10 mg/kg i.p.; Vibac, Carros, France)을 복강에 주입하여 마취한 후, 뇌정위수술 기구(stereotaxic frame; myNeurolab. Co., St.
Louis, MO, USA)를 이용하여 고정 하였다. 머리 부분의 털 을 면도하고 두피를 절개한 후 우측 측두근을 일부 박리하 여 외측으로 견인한 후 6 mm의 천두공(burr hole)을 정중선 (midline)에서 1.5 mm 떨어져 bregma와 lambda사이(천공공의 앞쪽 끝 부분이 bregma에서 2 mm 정도 뒤로 위치)에 전기 드릴을 이용하여 뚫었다. 이때 경막은 보존하였다. 이후 직 경 5 mm의 끝부분이 둥글게 처리된 stainless steel로 된 im- pactor tip을 경막이 약간 눌리게 한 점을 영점으로 하였고, 변형 깊이는 2.5 mm, 충격속도는 5.0 meters/sec, 변형지속 시간은 0.2 sec로 하여 뇌정위 충격 장치(myNeurolab. Co.)를 이용하여 외상성 뇌손상 모델을 유발하였다. 뇌에 충격을 가한 후 절개부위를 봉합하였으며, 외상성 뇌손상을 유발 시킨 후 1일간 안정을 취하였다. 외상성 뇌손상을 유발 시 키지 않은 대조군의 흰쥐들은 두피를 절개하고 구멍을 뚫 었으나 뇌 충격을 가하지 않았다(Kim BK et al., 2011).
3. 기억 능력 측정
기억능력은 passive avoidance task를 사용하였으며, 실험동 물들에게 최초 7×25 cm platform (2.5 cm high)에서 2분간 안정 을 취하게 한 후 기억 능력 측정을 실시하였다. 기록은 수술 후 14일째 platform위에 놓인 쥐들이 평행하게 놓인 1 cm 간 격의 스테인레스 강철막대기(42×25 cm)에 네 발이 모두 닿았 을 때 측정하였다. Platform에서 내려온 흰쥐들에게 0.3-mA의 전기 자극을 2초간 주어 쥐들에게 기억을 시킨 뒤 24시간 후 기록측정을 실시하였다. 기록측정은 동일한 절차로 하였으 며, 전기 자극은 부여하지 않았다. Platform에서 네발이 모두 닿는 시간은 최대 600초까지 기록하였다(Kim BK et al., 2011).
4. 운동 프로토콜과 BrdU 주사
운동 그룹은 수술 후 2일 후 1일 1회, 30분간 실시하였으 며, 운동의 강도는 경사도를 0%로 고정시킨 상태에서 운 동시작 처음 5분 동안 2 m/min, 다음 5분은 5 m/min, 마지
막 20분은 8 m/min의 점증적 강도로 연속 10일간의 운동으 로 구성하였다. BrdU (50 mg/kg in saline)는 처음 운동 시작 부터 5일간, 1일 1회 피하 내로 운동 30분 전에 주사하였다 (Lee HH et al., 2009).
5. 조직 처리
실험이 끝난 동물은 Zoletile 50Ⓡ (Vibac)으로 마취시킨 후 흉강을 열고 좌심실을 통하여 50 mM phosphate buffer saline (PBS)를 5분간 주입하였다. 계속해서 100 mM의 phosphate buffer (PB)에 녹인 4% paraformaldehyde (PFA)용액을 4oC에서 10분간 관류시킨 후 뇌를 적출하고, 4oC에서 4% PFA용액 에 24시간 침전시킨 후 고정하였다. 고정된 뇌조직을 30%
sucrose 용액에 일주일간 침전시킨 후 freezing microtome (Leica, Nussloch, Germany)으로 40 μm의 두께로 연속관상 절편을 제작하였다.
