Corresponding author: Sang-Won Lee, Department of Medicinal Crop Research, National Institute of Horticultural & Herbal Science, Rural Development Administration, 80, Bisan-li, Soi-myun, Eumseong 369-873, Korea
Tel: +82-43-871-5611, E-mail: [email protected] Received August 15, 2011, Revised September 1, 2011 Accepted September 20, 2011
This work was carried out with the support of “Development of Antiobesity Agent from Herbal Medicine (PJ007415042011)” Rural Development Administration, Korea.
A Promoter Polymorphism (rs3737965, -336 C/T) of Methylenetetrahydrofolate Reductase (MTHFR) Is
Associated with Obesity in Korean Population
Department of Medicinal Crop Research, National Institute of Horticultural & Herbal Science, Rural Development Administration, Eumseong, Korea
Young-Ock Kim, Seung-Yu Kim, Sang-Won Lee
Methylenetetrahydrofolate reductase (MTHFR) is a major enzyme in the metabolism of folate. Several studies have shown the relationship between the MTHFR gene and obesity. In this study, we investigated whether single nucleotide polymorphisms (SNPs) of the MTHFR gene were associated with obesity in Korean population. One hundred twenty two overweight/obese and 125 control subjects were recruited. Two SNPs (rs3737965, -336 C/T and rs1801131, Glu429Ala) of the MTHFR gene were selected, and genotypes of these two SNPs were determined using direct sequencing. SNPStats, SPSS 18.0, and Haploview 4.2 softwares were used to analyze genetic data. Multiple logistic regression models (codominant1, codominant2, dominant, recessive, overdominant, and log-additive) were conducted for odds ratio (OR), 95% confidence interval (CI), and p value. For the defined results, p values were recalculated by Bonferroni correction (p
c). After Bonferroni correction, allele distributions of one promoter SNP (rs3737965) showed significant association with overweight/obese (OR=2.27, 95% CI=1.14∼4.51, p
c=0.038). The T allele frequency of rs3737965 in the overweight/obese group (11.1%) was higher than that in the control group (5.2%). These results indicate that the MTHFR gene may be associated with obesity in Korean population. (Korean J Str Res 2011;19:237∼242)
Key Words: Methylenetetrahydrofolate reductase, Obesity, Overweight, Single nucleotide polymorphism
INTRODUCTION
Obesity is one of serious medical problems and may be
implicated in the pathogenesis of diabetes (Barth, 2011), heart diseases (Patabiraman et al., 2011), and cancers (Sánchez-Lara et al., 2010). Obesity is caused by a combination of genetic and environmental factors including excessive food energy intake and lack of physical activity (Ravussin et al., 2000). In a genome-wide association study, rs12970134 located near the melanocortin 4 receptor (MC4R) gene was associated with waist circumference (Chambers et al., 2008). They concluded that genetic variation near MC4R was associated with a risk of adiposity and insulin resistance. Calton et al.(2009) reported that the total prevalence of rare MC4R variants in the severely obese adults was 2.25%
compared with 0.64% in the lean controls (p<0.005). The fat
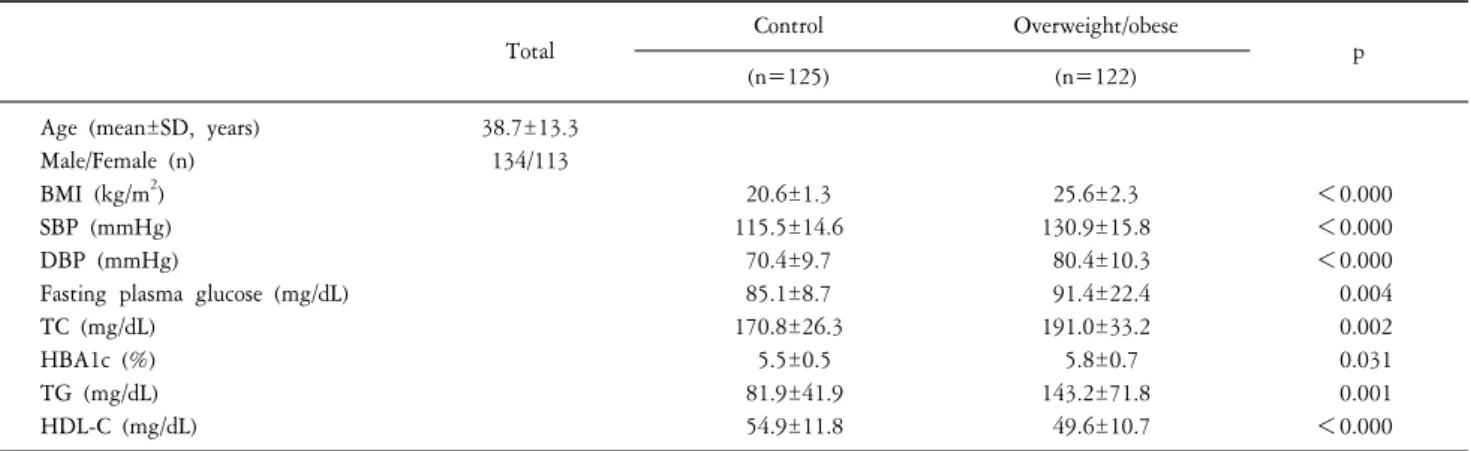
Table 1. Clinical and biochemical characteristics in overweight/obese and control subjects.
