696
Copyright © 2015 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
반수체
(haploid)
는세포당하나의유전체또는염색체조가존재하는것으로
,
자성발생성이배체(gynogenesis diploid)
또는 웅성발생성이배체(androgenesis diploid)
유도기술에의해서 생산되어왔다.
이들은자외선에의해서유전물질이불활성화 된정자또는난자를이용하여유도됨으로써,
오로지난자또는 정자의유전물질만으로발생하게된다(Tanaka et al., 2004; Yi et al., 2009).
경골어류의 반수체에 대한 연구는 금붕어
(goldfish, Caras- sius auratus),
송사리(medaka, Oryzias latipes)
및 제브라피 쉬(zebrafish, Danio rerio)
에서보고된바있다(Walker, 1999;
Araki, 2001; Luo and Li, 2003).
이들의연구결과경골어류 의반수체는일반적으로해로운열성치사유전자존재또는유 전자산물의부족때문에부종(edema),
왜소증(dwarfism),
소 두증(microcephaly)
및소안구(microphthalmia)
등과같은이 형적인특징이관찰되었고,
부화전후또는난황흡수후모두 사망하는것으로보고되어왔으며(Felip et al., 1999; Luo and
Li, 2003),
이러한반수체의특이적인 이상발생 현상은반수체증후군
(haploid syndrome)
으로명명되어왔다(Tvedt et al.,
2006).
그러나반수체는한개의염색체조만을가짐으로써우성유전자형또는열성유전자형에대한표현형을모두관찰할 수있기때문에효율적인유전자분석및검색의방법론으로이 용되고있다
(Yi et al., 2009).
이에제브라피쉬는반수체배아의반수체 넙치(Paralichthys olivaceus)의 발생학적 특성 분석
정효선·김연경·김현철
2·노재구
2·이정호
2·김동수
1*
부경대학교 해양수산 LMO 위해성평가센터, 1부경대학교 해양바이오신소재학과, 2국립수산과학원 육종연구센터
Embryonic Development of Haploid Olive Flounder Paralichthys olivaceus
Hyo Sun Jung, Youn Kyoung Kim, Hyun Chul Kim
2
, Jae-Koo Noh2
, Jeong-Ho Lee2
and Dong Soo Kim1
*Center for Risk Assessment of Oceans and Fisheries Living Modified Organisms, Pukyong National University, Busan 48513, Korea
1
Department of Marine Bio-Materials & Aquaculture, Pukyong National University, Busan 48513, Korea
2
Genetics and Breeding Research Center, National Fisheries Research and Development Institute, Geoje 53334, Korea We investigated the characteristics of embryonic and abnormal organ development in haploid olive flounder, Parali- chthys olivaceus, by comparing egg development and histological sections in haploid and diploid individuals. After the mid-blastula transition, abnormal development was observed in haploid fish, including delayed epiboly and mal- formation of the germ ring and embryonic body. In haploid flounder, Kupffer’s vesicles are irregularly shaped and of variable size compared to diploids. The embryonic body of haploids was shorter and broader than that of diploids and the tail length and size were variable. Most haploid embryos failed to hatch and the few larvae that did, did not survive for more than 6 h. The histological analysis of haploid larvae revealed deformed development in diverse or- gans, including the eye, otic vesicles, notochord, and neural tube. These results may be related to an abnormality in the axial system of haploid larvae. This study confirmed that the abnormalities of haploid olive flounder were similar to the reported characteristics of haploid syndrome. The abnormalities are caused by delayed epiboly and involution and deformity of Kupffer’s vesicle during egg development.
Key words: Haploid , Olive flounder, Paralichthys olivaceus
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2015.0696 Korean J Fish Aquat Sci 48(5) 696-703, October 2015
Received 6 June 2015; Revised 30 August 2015; Accepted 16 September 2015
*Corresponding author: Tel: +82. 51. 629. 5914 Fax: +82. 51. 629. 5908
E-mail address: [email protected]
돌연변이유발유전자들에대한검색을통해심장
(heart),
척색(notochord)
및근육(muscle)
의발달에필요한유전자를확인 한바있으며(Wiellette et al., 2004),
반수체배아에서돌연변 이눈의발생을통해눈이정상적으로발생되는데필요한유전 자발현조건을금붕어에서보고한바있다(Luo and Li, 2003).
