149
책임저자: 정상영, 광주시 동구 학1동 8번지 501-757, 전남대학교병원 외과 Tel: 062-220-6472, 6456, Fax: 062-227-7635 E-mail: [email protected]
접수일:2008년 3월 26일, 게재승인일:2008년 6월 2일
Mycophenolic Acid가 사람 Jurkat 세포의 자멸사에서 활성 산소 생성에 미치는 영향
전남대학교 의과대학 외과학교실
이동규ㆍ이호균ㆍ최수진나ㆍ정상영
Mycophenolic Acid Induced Apoptosis in Human Jurkat Cells via the Generation of Reactive Oxygen Species
Dong Kyu Lee, M.D., Ho Kyun Lee, M.D., Soo Jin Na Choi, M.D., Sang Young Chung, M.D.
Department of Surgery, Chonnam National University Medical School, Gwangju, Korea
Purpose: Mycophenolic acid (MPA) is the active agent of mycophenolate mofetil (MMF), which is an immunosuppressive drug. MPA is a selective inhibitor of inosine monophosphate dehydrogenase. The aim of this study was for demonstrate that mycophenolic acid induces apoptosis in human Jurkat cells via the generation of reactive oxygen species (ROS).
Methods: The cells were cultured in the presence or absence of MPA. Flow cytometric analysis was performed after propidium iodide staining. Western blotting for caspase 3, Bcl-2 and Bax proteins was also performed.
Results: MPA decreased the viability of Jurkat cells in a dose- and time-dependent manner. The MPA induced apoptotic cell death displayed nuclear fragmentation and sub G0/G1 phase arrest in the Jurkat cells. The expression of caspase-3 proteases in the MPA treated-Jurkat cells increased in a time-dependent manner. Treatment with MPA resulted in increased ROS generation in the Jurkat cells. There was a decreased expression of Bcl-2 and an increased expression of Bax protein in the MPA treated Jurkat cells.
Conclusion: This result suggests that MPA-induced cytotoxicity is associated with a direct increase of both ROS generation and the expression of Bax protein. (J Korean Surg Soc 2008;75:149-155)
Key Words: Mycophenolic acid, Reactive oxygen species, Immunosuppressive agent 중심 단어: 마이코페놀릭산, 활성 산소종, 면역 억제제
서 론
임상적으로 사이클로스포린(Cyclosporin A, Csy)이 널리 사용된 이후 여러 면역억제제들이 개발되었는데, 그 중 mycophenolate mofetil (MMF)은 동물 실험과 임상연구를 통 해 신장이식 후 급성 거부반응의 발생 억제에 매우 효과적
인 약물로 평가 받고 있다.(1) Mycophenolic acid (MPA)의 morpholinoethylester 형태인 MMF는 MPA의 생체 이용률을 증가시키기 위해 합성되었으며, 간에서 ester가 가수 분해되 어 MPA로 전환된다.(2) MPA는 Penicillium 종에서 추출되 어 항진균제 및 항생제로 개발된 후 림프구의 증식에 대한 선택적 억제작용이 보고되었다.(3) 현재 잘 알려진 MPA의 생체 내 면역억제 기전은 퓨린 신 합성에 중요한 inosine monophosphate dehydrogenase (IMPDH)와 guanylate synthe- tase의 활성을 저해하여 세포 내 guanosine triphosphate (GTP)와 deoxy-GTP (dGTP)를 고갈시켜 DNA 합성을 차단 하고 세포 증식을 억제하는 것이다. 특히, 림프구는 핵산 합
성을 퓨린 신 합성에 의존함으로 MPA에 선택적으로 증식 이 억제된다.(4) 그 외에 MPA는 활성화된 B 림프구의 항체 생성을 억제하며,(1) 동맥 평활근 세포의 증식을 저해한 다.(5) 또한, MPA는 활성화된 사람 T 및 B 세포에서 세포 자멸사를 유도한다.(6)
세포 자멸사는 세포내부의 계획된 신호 체계에 따라 여 러 유전자들이 발현되는 능동적인 죽음이며, 그 과정에서 생성된 세포 자멸사 소포체들은 주변의 식세포 작용에 의 해 제거되어 염증을 유발하지 않는다. 세포 자멸사의 형태 적, 생리적 특징으로는 세포의 수축, 세포막 융기, 염색사의 응축, DNA의 분절 및 세포 자멸사 소포체의 형성 등이 보 고되었다.(7) 면역계에서 발생하는 세포 자멸사는 그들의 기능 수행이나 분화과정 기작으로 이해되어진다. 세포의 죽음이 자가 반응성 T 세포와 무반응성 B 세포 제거와 같은 정상적인 면역체계 형성에 중요하기 때문이다.(8)
활성 산소종은 활성화된 호중구, xanthine oxidase, 미토콘 드리아 호흡 및 아라키돈산 대사 과정을 통해서 생성된 다.(9) Superoxide dismutase, catalase, glutathione peroxidase 같은 항산화 효소 등이 세포 내에서 생성된 활성 산소종을 제거하기 위한 생리적 방어 기전으로 작용하지만, 과도한 활성 산소종은 이들 방어 체계를 초과하여 세포 손상을 초 래한다.(10,11)
MPA에 의한 사람 T 림프구의 세포 죽음에 대한 다양한 보고가 있으나 이에 대한 정확한 기전은 부족한 실정이다.