Total Control Overweight/obese
p
(n=125) (n=122)
Age (mean±SD, years) Male/Female (n) BMI (kg/m2) SBP (mmHg) DBP (mmHg)
Fasting plasma glucose (mg/dL) TC (mg/dL)
HBA1c (%) TG (mg/dL) HDL-C (mg/dL)
38.7±13.3 134/113
20.6±1.3 115.5±14.6
70.4±9.7 85.1±8.7 170.8±26.3
5.5±0.5 81.9±41.9 54.9±11.8
25.6±2.3 130.9±15.8 80.4±10.3 91.4±22.4 191.0±33.2 5.8±0.7 143.2±71.8 49.6±10.7
<0.000
<0.000
<0.000 0.004 0.002 0.031 0.001
<0.000 Data are mean±standard deviation (SD). n: number of subjects, BMI: body mass index, SBP: systolic blood pressure, DBP: diastolic blood pressure, TC:
total cholesterol, HBA1c: hemoglobin A1c, TG: triglyceride, HDL-C: high-density lipoprotein cholesterol.
mass and obesity associated (FTO) gene had shown to be a risk factor for obesity in Caucasian (Grant et al., 2008), Asian (Hotta et al., 2008), and African American (Grant et al., 2008). Sun et al.(2010) revealed that a FTO polymorphism (rs9939609) was associated with obesity in Chinese population. Jermendy et al.(2011) reported that the association between peroxisome proli- ferator-activated receptor gamma (PPARG) Prol2Ala polymorphism and glucose metabolism was already present in children and adolescents with obesity who might be at the very beginning of the natural course of type 2 diabetes. Pereira et al.(2011) demonstrated that the leptin receptor (LEPR) and the adrenergic, beta-2, receptor, surface (ADRB2) genes had synergistic effects on the risk of obesity in Japanese.
Methylenetetrahydrofolate reductase (MTHFR) plays a major role in folate metabolism (Guenther et al., 1999). MTHFR catalyzes the conversion of 5,10-methylenetetrahydrofolate to 5-methyltetrahydrofolate, a co-substrate for homocysteine reme- thylation to methionine. Previously, several studies have been demonstrated the associations between single nucleotide polymor- phisms (SNPs) of the MTHFR gene and obesity. Frelut et al.(2006) reported that folate intake and the MTHFR SNP represented a part of the link between antioxidant status and liver disease in obese adolescent girls. Lambrinoudaki et al.(2008) found the MTHFR Ala222Val SNP was associated with increased androgenicity, elevated body mass index (BMI), and increased waist-to-hip ratio (WHR) in healthy postmenopausal women.
In this study, we investigated whether SNPs of the MTHFR
gene are associated with obesity in Korean population.