넙치
(olive flounder, Paralichthys olivaceus)
는한국,
일본및 중국을포함하는아시아에서양식되는중요한어류로써(Zheng
and Sun, 2011)
이들의생산성및경제성을높이기위해서성장
,
면역체계및형태등에대한유전자연구가이루어져왔으 나(Yang et al., 2013),
이는이배체를이용하여이루어졌기때 문에반수체를이용한더정확하고복합적인유전자검색연구 가필요시된다.
이를위해본연구에서는넙치의반수체를유 도하여그의발생과정및기형적인발생특징에대하여이배체 와비교분석하였다.
재료 및 방법
실험어
본연구에사용된넙치는국립수산과학원육종연구센터에서 선발육종기법을통하여생산된킹넙치
(KingNupchi)
가사용되 었으며,
복부압박법으로암컷7
미와수컷3
미로부터확보된난 과정자를실험에사용하였다.
반수체 유도
반수체
(haploid)
는자외선조사에의해유전적으로불활성화된정자와정상난자를수정함으로써유도되었다
. Kim et al.
(1993)
과Yamamoto (1999)
에서보고된자외선조사량을바탕 으로,
넙치정자의완벽한유전적불활성화를위해5,400 ergs/
mm
2의자외선을조사한후습식법으로인공수정시킨뒤,
수정란은
18±1℃
의부화수조에서발생을유도하였다.
DNA 함량분석
유도된반수체의
DNA
함량분석을위하여flow cytometry
를 수행하였다.
반수체증후군(haploid syndrome)
의형태를가지 고있는자어10
미를모아균질기로세포를파쇄한후, Jung et
al. (2014)
의방법론에따라핵막제거및염색과정을수행하였다
.
염색된시료들은PA II flow cytometry (Partec Co., Mün- ster, Germany)
를이용분석하였으며,
성체넙치꼬리지느러미 및이배체자어를대조군으로사용하였다.
난발생 관찰
넙치반수체및이배체배아는
18±1℃
의수온에서부화시키며
,
해부현미경(AZ100; Nikon Co., Japan)
과이에부착된디지 털카메라(DS-Ril; Nikon Co., Tokyo, Japan)
를이용하여관찰 과촬영을하였다.
각발생단계는Kimmel et al. (1995)
의배발생단계를기준으로정하였고
,
수정된개체의90%
이상이동일한단계에도달하였을시기를기준으로각발생단계별소요시
간과특징을기록하였다
.
또한발생단계별정확한시간을측정하기위해
16±1℃
와20±1℃
의온도조건에서발생시간을비교하여분석하였다
. 조직학적 분석
반수체의특이적인기관발생을관찰하기위해부화자어의 조직을분석하였다
.
형태학적으로반수체증후군의형태를가 지고있는자어를4% formaldehyde
에고정한뒤,
통상의조직 학적분석방법에따라탈수및파라핀을이용한포매과정을수행하였다
.
준비된표본는6 µm
두께로연속절편하여hema-
toxylin
과eosin
으로염색을한후,
광학현미경(Eclipse E400;
Nikon Co., Japan)
하에서검경하였고,
이배체와비교하여특 이적인 조직상은 광학현미경에 부착된디지털 카메라(Moti- campPro 205A; Motic Co., China)
로촬영하였다.
결과 및 고찰
하나의염색체조만을가지는반수체는효율적인 유전자분 석및검색의방법론으로이용되고있다
.
그러나이들분석방 법에반수체를이용하기위해서는먼저유도된반수체의발생 학적특징에대한연구가선행되어야한다.
이에본연구는넙 치반수체를유도하였고,
생물의고유한DNA
함량으로배수성을판단할수있는
flow cytometry
분석법을이용하여유도된반수체넙치의
DNA 햠
량을측정하였다.