6. BrdU 면역조직화학법
BrdU 면역조직화학법을 이용하여 해마 치상회에서 새로 생성된 세포를 측정하였다(Lee HH et al., 2009). 각 그룹에서 각각 최소 12장의 뇌 절편을 선택하여 50 mM PBS에서 3분 씩 3번 세척한 후, 0.5% Triton X-100으로 20분간 incubation 시켰다. 그 다음 50 mM PBS에서 3분씩 3번 세척한 후, 50%
formamide-2×standard saline citrate (SSC) 용액에 조직을 넣고 65oC의 수조에서 2시간 동안 shaking incubation 시키며, 다시 2×SSC 용액으로 5분간 2번 세척한 후, 2 N HCl에 조직을 옮 겨 37oC에서 30분간 incubation 시켰다. 계속해서 100 mM sodium borate (pH 8.5)로 25oC에서 10분 이상 중화시키고 50 mM PBS에서 3분씩 3번 세척한 후 1시간 동안 1% BSA와 10%의 horse serum 그리고 3% Triton X-100으로 blocking을 실시하였다. Blocking 후 primary antibody인 anti-BrdU Ab (1:600; Roche, Mannheim, Germany)로 24시간 incubation시키 고, 50 mM PBS에서 3분씩 3번 세척한 후 실온에서 1시간 동안 secondary antibody (1:200; Vector Laboratories, Burlingame, CA, USA)로 반응시켰다. 다시 50 mM PBS에서 3분씩 3번 세 척한 후 1시간 동안 avidin-biotin-horseadish peroxidase complex (ABC) 용액으로 처치하였다. 50 mM PBS에서 3분씩 3번 세 척한 다음 조직을 50 mM Tris-HCl (pH 7.6) 용액에 0.02%
3,3’-diaminobenzidine containing nickel chloride (40 mg/ml, nickel-DAB)과 0.03% 과산화수소를 넣어 반응시켰다.
BrdU 면역조직화학법을 실시한 후 신경전구세포에서 신 경세포로 분화되는 신경세포 생성을 알아보기 위하여
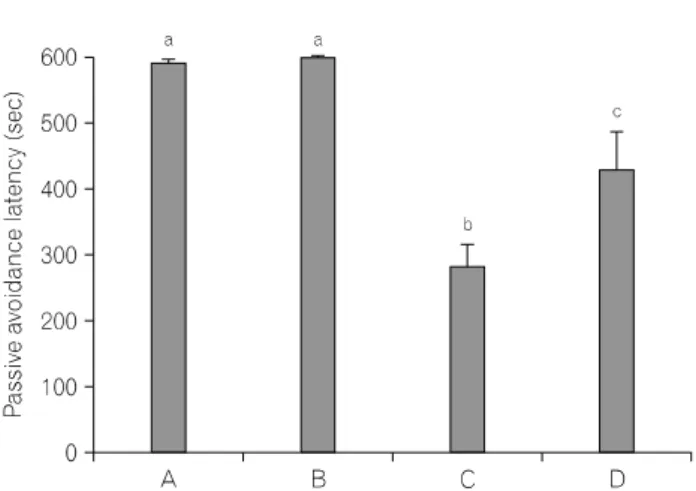
Fig. 1. Effect of treadmill exercise on the latency in the step-down
avoidance task. (A) sham-operation group, (B) sham-operation and tread- mill exercise group, (C) traumatic brain injury (TBI)-induction group, (D) TBI-induction and treadmill exercise group. Values are represented as the mean±standard error of the mean (SEM). Different letters (a∼c) denote statistically significant differences (p<0.05) after Duncan post-hoc, i.e., groups marked with different letters differ statistically.blocking 후 primary antibody인 anti-neuronal nucleic (NeuN) 항 체(1:300; Chemicon International, Temecula, CA, USA)로 24시 간 incubation시키고, 50 mM PBS에서 3분씩 3번 세척한 후 실온에서 1시간 동안 secondary antibody (1:200; Vector Labo- ratories, Burlingame, CA, USA)로 반응시켰다. 다시 50 mM PBS에서 3분씩 세척한 후 1시간 동안 ABC 용액으로 처치 하였다. 50 mM PBS에서 3분씩 3번 세척한 다음 조직을 DAB 발색용액(pH 7.6)으로 반응시킨 후 gelatin-coating된 슬 라이드에 놓고 탈수시킨 후 고정하였다.