MATERIALS AND METHODS
1. Subjects
BMI was measured by dividing the subject's mass by the square of his or her height (kg/m
2). According to the classification of Korean Society for the Study of Obesity (underweight, BMI
<18; normal, BMI 18 to <23; moderately obese, BMI 23 to
<25; obesity I, BMI 25 to <30; obesity II, BMI≥30), we divided into two groups (overweight/obese, BMI≥23, n=122;
normal control, BMI 18 to <23, n=125). Participants with severe diseases, such as cancers, stoke, and psychiatric disorders were excluded. Our study was conducted according to the Declaration of Helsinki guidelines and approved by the Ethics Review Committee of the Medical Research Institute, School of Medicine, Kyung Hee University. Written informed consent was obtained to each subject. All subjects were measured the following clinical and biochemical characteristics: age, BMI, systolic blood pressure (SBP), diastolic blood pressure (DBP), fasting plasma glucose, total cholesterol (TC), hemoglobin A1c (HBA1c), triglyceride (TG), and high-density lipoprotein cholesterol (HDL-C) (Table 1).
2. SNP selection and genotyping
In recent, Kim SK et al.(2010) investigated the relationship
between ten MTHFR SNPs [rs1537514 (3’untranslated region,
3’UTR), rs11559040 (3’UTR), and rs1801133 (Ala222Val) in exon region; rs3818762, rs12121543, rs1994798, rs6541003, rs4846052, rs17037396, and rs17367504 in intron region] and obesity. In this study, we selected one promoter SNP (rs3737965, -336 C/T) and one missense SNP (rs1801131, Glu429Ala) in the MTHFR gene. The heterozygosity of rs3737965 and rs1801131 was 0.133 and 0.489, respectively (http://www.ncbi. nlm.nih.gov/SNP, BUILD 132). DNAs were extracted using DNA isolation kit for blood (Roche, Indianapolis, IN, USA). Genotyping of each SNP was determined by direct sequencing. For the polymerase chain reaction (PCR), we used the following primers: for rs3737965 (sense primer, 5’-GAAACATGGGTGGGATTTACTG-3’; antisense primer, 5’-AGAGTGGCAGTAAAGGAGGAAG-3’; product size, 356 bp) and for rs1801131 (sense primer, 5’-GAGTCAGGGG- CAGAATTTACAG-3’; antisense primer, 5’-ACAGGATGGGG- AAGTCACAG-3’; product size, 376 bp). PCR products were sequenced by an ABI PRISM 3730XL analyzer (PE Applied Biosystems, Foster City, CA, USA) and sequencing data were analyzed using SeqManII software v2.3 (DNASTAR Inc., Madison, WI, USA).
3. Statistical analysis
SNPStats (http://bioinfo.iconcologia.net/index.php) and SPSS 18.0 (SPSS Inc., Chicago, IL, USA) were used to analyze genetic data. Multiple logistic regression models (codominant1, codomi- nant2, dominant, recessive, overdominant, and log-additive) were conducted for odds ratio (OR), 95% confidence interval (CI), and p value, adjusting for age and gender as covariables. To obtain the defined results, p values were recalculated by Bonferroni correction. A linkage disequilibrium (LD) block was estimated using Haploview 4.2 (Daly Lab Inc., Cambridge, MA, USA). The clinical and biochemical characteristics in overweight/obese and control subjects were analyzed using SPSS 18.0. The statistical significance was set at p<0.05.
RESULTS
Table 1 shows the clinical and biochemical characteristics in overweight/obese and control subjects. The total numbers of males/females were 134/113 (38.7±13.3 years, mean±SD). All participants were divided into two groups according to BMI
values (the overweight/obese group, BMI≥23, n=122; the control group, BMI<23, n=125). The mean of BMI was significant difference between the overweight/obese group and the control group (p<0.001). The values of SBP, DBP, fasting plasma glucose, TC, HBA1c, and TG in overweight/obese subjects were higher than those in control subjects (p<0.05). The value of HDL-C in the overweight/obese group was lower than that in the control group (p<0.000).
In this study, we evaluated whether MTHFR SNPs (rs3737965, -336 C/T; rs1801131, Glu429Ala) were contributed to obesity.