이에이미보고되어있는넙치의
DNA
함량(0.71 pg/cell; Animal genome size database, 2014)
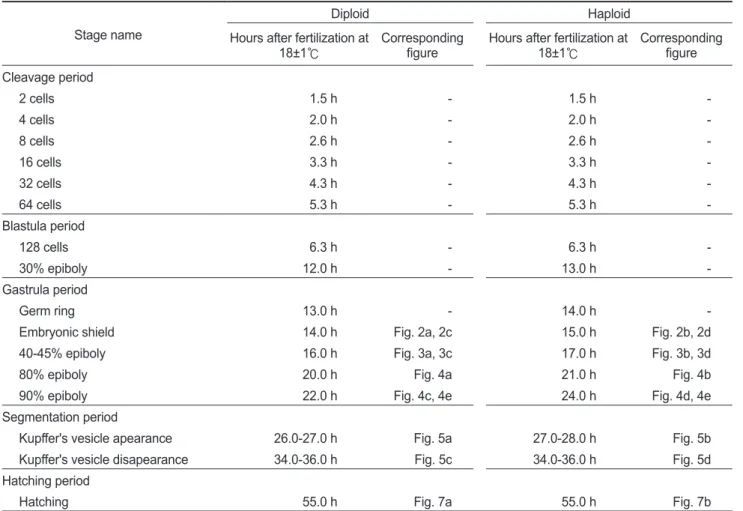
을대조군으로하여측정한결과,
이배체부화 자어의DNA
함량은0.71±0.02 pg/cell
으로확인되었고,
반수 체는0.34±0.01 pg/cell
으로확인되었다(Fig. 1).
이는이배체넙치의
DNA
함량의50%
에해당하는것으로써성공적으로반수체가유도된것으로확인할수있었다
.
유도된반수체넙치의배발생시기및특징을관찰한결과
, 2
세포기에서128
세포기에해당하는난할및초기포배기동안 은반수체와이배체의발생시기및형태가동일한것으로관 찰되었다(Table 1). Walker (1999)
는제브라피쉬반수체배아 에서도초기포배기까지이배체와발생시간이동일함을보고 한바있다.
128
세포기이후부터반수체의발생시간이대조군에비해점차느려져
30%
피포형성이이배체는수정후12
시간이소요됨에비하여반수체의경우
13
시간이소요되었다(Table 1).
이후 이배체에서는수정13
시간후최초로배환(germ ring)
이관찰 되었고,
반수체배아에서는수정14
시간후에관찰되었다.
이때 반수체배환은이배체에비하여진하고두껍게형성되는것으 로확인되었다(Figs. 2-4).
더욱이피포형성비율이높아질수록 이배체의배환은옅어지는반면반수체는배환이점차진하고 두껍게형성되는것이관찰되었으며피포형성속도또한대조군에비하여점차더지연되어
90%
피포형성은반수체가2
시간정도늦게관찰되었다
(Table 1, Figs. 2-4).
배체(embryonic
body)
의형성은이배체배아의경우80%
피포형성단계에서형태가나타나기시작하여
90%
피포형성단계에서뚜렷한배체가형성되었으나
,
반수체배아는이들보다늦어져90%
피포 형성단계에도달해야비로소불명확한형태의짧은배체가넓 게형성됨이관찰되었다(Fig. 4).
낭배기에배환은배반엽
(blastoderm)
이난황을감싸면서일어나는 회절
(involution),
수렴(convergence)
및 확장(exten- sion)
의형태발생세포운동(morphogenetic cell movements)
중가장먼저관찰되는회절
(involution)
이일어남으로써형성된다
(Kimmel et al., 1995).
본연구결과반수체배아의배환 이대조군에비하여매우진하고두껍게형성되며피포형성시 간또한늦어지는것이관찰되었다. Araki et al. (2001)
는송사 리반수체배아가이배체배아보다피포와회절이늦게관찰되 는것은반수체가한쌍의염색체조만을가짐으로써비특이적인 세포사멸이일어나고,
정상적인피포와회절발생에영향을주 게됨으로써이배체에비해늦은속도로발생한다고보고한바 있다.
이에넙치반수체배아도비특이적으로사멸된세포들이 존재하게되면서이배체에비해두껍고진한배환이형성됨으로피포및회절발생이지연되는것으로판단된다
.