7. BDNF 단백질 발현의 측정
해마의 BDNF 단백질 발현을 western blot으로 측정하였 다(Lee HH et al., 2009). 단백질을 PBS로 washing 후 extraction buffer로 세포를 lysis 시켰다. 10분간 ice에 방치 후 4oC에서 12,000×g로 15분간 원심 분리하였다. 상청액을 새로운 tube 에 옮겨 단백질을 정량 후 사용하였다. 단백질의 정량은 BioRad colorimetric protein assay kit (BioRad, Hercules, CA, USA)를 사용하였으며, 분리된 단백질을 SDS-polyacrylamide gel에서 전기 영동하였다. Sample과 4배의 sample buffer를 넣 은 뒤 100oC에서 1분간 heating 한 후 sample을 well에 넣은 뒤 100 V로 1시간 정도 run 시켰다. Coomassic blue로 5분간 gel을 stain한 후 destaining solution으로 washing하고 electro- phoresis된 gel을 membrane으로 이전하였다. Blocking solution 에서 천천히 흔들어 세척한 후, 보합결합을 위한 주머니에 흡착지를 넣고 2∼4시간 1차 항체와 결합시키고, 깨끗이 세척한 다음 이차 항체-효소와 다시 1시간 동안 결합시킨 후 발색 용액을 넣고 film에 발색시켰다.
8. BrdU-양성세포수 측정
각각의 항체들과 면역 반응이 일어난 세포들은 광학현 미경(BX-51, Olympus, Tokyo, Japan)을 이용하여 관찰하였으 며, BrdU-양성 세포 수를 mm2당으로 정량적인 분석을 실 시하였다.
9. 자료처리
본 실험에서 얻은 자료는 SPSS (version 12.0) 통계프로그 램을 이용하여 처리하였다. 측정치는 평균±표준편차로 표 시하였고, 그룹 간 비교를 위하여 one-way ANOVA와 Duncan's post-hoc test를 실시하였다. 통계적 유의수준은 p
<0.05로 설정하였다.
결 과
1. 외상의 정도가 그룹별에 같음을 보여주는 결과 외상성 뇌손상 정도가 외상성 뇌손상 그룹과 외상성 뇌 손상 후 트레드밀 달리군의 같음을 알아보기 위하여 외 상성 뇌손상 유발 후 하루 휴식 후 각 그룹 당 3마리씩 희생을 하여 세포사멸을 알아 볼 수 있는 caspase-3를 면역 화학조직법을 통하여 유발정도를 알아보았다. 외상성 뇌 손상군은 255.83±10.03/mm2, 그리고 외상성 뇌손상 후 트 레드밀 달리기군은 265.67±9.01/mm2이었다. 운동을 하기 전두 그룹의 외상의 정도는 차이가 없었다(p<0.05, F=
0.53).
2. 트레드밀 달리기 운동이 단기기억에 미치는 영향 기억능력을 측정하기 위하여 passive avoidance task를 실시 하였다(Fig. 1). 대조군은 589.22±7.42 sec, 대조 트레드밀 달 리기군은 597.70±2.30 sec, 외상성 뇌손상군은 283.67±31.15 sec, 그리고 외상성 뇌손상 후 트레드밀 달리기군은 427.00±
55.83 sec이었다. 외상성 뇌손상군은 대조군과 비교해 단기 기억이 유의하게 감소되었으며(p<0.05), 외상성 뇌손상 후 트레드밀 운동은 단기기억을 유의하게 향상시킨 것으로 나타났다(p<0.05, F=19.81).