따라서 본 연구에서는 MPA에 의한 Jurkat 세포의 죽음에서 caspase family 단백질의 활성화, 활성 산소 생성, Bcl-2 family 단백질의 발현 변화 등을 조사하여 세포 자멸사 조 절 기전을 규명하고자 하였다.
방 법
1) 재료
(1) 세포주: 사람 T 림프구 세포주인 Jurkat 세포는 한국 세포주 은행(KNCC, 서울대학교)으로부터 구입, 계대 배양 하면서 실험을 실시하였다.
(2) 시약 및 기기: 실험에 필요한 RPMI 1640, 항생제 및 우태아 혈청(fetal bovine serum, FBS)은 GIBCO BRL사 (Grand Island, NY, USA) 제품을, 배양용기(24 well plate와 10 cm dish)는 Falcon사(Becton Dickinson, San Jose, CA, USA)에 서 구입하여 사용하였다. 3-[4,5-dimethylthiazol-2-yl]-2,5-diphe- nyltetra zolium bromide (MTT), DAPI, bicinchoninic acid
(BCA), dimethyl sulfoxide (DMSO), CoPPIX는 Sigma사(St.
Louis, Missouri, USA)에서 구입하였다.
2) 방법
(1) Jurkat 세포주 배양 및 시약 처리: Jurkat 세포는 CO2 세포 배양기(37oC, 5% CO2)에서 10% 우태아 혈청이 포함 된 RPMI 1640 배지로 배양하였으며, 24시간 간격으로 배양 액을 교체하여 log phase에 있는 세포에 MPA를 처리한 후 세포 자멸사 현상과 이에 연관된 생화학 실험을 수행하였 다. MPA는 10 mM 농도로 PBS에 녹여 70oC에 보관하였 고, RPMI 1640 배지에 희석하여 사용하였다.
(2) 세포 활성도 측정: 세포의 활성도 측정은 MTT 분석법 으로 측정하였다. 세포 배양판(24-well plate)에 세포(1×105 세포수/ml)를 1 ml씩 분주하여 3시간 이상 CO2 세포배양기 안에서 안정시킨 후 시료를 각각의 조건에 따라 처리하였 다. MTT 용액(5 mg/ml, phosphate buffered saline: PBS, pH7.4)은 각각의 배양 세포에 배양액의 1/10을 첨가하여 4 시간 반응시켰다. 살아있는 세포에 의해 생성된 보라색 formazan은 10% SDS가 포함된 0.01N HCl 용액을 1 ml/well 에 첨가하여 용해시킨 후, 분광 광도계(THERMO max, USA)를 이용하여 540 nm 파장에서 흡광도를 측정하였다.
세포 활성도는 정상 대조군과 비교하여 백분율(%)로 표시 하였다.
(3) 유식 세포분석(Flow cytometry)을 이용한 sub G0/G1 분 석: 세포 자멸사에 미치는 영향을 알아보기 위해 propidium iodide (PI)로 DNA를 염색한 후에 flow cytometry (FACS- Calibur, BD Biosciences)를 이용하여 형광의 세기를 측정하 였다. 세포에 MPA를 처리하여 시간별로 포집하여 PBS로 두 번 세척하였다. 세척한 세포(10×105)의 DNA는 PI 용액 (0.1% Triton X-100, 20μg/ml PI, 200μg/ml RNase) 600μl로 20분 반응시켰다. 유식 세포분석에서 얻은 정보의 분석은 CellQuest Software (Becton Dickinson)를 이용하였다.