또한배체 의발생은배반엽세포의수렴과확장과정에의해형성되고이 과정들이진행될수록배체는점차좁아지고신장되면서더명 확해지게된다(Kimmel et al., 1995).
그러나본연구의반수체 배아는대조군에비하여배체의형태가넓고,
짧으며,
분명하지 않은상태로발생하는것으로관찰되었다.
송사리반수체배아 에서도초기배체형태가짧고,
넓게형성되었고이것의원인을 회절지연현상으로보고한바있어(Araki et al., 2001),
넙치반 수체배아에서이형적인초기배체형태는비정상적인형태발 생세포운동(
회절,
수렴및신장)
과정에의한것으로판단된다. Kupffer’s vesicle
은반수체배아에서이배체보다한시간늦은수정후
27-28
시간에관찰되었고,
사라지는시기는거의동일하여이배체배아에비해적은시간동안존재하는것으로확 인되었다
(Table 1).
반수체배아에서Kupffer's vesicles
은이배체배아와동일한위치인꼬리싹
(tail bud)
에서형성이되지만,
그형태와크기가개체에따라불규칙적으로발달하는것으로 관찰되었다
(Fig. 5).
이에꼬리중배엽을형성하는데영향을주 는Kupffer's vesicles
이이형적으로형성됨으로써배체의꼬리Table 1. Stage of embryonic development in diploid and haploid Paralichthys olivaceus Stage name
Diploid Haploid
Hours after fertilization at
18±1℃ Corresponding
figure Hours after fertilization at
18±1℃ Corresponding
figure Cleavage period
2 cells 1.5 h - 1.5 h -
4 cells 2.0 h - 2.0 h -
8 cells 2.6 h - 2.6 h -
16 cells 3.3 h - 3.3 h -
32 cells 4.3 h - 4.3 h -
64 cells 5.3 h - 5.3 h -
Blastula period
128 cells 6.3 h - 6.3 h -
30% epiboly 12.0 h - 13.0 h -
Gastrula period
Germ ring 13.0 h - 14.0 h -
Embryonic shield 14.0 h Fig. 2a, 2c 15.0 h Fig. 2b, 2d
40-45% epiboly 16.0 h Fig. 3a, 3c 17.0 h Fig. 3b, 3d
80% epiboly 20.0 h Fig. 4a 21.0 h Fig. 4b
90% epiboly 22.0 h Fig. 4c, 4e 24.0 h Fig. 4d, 4e
Segmentation period
Kupffer's vesicle apearance 26.0-27.0 h Fig. 5a 27.0-28.0 h Fig. 5b
Kupffer's vesicle disapearance 34.0-36.0 h Fig. 5c 34.0-36.0 h Fig. 5d
Hatching period
Hatching 55.0 h Fig. 7a 55.0 h Fig. 7b
는대조군과비교하여이형적으로발생하는것으로관찰되었 다
(Fig. 7). Kupffer’s vesicle
은경골어류의배아에서만일시적 으로관찰되는구체기관으로,
섬모세포를이용하여좌우비대 칭을조정하는역할을한다(Essner et al., 2005).
이에Okabe et al. (2008)
은제브라피쉬발생과정중Kupffer’s vesicle
내에 서일어나는유체역학적인현상을주사전자현미경, video mi-
croscopy
및조직학적으로밝혀경골어류의발생과정중그의중요성을입증한바있다
.
본연구에서넙치의반수체배아의Kupffer’s vesicle
은모양과크기가다양하였으며,
배체의형태 도대조군과뚜렷하게구별되었다.
이는좌-
우축형성에영향을 미치는Kupffer’s vesicle
이반수체배아에서이형적으로발생 함으로써,
이들의후기배발생과정에서다양한기관들의정상 적인발달에부정적인영향을준것으로판단된다.
Kupffer’s vesicle
이소실된후배체가부화하기까지두실험 군간발생시간의차이가관찰되지않아최초의부화는반수체 와이배체모두수정후55
시간으로동일하였으나,
이배체및반수체부화율이각각
80.0±9.4 %
및6.9±3.0%
로반수체는 일부개체만이부화하는것으로관찰되었다.