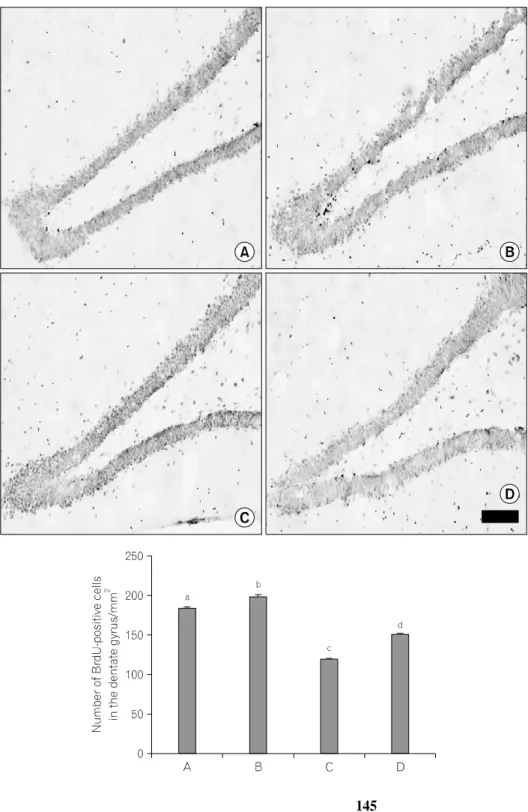
Fig. 2. Effect of treadmill exercise on
the cell proliferation in the hippocampal dentate gyrus. (A) sham-operation gro- up, (B) sham-operation and treadmill exercise group, (C) traumatic brain in- jury (TBI)-induction group, (D) TBI-in- duction and treadmill exercise group.Values are represented as the mean±
standard error of the mean (SEM).
Different letters (a∼d) denote statisti- cally significant differences (p<0.05) after Duncan post-hoc, i.e., groups mar- ked with different letters differ statisti- cally. The scale bar represents 50 μm.
3. 트레드밀 달리기 운동이 해마 신경세포생성에 미 치는 영향
해마 치상회 부위에서의 신경세포의 생성을 알 수 있는 BrdU immunohistochemistry를 실시하였다(Fig. 2). BrdU-양성 세포 수는 대조군은 184.50±1.44/mm2, 대조 트레드밀 달리
군은 198.90±1.94/mm2, 외상성 뇌손상군은 120.08±0.72/mm2, 그리고 외상성 뇌손상 후 트레드밀 달리기군은 150.40±
0.96/mm2이었다. 외상성 뇌손상군은 대조군과 비교해 해마 치상회 부위에서 BrdU-양성 세포 수가 유의하게 감소되었 으며(p<0.05), 외상성 뇌손상 후 트레드밀 운동은 BrdU-양 성 세포 수를 유의하게 증가시키는 것으로 나타났다(p
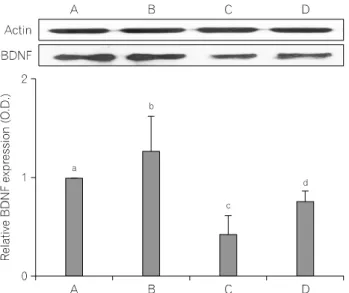
Fig. 3. Effect of treadmill exercise on the brain-derived neurotrophic factor
(BDNF) expression in the hippocampus. Upper: Representative expressions of the BDNF and actin in the hippocampus. Lower: The density of BDNF expression in the hippocampus. Molecular weights of BDNF=15 kDa;Actin=42 kDa. (A) sham-operation group, (B) sham-operation and treadmill exercise group, (C) traumatic brain injury (TBI)-induction group, (D) TBI-induction and treadmill exercise group. Values are represented as the mean±standard error of the mean (SEM). Different letters (a∼d) denote statistically significant differences (p<0.05) after Duncan post-hoc, i.e., groups marked with different letters differ statistically.
<0.05, F=830.55).
4. 트레드밀 달리기 운동이 해마 BDNF의 발현에 미치 는 영향
해마에서 BDNF 단백질 발현을 western blot으로 분석하 였다(Fig. 3). BDNF 단백질은 대조군을 1.00±0.00%으로 하 였을 때, 대조 트레드밀 달리기군은 1.27±0.35%, 외상성 뇌 손상군은 0.43±0.19%, 그리고 외상성 뇌손상 후 트레드밀 달리기군은 0.75±0.11%이었다. 외상성 뇌손상군은 대조군 과 비교해 해마치상회 부위에서 BDNF의 발현이 유의하게 감소되었으며(p<0.05), 외상성 뇌손상 후 트레드밀 운동은 BDNF의 발현을 유의하게 증가시키는 것으로 나타났다(p
<0.05, F=111.66).