(4) Western blot analysis: MPA를 처리한 Jurkat 세포는 포집하여, 냉 PBS로 2회 세척하였다. 얻어진 세포는 파쇄 용액(50 mM HEPES, pH7.4, 150 mM NaCl, 1% deoxycholate, 1 mM EDTA, 1 mM PMSF, 1 μg/ml aprotinin)과 4oC에서 30분 반응시켰다. 동량의 세포 파쇄액(200μg)과 2X sample buffer를 혼합하여 100oC에서 3분 가열하여 단백질의 변성 을 유도한 후에 12.5% 및 15% SDS-polyacrylamide gel elec- trophoresis (SDS-PAGE)를 시행하였다. 전기 영동이 끝난 gel의 단백질은 electrotransfer system (Ellard Inc, Seattle, WA,
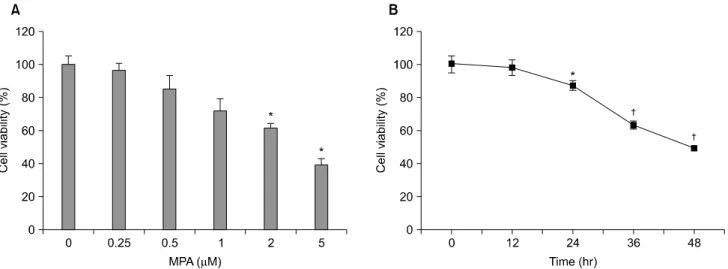
Fig. 1. MPA decreased the viability of Jurkat cells in a dose- and time-dependent manners. Cells are treated with various concentrations of MPA for 48 hrs (A) or various periods (B) and then, viability is measured by MTT assay after MPA treatment. Data represent the mean±standard deviation (S.D.) of quadruplicates. *P<0.05, †P<0.01 by Student's t-test, compared to control group.
USA)을 이용(0.8 mA/cm)하여 nitrocellulose membrane으로 이동시키고, blocking buffer (5% skim milk)와 상온에서 2시 간 반응시켰다. Caspase-3, Bcl-2, Bax 및 β-actin에 대한 항 체는 0.05% (v/v)의 tween-20이 함유된 Tris-buffered saline (TBS-T)에 1:1,000으로 희석하여 nitrocellulose membrane과 상온에서 2시간 반응시켰으며, 각 항체에 대한 이차항체 anti-rabbit IgG conjugated horse radish peroxidase (HRP)와 anti-mouse IgG conjugated HRP는 TBS-T로 희석(1:3,000)하 여 상온에서 1시간 반응시킨 후, enhanced chemillumine- scence (ECL) kit (Amersham, England)를 이용하여 현상하였 다.
(5) 세포 내 활성 산소 생성(reactive oxygen species)의 측 정: MPA에 의한 세포 내 활성화 산소의 생성을 측정하기 위하여 형광 probe 2',7'-dichlorofluorescein diacetate (DCF-DA;
Sigma)를 이용하였다. 비형광 물질인 DCF-DA는 세포 내 hydrogen peroxide (H2O2)와 관련된 peroxides 존재 시 형광의 DCF로 변환되어 녹색의 형광을 발한다. Jurkat 세포에 MPA 를 처리한 후 세포를 수확하기 전에 5μM DCF-DA를 처리 하여 37oC에서 30분 배양하였다. 배양한 세포는 PBS (pH 7.4)로 세척하여 수확하고 flow cytometry (FACSCalibur, BD Biosciences)로 형광을 측정하고 CellQuest Software (Becton Dickinson)를 이용하여 분석하였다.
(6) 통계처리: 표시된 결과는 4번의 독립적인 실험 결과이 며, 실험 결과의 통계 처리는 student's t-test에 준하여 처리 하였고, P-value가 0.05 (P<0.05) 이하인 경우를 유의한 것
으로 판정하였다.
결 과
1) MPA가 T 림프구, Jurkat 세포의 세포 생존율에 미치 는 영향
MPA의 Jurkat 생존에 대한 영향을 조사하기 위하여 MPA 를 다양한 농도로 48시간 처리한 후, 세포 활성도를 MTT 방법으로 조사하였다. MPA 0.25μM 농도에서 세포 생존율 은 96%, 0.5μM 농도에서는 85%, 1μM 농도에서는 72%, 2μM 농도에서는 61%의 세포 생존율을 보였으며, 5μM의 농도에서는 대조군에 비교하여 39%의 세포 생존율을 보였 다(Fig. 1A). 또한, MPA에 의한 Jurkat 세포 죽음의 시간의 의존적인 변화를 확인하기 위하여 5μM 농도의 MPA를 처 리한 후 시간별로 세포 생존율을 조사하였다. Jurkat 세포의 생존율은 MPA 처리 24시간 후부터 감소하기 시작하여, 48 시간 이후에는 약 40%로 유의한 세포 생존율의 감소를 보 였다(Fig. 1B). 이상의 결과는 MPA에 의한 세포 독성이 Jurkat 세포에서 농도 및 시간 의존적으로 축적되어 증가함 을 시사하였다.