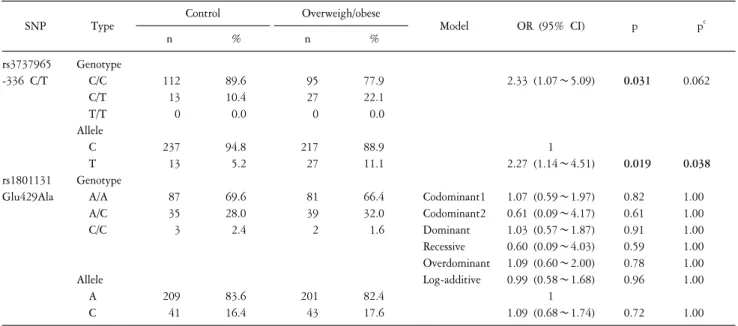
Genotype frequencies of these two SNPs were Hardy-Weinberg equilibrium in both groups (rs3737965, p=1.00 in the control group, p=0.36 in the overweight/obese group; rs1801131, p=0.47 in the control group, p=1.00 in the overweight/obese group). Genotype and allele frequencies of two tested SNPs in overweight/obese and control subjects are shown in Table 2.
Because there was no minor genotype (T/T) of rs3737965 in the overweight/obese and control groups, rs3737965 SNP was not applied to the multiple logistic regression models (codominant1, codominant2, dominant, recessive, overdominant, and log-additive models), and we simply analyzed the logistic regression (C/C versus C/T). Genotype distributions of rs3737965 had significant difference between two groups (OR=2.33, 95% CI=1.07∼5.09, p=0.031). The C/T genotype frequencies of rs3737965 in the control group and overweight/obese group were 10.4% and 22.1%, respectively. However, above association was disappeared after Bonferroni correction (p
c=0.062). In allele distributions of rs3737965, we also found significant association (OR=2.27, 95%
CI=1.14∼4.51, p=0.019). The significant association was remained after Bonferroni correction (p
c=0.038). A missense SNP rs1801131 did not show any significant differences in genotype and allele distributions (Table 2).
LD and haplotypes of rs3737965 and rs1801131 were condu- cted using Haploview 4.2 program. The LD block was construc- ted (Fig. 1,│D'│=1.0 and r
2=0.28). In haplotype analysis, there were three haplotypes (CA, frequency=0.82; CC, frequency=0.10;
TC, frequency=0.07). TC haplotype was weakly associated with
overweight/obese (p=0.05), but CA and CC haplotypes not
(p>0.05) (Table 3).
Table 2. Genotype and allele distributions of MTHFR SNPs in overweight/obese and control subjects.
SNP Type Control Overweigh/obese
Model OR (95% CI) p pc
n % n %
rs3737965 -336 C/T
rs1801131 Glu429Ala
Genotype C/C C/T T/T Allele C T Genotype A/A A/C C/C
Allele A C
112 13 0
237 13
87 35 3
209 41
89.6 10.4 0.0
94.8 5.2
69.6 28.0 2.4
83.6 16.4
95 27 0
217 27
81 39 2
201 43
77.9 22.1 0.0
88.9 11.1
66.4 32.0 1.6
82.4 17.6
Codominant1 Codominant2 Dominant Recessive Overdominant Log-additive
2.33 (1.07∼5.09)
1 2.27 (1.14∼4.51)
1.07 (0.59∼1.97) 0.61 (0.09∼4.17) 1.03 (0.57∼1.87) 0.60 (0.09∼4.03) 1.09 (0.60∼2.00) 0.99 (0.58∼1.68)
1 1.09 (0.68∼1.74)
0.031
0.019 0.82 0.61 0.91 0.59 0.78 0.96
0.72
0.062
0.038 1.00 1.00 1.00 1.00 1.00 1.00
1.00 Genotype and allele distributions are shown as number (%). p values were from logistic regression analyses with the codominant1, codominant2, dominant, recessive, overdominant, and log-additive models controlling age and gender as covariates. pc values were calculated using Bonferroni correction.
Bold numbers mean significant associations. SNP: single nucleotide polymorphism, MTHFR: methylenetetrahydrofolate reductase, OR: odds ratio, CI:
confidence interval, n: number of subjects.
Fig. 1. Linkage disequilibrium (LD) in the methylenetetrahydrofolate reductase (MTHFR) gene. LD coefficient (│D'│) and LD block between rs3737965 and rs1801131. The LD block was constructed (│D'│=1.0 and r2=0.28).
Table 3. Frequencies of haplotypes in and overweight/obese and control subjects.