고 찰
외상성 뇌손상은 외부 충격에 의해 뇌가 손상되는 경우 를 의미하는데, 이로 인하여 의식상태의 저하 또는 소실이 나타난다(Flanagan, 2006). 외상성 뇌손상은 그 후유증이 다
양하고, 만성화되는 경우가 많아서 치료가 쉽지 않으며, 사 회ㆍ경제적인 비용도 매우 많이 소요된다. 또한 뇌손상의 후유증으로 고통 받고 있는 환자들의 수가 급격히 증가되 고 있다(Flanagan, 2006). 외상성 뇌손상으로 인한 일차적인 후유장애로는 의식의 변화와 함께 인지장애, 신체장애, 성 격의 변화와 퇴행(Borgaro et al., 2003), 정서장애와 사회적 적응장애(Kim BK et al., 2011) 등이 나타날 수 있다. 그 중 에서도 특히 인지적 결함(cognitive deficits)은 다른 기능성 장애보다 사회적 적응이나 직업으로의 복귀에 많은 영향 을 미치기 때문에(Borgaro et al., 2003), 외상성 뇌손상 환자 의 재활에서 다루어야 될 중요한 문제 중 하나이다. 이러 한 외상성 뇌손상의 이차상해에서 일어나는 생화학적 변 화는 염증반응을 유발하고 세포사를 일으킨다. 또한 회복 을 위한 성장과 관련된 단백질의 발현도 나타난다. 많은 연구에서 외상성 뇌손상은 학습능력과 기억형성에 손상을 유도하며, 뇌기능의 소실로 인한 새로운 정보의 인지과정 을 방해한다고 보고하였다(Cohen et al., 2007; Lee HH et al., 2009; Kim BK et al., 2011). 본 실험결과에서도 외상성 뇌손 상을 유발한 흰쥐에서 passive avoidance task에서 latency가 감소하여, 외상성 뇌손상에 의하여 단기기억이 현저히 저 하되었음을 보여주었다(Fig. 1).
운동의 기억력을 향상시키는 효과는 많은 연구에서 밝 혀졌다(Carro et al., 2001; Lee HH et al., 2009). 운동의 기억력 향진 작용은 해마에서 신경세포 생성을 증가시키고 BDNF, basic fibroblast growth factor (bFGF)와 같은 신경성장인자들 의 발현을 증가시켜 일어난다(Griesbach et al., 2008; Kim BK
et al., 2011). 본 실험의 결과 외상성 뇌손상에 의하여 감소
한 passive avoidance task의 latency가 트레드밀 운동에 의하 여 증가되어 외상성 뇌손상에 의하여 저하된 단기기억이 트레드밀 운동에 의하여 향상되었음을 나타낸다(Fig. 1).신경세포생성(neurogenesis)은 신경세포가 증식되고 생존 하여 분화하는 과정을 거치게 된다. 성숙한 쥐에서 신경전 구세포는 해마의 치상회의 과립세포하 영역에서 생성되어 과립층으로 지속적으로 이동하여 신경세포로 분화되고, 축삭을 내어 다른 신경세포와 시냅스를 이루어 성숙한 신 경세포를 형성한다(Liu et al., 1998; Mattson, 2000). 새로운 신경세포가 생성되는 곳으로는 측뇌실 전측의 뇌실하 영 역과 해마 치상회의 과립세포하 영역으로 알려져 있다 (Parent et al., 2002). 해마는 학습과 기억의 형성과정에 관여 하는 중요한 뇌 부위로서 신경발생 뿐 아니라 성장 후에도 신경세포생성이 활발하게 일어나는 부위이다. 따라서 뇌
영역 가운데 해마는 신경가소성이 가장 큰 부위이며(Fuchs
et al., 2000; Lie et al., 2004), 외상성 뇌손상 후 가장 치명적