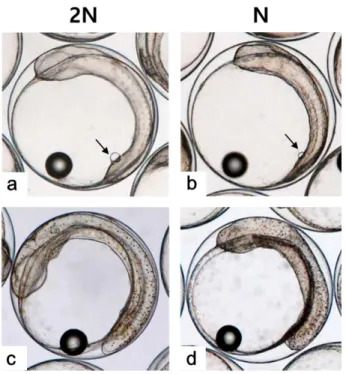
반수체부화자어 의형태는개체에따른차이가존재하나대부분꼬리가제대로 발달하지못하였고,
몸체의길이와넓이가이배체에비하여짧 고,
넓은것으로관찰되었다(Figs. 6, 7).
반수체처리군1,000
알중꼬리및몸체가기형적으로발생한개체는962
개로그 중꼬리가발달되지못한형태를띠는개체(Fig. 7b)
는903
개,
꼬리가생성은되었으나제대로발달되지못한개체
(Fig. 7c)
는
59
개로확인되었다.
이들중꼬리가전혀발달되지않은개 체들은부화를하지못하고죽는것으로관찰되었으며,
꼬리가 이형적으로발생된개체는부화하였을지라도6
시간이내에모 두죽는것으로관찰되었다.
금붕어,
송사리,
제브라피쉬,
터봇(turbot, Scophthalmus maximus)
그리고Atlantic halibut (Hip- poglossus hippoglossus)
에서도반수체배아의낮은부화율과 부화자어의짧은몸체,
꼬리의미발달및높은치사율이보고된 바있으며(Luo and Li, 2003; Araki et al., 2001; Walker, 1999;
Piferrer et al., 2004; Tvedt et al., 2006),
이러한현상을Felip et al. (1999)
는반수체의유전체부족에따른부화효소(hatching
enzyme)
와같은발생과정및부화에반드시필요한단백질합성량의부족때문으로추론한바있다
.
반수체의다양한배발생단계에서이상발생및발생의지연
Fig. 1. Flow cytometric histograms for DNA content in diploid (2N) and haploid (N) Paralichthys olivaceus.
720 640 560 480 100 320 240 160 8 0
No. of cells
2N
720 640 560 480 100 320 240 160 8
0 100 200 300 400 500 600
N
No. of cells
Fig. 2. External morphology of embryonic shield stage in diploid (a,c) and haploid (b,d) Paralichthys olivaceus. a, c: face view (a) and animal view (c) of embryonic shield stage in diploid at 14 h after fertilization. b, d: face view (b) and animal view (d) of embry- onic shield stage in haploid at 15 h after fertilization. Embryonic shield, arrow.
현상이관찰됨으로써반수체에서각기관들이발달되는형태 적특징을확인하고자부화자어의횡당면및종단면에 대한 조직분석을수행하였고그결과는
Fig. 8
및Fig. 9
와같다.
수 정체(lens)
와신경망막(neural retina)
이정상적으로발달된눈 이분명하게형성되어있는이배체자어와는달리,
반수체자어 는수정체와신경망막이불분명하고,
양쪽눈이비대칭적인위 치에서형성되는것으로확인되었다(Fig. 8a, 8b).
또한귀소포(otic vesicles)
의경우,
이배체에서는양쪽에각각한개씩위치하지만
(Fig. 8c),
반수체에서는이형적인형태와개수로비대칭적으로형성되는것이관찰되었다
(Fig. 8d).
금붕어반수체자 어의눈에서도색소망막(pigmented retina),
신경망막,
수정체,
각막
(cornea)
과같은눈을구성하는세포들이모두형성은되지만
,
각세포들이비정상적인위치에서형성됨으로써이형적 인눈의형태로발달된다고보고된바있다(Luo and Li, 2003;
Tanaka et al., 2004).
본연구의결과역시기존의보고에서기 술된결과들과유사한양상을나타내었다.
이에반수체에서도 각기관을구성하는세포들이형성은되지만,
그위치및형태 가비정상적으로발달하는것이반수체기관발달의특징인것 으로판단된다.