Haplo- type
Frequ- ency
Overweight/obese Control Chi Square p
+ - + -
CA CC TC
0.82 0.10 0.07
208.9 28.1 12.9
41.1 221.9 237.1
197.8 19.2 23.8
46.2 224.8 220.2
0.53 1.61 3.78
0.47 0.20 0.05 Haplotypes consist of rs3737965 and rs1801131.
DISCUSSION
Genetic variations of the MTHFR gene influence susceptibility
to several diseases, including cancers. Cui et al.(2011) reported the association between a missense MTHFR SNP (rs1801133, Ala222Val) and the risk of lung cancer, and the T allele of rs1801133 was a weak protective factor for the development of lung squamous cell carcinoma in a Korean population. Roffman et al.(2011) found that rs1801133 may affect behavior in schizophrenia patients, presumably via dopamine signaling.
Subbiah(2007) suggested that SNPs of MTHFR, apolipoprotein E
(APOE), apolipoprotein A-I (APOAI), leptin (LEP), and LEPR were
good candidates for understanding nutrigenomics. Seremak-Mrozi-
kiewicz et al.(2010) proposed the possible role of MTHFR
1793G/A polymorphism in the pathogenesis of recurrent misca-
rriages.
Recently, our group evaluated three SNPs (rs1537514, rs1559040, and rs1801133) in the exon region and seven SNPs (rs3818762, rs12121543, rs1994798, rs6541003, rs4846052, rs17037396, and rs17367504) in the intron region of the MTHFR gene in Korean obesity patients (Kim SK et al., 2010).
They found that two SNPs (rs1537514 and rs17367504) were associated with overweight/obese in Korean population. Genotype and allele distributions of rs1537514 and 17367504 showed significant differences between overweight/obese and control subjects. The CGACGCT haplotype was also associated with overweight/obese. In present study, we investigated whether SNPs (rs3737965, -336 C/T and rs1801131, Glu429Ala) were related to obesity. MTHFR SNPs have been implicated in the develo- pment of obesity (Frelut et al., 2006; Lambrinoudaki et al., 2008).
We also observed a significant association between a promoter SNP rs3737965 and overweight/obese.
MTHFR protein (P42898, UnitProt ID) consists of 656 amino acids and has a molecular mass of 74,597 Da (http://www.uniprot.
org). The rs3737965 SNP is located in the promoter region of the MTHFR gene. To determine whether alleles of rs3737965 affect transcription factors, we used an online program (AliBaba 2.1;http://www.gene-regulation.com/pub/programs/alibaba2). At the rs3737965 SNP site, T-containing sequences can act with C/EBP gamma transcription factor, but C/EBP gamma transcription factor disappears in C-containing sequences. Therefore, rs3737965 SNP may affect transcriptional activity.
In conclusion, we investigated whether SNPs of the MTHFR gene were related to overweight/obese in Korean population. The allele distributions of rs3737965 showed significant differences between overweight/obese and control subjects. This result suggests that a promoter SNP (rs3737965. -336 C/T) of the MTHFR gene may be associated with obesity in Korean population.
REFERENCES
Barth RJ (2011) Insulin resistance, obesity and the metabolic syndrome. S. D. Med. Spec No:22-27.
Calton MA, Ersoy BA, Zhang S et al. (2009) Association of functionally significant Melanocortin-4 but not Melanocortin-3 receptor mutations with severe adult obesity in a large North American case-control study. Hum. Mol. Genet. 18:1140-1147.
Chambers JC, Elliott P, Zabaneh D et al. (2008) Common genetic variation near MC4R is associated with waist circumference and insulin resistance. Nat. Genet. 40:716-718.
Cui LH, Shin MH, Kim HN et al. (2011) Methylenetetrahydrofolate reductase C677T polymorphism in patients with lung cancer in a Korean population. BMC Med. Genet. 22:28.
Grant SF, Li M, Bradfield JP et al. (2008) Association analysis of the FTO gene with obesity in children of Caucasian and African ancestry reveals a common tagging SNP. PLoS One. 3:1746.
Frelut ML, Emery-Fillon N, Guilland JC et al. (2006) Alanine amino transferase concentrations are linked to folate intakes and methylenetetrahydrofolate reductase polymorphism in obese adolescent girls. J. Pediatr. Gastroenterol. Nutr. 43:234-239.