인 영향을 받는 뇌 영역으로 알려져 있다(Clausen et al., 2005; Scheff et al., 2005). 본 실험결과 외상성 뇌손상에 의하 여 해마에서의 신경세포 생성이 억제되었다(Fig. 2).운동은 해마에서 신경전구세포의 증식유도를 통한 신경 세포생성을 가장 효율적으로 증가시킬 수 있는 인자로 알 려져 있다(Trejo et al., 2001). 해마에서 생성된 신경세포는 학습 및 기억능력 향성에 중요한 역할을 담당하게 되며, 운동으로 해마의 신경세포생성이 증가되면 학습 및 기억 능력이 개선됨이 보고되었다(Vaynman et al., 2004). 운동은 외상성 뇌손상 후에도 해마의 신경세포생성을 증가시켜 기억력을 향상시키는 것으로 알려져 있다(Lee HH et al., 2009; Kim BK et al., 2011). 운동은 뇌혈관의 형성 및 모세혈 관의 성장을 촉진한다(Isaacs et al., 1992; Blair et al., 1995). 또 한 운동은 신경세포 돌기의 수와 시냅스 연결을 증가시켜 (Trejo et al., 2001) 뇌의 장기강화(long-term potentiation: LTP) 를 증폭시킴으로써 뇌의 기능을 향상시킨다(Blomquist et
al., 1987). 본 실험결과에서 트레드밀 운동은 외상성 뇌손
상에 의하여 감소된 신경세포 생성을 증가시켰다(Fig. 2).BDNF는 해마 신경세포생성의 조절에 중요한 역할을 담 당하는 것으로도 알려져 있다(Donovan et al., 2008). 또한 해 마에서 BDNF는 신경세포생성과 시냅스 가소성을 향상시 켜 인지기능을 강화하는 물질로 알려져 있는데, 외상성 뇌 손상을 지닌 해마에서는 신경세포생성을 감소시키며, BDNF 단백질 발현을 감소시킨다고 보고되었다(Wagner et al., 2005; Lee HH et al., 2009; Kim BK et al., 2011). 본 실험결과 에서도 외상성 뇌손상을 유발한 흰쥐의 해마에서 BDNF 단백질 발현이 감소되었음을 확인하였다(Fig. 3).
운동은 BDNF의 발현을 증가시키는데, 운동이 신경세포 생성을 촉진시키는 작용은 BDNF의 작용을 매개로 일어난 다고 알려져 있다(Carro et al., 2001). 외상성 뇌손상 후 운동 에 의한 기능의 회복은 BDNF와 같은 신경성장인자들의 발현을 촉진하는 운동의 작용과 밀접한 관련이 있다(Bar- nabé-Heider et al., 2003; Ke et al., 2011). Griesbach et al.(2004) 은 신경 손상 후 회복 시 그리고 운동학습이나 신체활동 및 기억력 강화 시에도 BDNF 발현이 증가된다고 보고하 였다. 운동과 관련해서는 외상성 뇌손상 후 운동은 BDNF 와 synapsin I을 증가시켜 신경가소성을 향상시켰다고 보고 하였다(Griesbach et al., 2008; Kim BK et al., 2011). 본 실험결 과에서 트레드밀 운동은 외상성 뇌손상에 의하여 감소된
BDNF 발현을 증가시켰다(Fig. 3).
임상적으로는 지구성 트레이닝과 심폐조절 운동이 외상 성 뇌손상 환자의 인지기능 재활과정에 상당한 이점이 있 는 것으로 평가되고 있으며(Mossberg et al., 2010), 일부 연구 에서 운동은 외상성 뇌손상 환자를 위한 권고지침으로 제 시되고 있다(Weightman et al., 2010). 본 실험의 결과 트레드 밀 달리기 운동은 외상성 뇌손상 흰쥐에서 신경세포의 생 성과 BDNF의 발현을 증가시킴으로써 단기기억의 감퇴를 경감시키는 효과가 있음을 알 수 있다. 본 연구를 통하여 운동은 외상성 뇌손상과 같은 퇴행성 뇌질환의 재활에 도 움이 되는 효과적인 치료로 활용될 수 있다고 생각되어진 다.
참 고 문 헌
Almli CR, Levy TJ, Han BH et al. (2000) BDNF protects against spatial memory deficits following neonatal hypoxia-ischemia. Exp.
Neurol. 166:99-114.