2) MPA에 의한 Jurkat 세포 죽음의 성격 규명 Jurkat 세포에서 MPA에 의한 세포 죽음이 세포 자멸사 기전에 의해서 매개될 가능성을 조사하기 위하여, 다양한 농도의 MPA를 48시간 처리한 세포의 DNA를 PI 염색 후
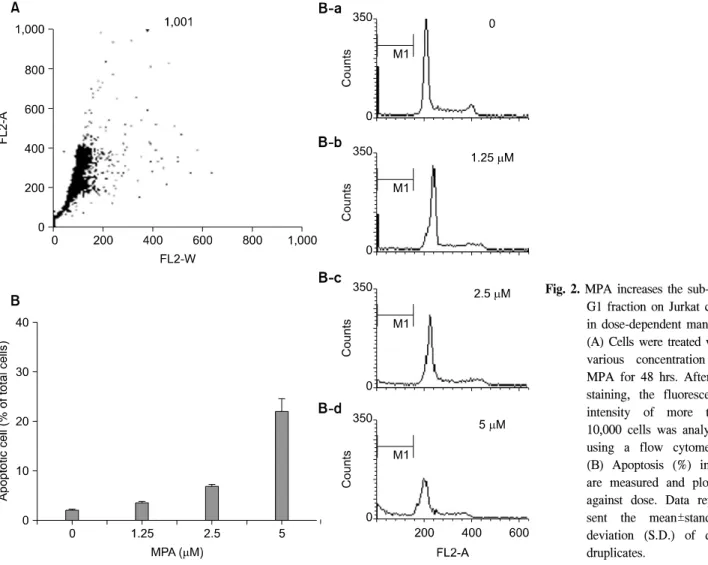
Fig. 2. MPA increases the sub-G0/
G1 fraction on Jurkat cells in dose-dependent manner.
(A) Cells were treated with various concentration of MPA for 48 hrs. After PI staining, the fluorescence intensity of more than 10,000 cells was analyzed using a flow cytometry.
(B) Apoptosis (%) in A are measured and plotted against dose. Data repre- sent the mean±standard deviation (S.D.) of qua- druplicates.
유식세포 분석기(Flow cytometry)를 이용하여 세포 자멸사 정도를 조사하였다. 1.25μM의 MPA를 48시간 처리한 Jurkat 세포의 sub-G0/G1 분획은 대조군 2.06%에 비해 3.86%, 2.5μM의 MPA 처리군에서는 7.49%, 그리고 5μM 의 MPA 처리군에서는 24.61%로 농도 의존적으로 점차 증 가하였다(Fig. 2).
3) MPA에 의한 Jurkat 세포 자멸사에서 caspase-3 pro- teases의 활성 변화
세포 자멸사의 특징적인 현상을 초래하는 세포 내 신호 전달 기전에서 caspase proteases의 중요성은 이미 잘 알려져 있다. MPA에 의한 Jurkat 세포 자멸사 현상이 caspase pro- teases의 활성과 관계가 있는지를 확인하기 위하여 caspase-3 의 발현 변화를 Western bolt 분석법으로 확인하였다. Caspase- proteases는 상방에 위치하는 효소에 의하여 pro form이 절 단되어 복합체를 형성하여 활성화 되며, 이는 하방 caspase
proteases를 활성화 시킨다. MPA 처리 후 caspase-proteases의 활성을 조사하기 위하여 각 pro-caspase proteases의 항체를 이용하였다. 12시간 간격으로 조사한 결과, MPA 처리 36시 간부터 procaspase-3 protease가 분절되어 활성화된 형태로 발현되었다(Fig. 3). 이때 단백질량은 β-actin을 통하여 동 일한 양임을 확인하였다. 이상의 결과는 MPA에 의한 Jurkat 세포의 세포 자멸사 효과는 caspase-3 protease의 활성과 관 련된 세포 자멸사 신호 전달 경로에 의함을 시사하였다.
4) MPA에 의한 Jurkat 세포 자멸사에서 세포 내 활성 산소 생성(reactive oxygen species)에 미치는 영향 MPA에 의한 Jurkat 세포의 자멸사가 H2O2에 의한 세포 자멸사임을 밝히기 위하여 2', 7'-dichlorofluorescin diacetate (DCF-DA) 염색으로 세포 내 H2O2의 생성을 flow cytometry (FACS)를 통하여 조사, 분석하고 M2 값으로 표시하였다.