신경관
(neural tube)
은 일반적으로 배발생과정에서 신경판(neural plate)
이neural keel, neural rod
그리고신경관의순서 대로발달하지만(Clarke, 2009),
본종의이배체부화자어는신 경관의내부공간이생성되지않은형태인neural rod
까지발달 하는것으로확인되었다(Fig. 8e).
이는넙치가짧은배발생기 간을가지기때문에신경관까지발달하지못하고부화하는것 으로보인다.
그러나반수체에서는구형의형태조차띄지않는neural keel
의형태로관찰되어신경기관의발생지연현상이관찰되었다
(Fig. 8f).
또한반수체에서장(intestine)
은형태,
크기및내강
(lumen)
의유무가각개체별로불규칙적으로발달되는것으로관찰되었다
(Fig. 8f).
Fig. 3. External morphology of 40 - 45% epiboly stage in diploid (a,c) and haploid (b,d) Paralichthys olivaceus. a, c: face view (a) and animal view (b) of 40-45% epiboly stage in diploid at 16 h after fertilization. b, d: face view (b) and animal view (d) of 40- 45% epiboly stage in haploid at 17 h after fertilization. Germ ring, arrow.
Fig. 4. Morphology of embryonic body in 80-90% epiboly stage of diploid (a, c, e) and haploid (b, d, e) Paralichthys olivaceus. a:
Ventral view of 80% epiboly stage in diploid at 20 h after fertiliza- tion. b: ventral view of 80% epiboly stage in haploid at 21 h after fertilization. c, e: Ventral view (c) and left side view (e) of 90%
epiboly stage in diploid at 21 h after fertilization. d, f: Ventral view (d) and left side view (f) of 90% epiboly stage in haploid at 24 h after fertilization. Embryonic body, arrow.
중요한뼈대역할을하는척색
(notochord)
은배발생과정에서 중심세포가부풀게되면서액포가있는상태로발달한뒤,
길게 늘어진형태가되는과정을거치게된다(Stemple et al., 1996;
Anderson et al., 2007).
그러나반수체에서척색은액포세포가 길게늘어지지않고동그란형태를유지하고,
그형태및크기 가균일하지않게발달하는것으로관찰되었다(Fig. 8f).
송사리 반수체에서도본종의관찰결과와동일하게척색의중심에존 재하는액포세포들이길게늘어나는과정없이불규칙하게둥 근형태를유지하는것이확인된바있다(Araki et al., 2001).
부화자어의종단면조직분석 결과
,
반수체에서도체절(so-
mite)
이발달은되지만,
그형태가불규칙하고각체절간의구분이불분명한것으로관찰되었다
(Fig. 9).
또한장이몸통부(trunk)
까지 발달되어 항문을 형성하는 이배체와는 달리,
반수체는꼬리가거의형성되지못함으로써장이몸통부까지형 성되지못하고항문또한형성되지못하는것으로관찰되었다
(Fig. 9).
이상의연구결과
,
넙치반수체배아는피포가시작되는후기 포배기부터발생지연및이상발생현상나타나기시작하여짧 은꼬리및넓은몸통을가진배체로발생하였다.
또한부화단계에서일부개체만이부화하였으며부화한개체도모두치사 함으로써
,
넙치반수체의이상발생현상은다양한경골어류에 서보고된반수체증후군(haploid syndrome)
의특징들과유사 한것으로확인되었다.
이러한넙치반수체이상발생및치사현 상은기존의연구에서보고된바와같이한개의유전체만을가 지는반수체특성에의한열성치사유전자의발현또는유전자 산물의부족으로인한것으로판단된다.
또한반수체에서이형적으로발달된기관들은대부분축시스템
(axial system)
과연관된것들로눈및귀소포와같은기관은좌
-
우비대칭적으로 발생하였고신경관,
척색,
장및체절과같이몸체에서중심을 이루는기관들은대부분불완전하게발생하거나발생하지않 는것으로관찰되었다.
이는반수체배발생과정에서Kupffer’s
vesicle
이이형적으로발생함으로써정상적인좌-
우축이형성되지못하여눈및귀소포의이상발생이나타난것으로판단되 고
,
앞-
뒤축이형성되는낭배기시기의이상발생으로신경관,
척색및장과같은기관들에서이상발생이관찰되는것으로보 여진다.