Guenther BD, Sheppard CA, Tran P et al. (1999) The structure and properties of methylenetetrahydrofolate reductase from Esche- richia coli suggest how folate ameliorates human hyperhomocy- steinemia. Nat. Struct. Biol. 6:359-365.
Hotta K, Nakata Y, Matsuo T et al. (2008) Variations in the FTO gene are associated with severe obesity in the Japanese. J. Hum.
Genet. 53:546-553.
Jermendy A, Körner A, Kovács M et al. (2011) PPAR-gamma2 pro12Ala polymorphism is associated with post-challenge abnormalities of glucose homeostasis in children and adolescents with obesity. J. Pediatr. Endocrinol. Metab. 24:55-59.
Kim SK, Yang SA (2010) Association of methylenetetrahydrofolate reductase (MTHFR) gene polymorphisms with obesity in Korean population. Korean J. Stress Res. 18:237-243.
Lambrinoudaki I, Kaparos G, Papadimitriou D et al. (2008) Methyle- netetrahydrofolate reductase C677T polymorphism is associated with central adiposity and increased androgenicity in healthy postmenopausal women. Eur. J. Endocrinol. 159:233-241.
Patabiraman P, Pham S (2011) Obesity and cardiovascular disease. S.
D. Med. Spec No:36-39.
Pereira TV, Mingroni-Netto RC, Yamada Y (2011) ADRB2 and LEPR gene polymorphisms: synergistic effects on the risk of obesity in Japanese. Obesity 17:1523-1527.
Ravussin E, Bogardus C (2000) Energy balance and weight regulation: genetics versus environment. Br. J. Nutr. 83:17-20.
Roffman JL, Brohawn DG, Friedman JS et al. (2011) MTHFR 677 C>T effects on anterior cingulate structure and function during response monitoring in schizophrenia: a preliminary study. Brain Imaging Behav. 5:65-75.
Sánchez-Lara K, Morales-Graf L, Green D et al. (2010) Cancer and obesity. Gac. Med. Mex. 146:326-331.
Seremak-Mrozikiewicz A, Drews K, Kurzawinska G et al. (2010) The significance of 1793G>A polymorphism in MTHFR gene in women with first trimester recurrent miscarriages. Neuro.
Endocrinol. Lett. 31:717-723.
= 국문초록 =
Methylenetetrahydrofolate reductase (MTHFR)은 folate 대사에 관여하는 효소이며, 여러 연구에서 MTHFR 유전자와 비만 과의 관련성이 보고되었다. 본 연구에서는 MTHFR 유전자의 단일염기다형성과 비만과의 관련성을 보기 위하여, 122 명의 과체중/비만 환자와 125명의 정상체중인 사람을 분석하였다. MTHFR 유전자의 단일염기다형성 중 하나의 프로 모터 단일염기다형성(rs3737965, -336 C/T)와 하나의 missense 단일염기다형성(rs1801131, Glu429Ala)을 선정하였다. 염 기서열 분석은 direct sequencing을 이용하였다. 유전자 분석은 SNPStats, SPSS 18.0, Haploview 4.2 소프트웨어를 이용하 였고, Bonferroni correction을 하였다. 분석결과, Bonferroni correction 후에도 rs3737965의 allele가 과체중/비만과 유의성이 있었다(OR=2.27, 95% CI=1.14∼4.51, p
c=0.038). rs3737965의 T allele가 과체중/비만 환자 그룹에서는 11.1%이고 정상 그룹에서는 5.2%으로, 정상그룹에 비해 과체중/비만 환자 그룹에서 약 두 배 높게 나타났다. 이러한 결과는 한국인에 서 MTHFR 유전자가 비만과 관련이 있음을 시사한다.
중심단어: Methylenetetrahydrofolate reductase, 비만, 과체중, 단일염기다형성
Subbiah MT (2007) Nutrigenetics and nutraceuticals: the next waveriding on personalized medicine. Transl. Res. 149:55-61.
Sun Y, Sun J, Wang X et al. (2010) Variants in the fat mass and
obesity associated (FTO) gene are associated with obesity and C-reactive protein levels in Chinese Han populations. Clin. Invest.
Med. 33:405-412.