Barnabé-Heider F, Miller FD (2003) Endogenously produced neurot- rophins regulate survival and differentiation of cortical progeni- tors via distinct signaling pathways. J. Neurosci. 23:5149-5160.
Blair SN, Kohl HW, Barlow CE et al. (1995) Changes in physical fitness and all-cause mortality. A prospective study of healthy and unhealthy men. J. Am. Med. Assoc. 273:1093-1098.
Borgaro SR, Prigatano GP, Kwasnica C et al. (2003) Cognitive and affective sequelae in complicated and uncomplicated mild traumatic brain injury. Brain Inj. 17:189-198.
Carro E, Trejo JL, Busiguina S et al. (2001) Circulating insulin-like growth factor I mediates the protective effects of physical exercise against brain insults of different etiology and anatomy.
J. Neurosci. 21:5678-5684.
Clausen F, Lewen A, Marklund N et al. (2005) Correlation of hippocampal morphological changes and morris water maze performance after cortical contusion injury in rats. Neurosurgery 57:154-163.
Cohen AS, Pfister BJ, Schwarzbach E (2007) Injury-induced altera- tions in CNS electrophysiology. Prog. Brain Res. 161:143-169.
Donovan MH, Yamaguchi M, Eisch AJ (2008) Dynamic expression of TrkB receptor protein on proliferating and maturing cells in the adult mouse dentate gyrus. Hippocampus 18:435-439.
Flanagan SR, Hibbard MR, Riordan B et al. (2006) Traumatic brain injury in the elderly: diagnostic and treatment challenges. Clin.
Geriatr. Med. 22:449-468.
Fuchs E, Gould E (2000) In vivo neurogenesis in the adult brain:
regulation and functional implications. Eur. J. Neurosci. 12:2211-
2214.
Griesbach GS, Hovda DA, Gomez-Pinilla FS et al. (2008) Voluntary exercise or amphetamine treatment, but not the combination, increases hippocampal brain-derived neurotrophic factor and synapsin I following cortical contusion injury in rats. Neuroscience 154:530-540.
Griesbach GS, Hovda DA, Molteni R et al. (2004) Voluntary exercise following traumatic brain injury: brain-derived neurotrophic factor upregulation and recovery of function. Neuroscience 125:
129-139.
Guezennec CY, Abdelmalki A, Serrurier B et al. (1998) Effects of prolonged exercise on brain ammonia and amino acids. Int. J.
Sports Med. 19:323-327.
Hall PG, Sullivan TR, Gibson KM et al. (2005) Spatial and temporal characteristics of neurodegeneration after controlled cortical impact in mice: more than a focal brain injury. J. Neurotrauma 22:252-265.
Hicks RR, Li C, Zhang L et al. (1999) Alterations in BDNF and trkB mRNA levels in the cerebral cortex following experimental brain trauma in rats. J. Neurotrauma 16:501-510.
Isaacs KR, Anderson BJ, Alcantara AA et al. (1992) Exercise and the rain: angiogenesis in the adult rat cerebellum after vigorous physical activity and motor skill learning. J. Cereb. Blood Flow Metab. 12:110-119.
Junque C, Ramirez-Ruiz B, Tolosa E et al. (2005) Amygdalar and hippocampal MRI volumetric reductions in Parkinson's disease with dementia. Mov. Disord. 20:540-544.
Ke Z, Yip SP, Li L et al. (2011) The effects of voluntary, involun- tary, and forced exercises on brain-derived neurotrophic factor and motor function recovery: a rat brain ischemia model. PLoS One 6:16643.
Kim BK, Sung YH, Kim SE et al. (2011) Swimming alleviates traumatic brain injury-induced memory impairment by enhancing of brain-derived neurotrophic factor in the hippocampus of rats.
Korean J. Str. Res. 19:205-212.
Lee HH, Yoon JH, Jeong IG et al. (2009) Effects of exercise intensity on hippocampal cell proliferation and BDNF expression in intracerebroventricular streptozotocin induced-memory impair- ment in rats. Exer. Sci. 18:475-488.
Lie DC, Song H, Colamarino SA et al. (2004) Neurogenesis in the adult brain: new strategies for central nervous system diseases.