앞으로반수체자어의부화전후나타나는조기치사현상으
Fig. 5. Appearance and disappearance of Kupffer's vesicle in dip- loid (a,c) and haploid (b,d) Paralichthys olivaceus. a: Appearance of Kupffer's vesicle (arrow) in diploid at 25 h after fertilization. b:
Appearance of Kupffer's vesicle (arrow) in haploid at 26 h after fertilization. c, d: Disappearance of Kupffer's vesicle in diploid (c) and haploid (d) at 34-36 h after fertilization. Kupffer's vesicle, ar- row.
Fig. 6. Extension of tail bud in diploid (a, c, e) and haploid (b, d, f) Paralichthys olivaceus. a, b: The diploid (a) and haploid (b) em- bryo at 34 h after fertilization. c, d: The diploid (c) and haploid (d) embryo at 42 h after fertilization. e,f: The diploid (e) and haploid (f) embryo at at 54 h after fertilization.
Fig. 8. Histological cross sections of head region in diploid and haploid Paralichthys olivaceus at hatching stage. a, b: Eyes (ey) of a diploid (a) and haploid (b) larva. Neural retina, arrows; lens, ar- rowheads. c, d: Otic vesicles of a diploid (c) and haploid (d) larva.
Otic vesicles, arrowheads. e: Neural rod (nr), notochord (n) and intestine (i) of a diploid larva. f: Neural keel (nk), notochord (n) and intestine (i) of a haploid larva. All scale bars indicate 50 μm.
로인하여그연구에한계가있는넙치의우성및열성유전자 검색연구를위해반수체
-
이배체키메라(haploid-diploid chi- mera)
의유도연구가뒤따라야할것이다.
사 사
본 연구는 농림축산식품부
∙
해양수산부∙
농촌진흥청∙
산림 청의 지원과제 수출용 킹넙치의 배수체 종자개발(
과제번호213004-04-3-1SB220)
에의해수행되었습니다.
References
Anderson C, Bartlett SJ, Gansner JM, Wilson D, He L, Gitlin JD, Kelsh RN and Dowden J. 2007. Chemical genetics suggests a critical role for lysyl oxidase in zebrafish noto- chord morphogenesis. Mol Biosyst 3, 51-59. http://dx.doi.
org/10.1039/B613673G.
Animal Genome Size Database. 2014. Vertebrates-Fishes hap- loid C-value. Retrieved from http://www.genomesize.com on March 27.
Araki K, Okamoto H, Graveson AC, Nakayama I, Nagoya H.
2001. Analysis of haploid development based on expres- sion patterns of developmental genes in the medaka Oryzias
latipes. Develop Growth Differ 43, 591-599. http://dx.doi.
org/10.1046/j.1440-169X.2001.00601.x.
Clarke J. 2009. Role of polarized cell divisions in zebrafish neu- ral tube formation. Curr Opin Neurobiol 19, 134-138. http://
dx.doi.org/10.1016/j.conb.2009.04.010.
Essner JJ, Amack JD, Nyholm MK, Harris EB and Yost HJ.
2005. Kupffer's vesicle is a ciliated organ of asymmetry in the zebrafish embryo that initiates left-right development of Fig. 7. The shape and length of tail in Just-hatched larva of diploid
(a) and haploid (b) Paralichthys olivaceus. All scale bars indicate 0.25 mm
Fig. 9. Microphotography and histological section of diploid and haploid Paralichthys olivaceus at hatching stage. a: Microphotog- raphy of just-hatched larva in diploid. b: Microphotography of haploid embryo at hatching stage. c, d: Longitudinal section of just-hatched larva in diploid (c) and haploid (d). Anus (urogenital opening), arrow.
the brain, heart and gut. Dev 132, 1247-1260. http://dx.doi.
org/10.1242/dev.01663.
Felip A, Piferrer F, Carrillo M and Zanuy S.1999. The relation- ship between the effects of UV light and thermal shock on gametes and the viability of early developmental stages in a marine teleost fish, the sea bass (Dicentrarchus labrax L.). Heredity 83, 387-397. http://dx.doi.org/10.1038/sj.hdy.