Annu. Rev. Pharmacol. Toxicol. 44:399-421.
Liu J, Solway K, Messing RO et al. (1998) Increased neurogenesis in
the dentate gyrus after transient global ischemia in gerbils. J.
Neurosci. 18:7768-7778.
Lopez-Toledano MA, Shelanski ML (2004) Neurogenic effect of beta-amyloid peptide in the development of neural stem cells. J.
Neurosci. 24:5439-5444.
Mattson MP (2000) Neuroprotective signaling and the aging brain:
take away my food and let me run. Brain Res. 886:47-53.
Mossberg KA, Amonette WE, Masel BE (2010) Endurance training and cardiorespiratory conditioning after traumatic brain injury. J.
Head Trauma Rehabil. 25:173-183.
Neeper SA, Gomez-pinilla F, Choi J et al. (1996) Physical activity increases mRNA for brain-derived neurotrophic factor and nerve growth factor in rat brain. Brain Res. 726:49-56.
Parent JM, Valentin VV, Lowenstein DH (2002) Prolonged seizures increase proliferating neuroblasts in the adult rat subventricular zone–olfactory bulb pathway. J. Neurosci. 22:3174-3188.
Ponsford J, Draper K, Schönberger M (2008) Functional outcome 10 years after traumatic brain injury: its relationship with demographic, injury severity, and cognitive and emotional status.
J. Int. Neuropsychol. Soc. 14:233-242.
Rudehill S, Muhallab S, Wennersten A et al. (2006) Autoreactive antibodies against neurons and basal lamina found in serum following experimental brain contusion in rats. Acta Neurochir.
148:199-205.
Scheff SW, Price DA, Hicks RR (2005) Synaptogenesis in the hippocampal CA1 field following traumatic brain injury. J.
Neurotrauma 22:719-732.
Trejo JL, Carro E, Torres-Aleman I (2001) Circulating insulin-like growth factor I mediates exercise-induced increases in the number of new neurons in the adult hippocampus. J. Neurosci.
21:1628-1634.
Vaynman S, Ying Z, Gomez-Pinilla F (2004) Hippocampal BDNF mediates the efficacy of exercise on synaptic plasticity and cognition. Eur. J. Neurosci. 20:2580-2590.
Wagner AK, Chen X, Kline AE et al. (2005) Gender and environ- mental enrichment impact dopamine transporter expression after experimental traumatic brain injury. Exp. Neurol. 195:475-483.
Weicker H, Struder HK (2001) Influence of exercise on serotonergic neuromodulation in the brain. Amino Acids 20:35-47.
Weightman MM, Bolgla R, McCulloch KL (2010) Physical therapy
recommendations for service members with mild traumatic brain
injury. J. Head Trauma Rehabil. 25:206-218.
= 국문초록 =
외상성 뇌손상은 많은 신경정신과적 이상을 유발시키며, 특히 기억 상실은 외상성 뇌손상의 많은 증상 중 하나이다.
외상성 뇌 손상에 대한 뚜렷한 치료법이 아직 없는 반면, 운동이 신경세포의 보호 작용이 있음은 여러 연구들에 의하여 잘 알려져 있다. 본 연구에서는 흰쥐를 이용하여 외상성 뇌 손상을 유발시켰고, 트레드밀 달리기가 외상성 뇌 손상에 미치는 영향을 연구하였다. 외상성 뇌 손상은 기억력의 감퇴를 초래하였고, 해마 치상회에서 신경세포생 성과 해마에서의 뇌유래 신경영양인자(brain-derived neurotrophic factor)의 발현을 감소시켰다. 트레드밀 달리기 운동은 해마 치상회에서 신경세포생성과 해마에서 뇌 유래 영양인자발현을 증가시켰으며, 외상성 뇌손상에 의한 기억력의 감퇴를 경감시켰다. 본 연구를 통하여 외상성 뇌 손상에 의한 신경계 후유증에 트레드밀 달리기 운동이 효과가 있음 을 알 수 있었다.
중심단어: 외상성 뇌손상, 트레드밀 달리기, 단기기억, 뇌유래 신경영양인자, 흰쥐