6886030.
Jung HS, Kim YK, Kim HC, Noh JK, Lee JH and Kim DS.
2014. Cytogenetic Analysis of Starry Flounder Platichthys stellatus from Korea. Korean J Fish Aquat Sci 47, 431-434.
http://dx.doi.org/10.5657/KFAS.2014.0431.
Kim DS, Kim JH, Jo JY, Moon YB and Cho KC. 1993. Induc- tion of gynogenetic diploid in Paralichthys olivaceus. Ko- rean J Genetics 15, 179-186.
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B and Schil- ling TF. 1995. Stages of embryonic development of the ze- brafish. Dev Dyn 203, 253-310. http://dx.doi.org/10.1002/
aja.1002030302.
Luo C and Li B. 2003. Diploid-dependent regulation of gene expression: a genetic cause of abnormal development in fish haploid embryos. Heredity 90, 405-409. http://dx.doi.
org/10.1038/sj.hdy.6800263.
Okabe N, Xu B and Burdine RD. 2008. Fluid dynamics in ze- brafish Kupffer's vesicle. Dev Dyn 237, 3602-3612. http://
dx.doi.org/10.1002/dvdy.21730.
Piferrer F, Cal RM, Gomez C, Alvarez-Blazquez B, Castro J and Martınez P. 2004. Induction of gynogenesis in the turbot (Scophthalmus maximus): Effects of UV irradiation on sperm motility, the Hertwig effect and viability during the first 6 months of age. Aquaculture 238, 403-419. http://
dx.doi.org/10.1016/j.aquaculture.2004.05.009.
Stemple DL, Solnica-Krezel L, Zwartkruis F, Neuhauss SC, Schier AF, Malicki J, Stainier DY, Abdelilah S, Rangini Z, Mountcastle-Shah E and Driever W. 1996. Mutations affect- ing development of the notochord in zebrafish. Dev 123, 117-128. http://dx.doi.org/10.5167/uzh-238.
Tanaka M, Yamaha E and Arai K. 2004. Survival capacity of haploid-diploid goldfish chimeras. J Exp Zool A Comp Exp Biol 301, 491-501. http://dx.doi.org/10.1002/jez.a.48.
Tvedt HB, Benfey TJ, Martin-Robichaud DJ, McGowan C and Reith M. 2006. Gynogenesis and sex determination in Atlantic Halibut (Hippoglossus hippoglossus). Aqua- culture 252, 573-583. http://dx.doi.org/10.1016/j.aquacul- ture.2005.06.042.
Walker C. 1999. Haploid screens and gamma-ray mutagenesis.
In: Methods in Cell Biology. Detrich HW and Westerfield M, eds. Academic Press, San Diego, U.S.A., 43-70.
Wiellette E, Grinblat Y, Austen M, Hirsinger E, Amsterdam A, Walker C, Westerfield M and Sive H. 2004. Combined hap- loid and insertional mutation screen in the zebrafish. Genesis 40, 231-240. http://dx.doi.org/10.1002/gene.20090.
Yamamoto E. 1999. Studies on sex-manipulation and produc- tion of cloned populations in hirame, Paralichthys olivaceus (Temminck et Schlegel). Aquaculture 173, 235-246. http://
dx.doi.org/10.1016/S0044-8486(98)00448-7.
Yang H, Lee YM, Lee JH, Noh JK, Kim HC, Park CJ, Park JW, Hwang IJ and Kim SY. 2013. Expression of perforin gene for early development of nephrons in olive flounder (Para-
lichthys olivaceus). Dev Reprod 17, 321-327. http://dx.doi.
org/10.12717/DR.2013.17.4.321.
Yi M, Hong N and Hong Y. 2009. Generation of medaka fish haploid embryonic stem cells. Science 326, 430-433. http://
dx.doi.org/10.1126/science.1175151.
Zheng WJ and Sun L. 2011. Evaluation of housekeeping genes as references for quantitative real time RT-PCR analysis of gene expression in Japanese flounder (Paralichthys oliva-
ceus). Fish Shellfish Immunol 30, 638-645. http://dx.doi.
org/10.1016/j.fsi.2010.12.014.