그 결과, 5μM 농도의 MPA 처리 시 대조군인 48.8%에 비
Fig. 3. Increased caspase-3 proteases expression on MPA treated- Jurkat cells in a time-dependent manner. Cells were treated with MPA for various periods. Cell lysates are separated on 15% SDS-PAGE and immunoblotted for anti- procaspase-3 protease and β-actin. The immunoreactive signals are visualized by ECL detection kit.
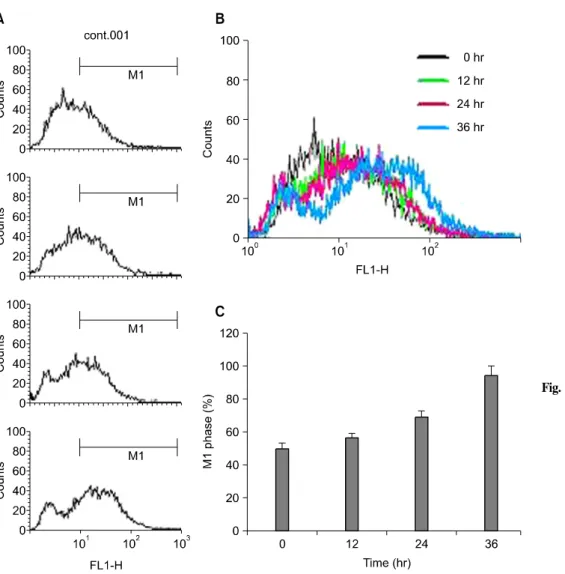
Fig. 4. Increased generation of H2O2 in MPA treated Jurkat cells.
Cells were treated with 5μM MPA for various periods.
Then, cells were incubated with the dye 2', 7'-dichloro- fluorescin diacetate (5μM) and the fluorescence intensity of more than 10,000 cells was analyzed using a flow cytometry.
하여, 12시간에 56.2%, 24시간에 68.1%, 48시간에 94.0%로 시간 의존적인 증가를 보였다(Fig. 4). 이상의 결과는 MPA 에 의한 세포 내 활성 산소인 H2O2 생성이 세포 자멸사에
기여하였을 가능성을 시사하였다.
5) MPA에 의한 Jurkat 세포 자멸사에서 Bcl-2 및 Bax 단백질의 발현에 미치는 영향
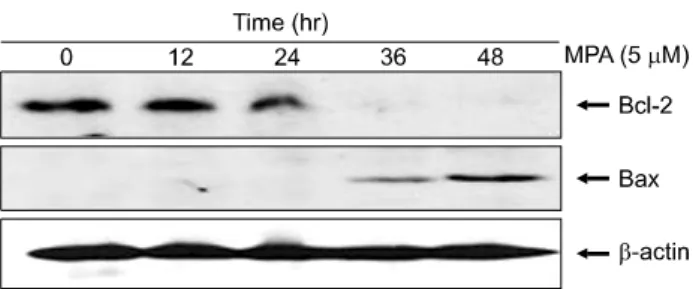
MPA 처리 시 Bcl-2 및 Bax 단백질 발현을 알아보기 위하 여 Jurkat 세포에 5μM의 MPA 처리 후 Western blot을 실시 하였다. 그 결과 세포 자멸사 억제 단백질인 Bcl-2의 발현 양상은 MPA 처리 24시간 후부터 현저하게 감소하였으며, 세포 자멸사 촉진 단백질인 Bax는 MPA 처리 36시간부터 증가하였다(Fig. 5).
고 찰
MMF는 inosine을 guanosine nucleotide로 전환시키는 IMPDH의 활성을 억제, T 림프구와 B 림프구의 증식을 선 택적으로 억제하며,(12) 급성 거부 반응의 예방과 스테로이
Fig. 5. Decreased expression of Bcl-2 and incresed Bax protein in MPA treated Jurkat cells. Cells were treated with 5 μM MPA for various periods. The equal amounts of protein from cell lysate were subjected on 15% SDS-PAGE, transferred onto nitrocellulose membrane and immuno- blotted with anti-Bcl-2, anti-Bax and anti-β-actin anti- bodies. The immunoreactive signals were visualized by ECL detection kit.
드 요법에 반응하지 않는 급성 거부 반응에 효과적이 다.(13) IMPDH는 type I, II 두 가지 isoform이 있는데, 활성 화된 림프구는 주로 type II에 민감하게 반응한다.(14) MPA 에 의한 GTP의 고갈은 림프구 증식 억제와 세포막 당단백 인 fucose나 mannose 전달을 차단한다. 이로 인해 세포 유착 분자는 이들 세포가 혈관 내피 세포에 부착하는 것을 저해 하고 염증 부위로의 이동을 감소시킨다.
본 연구 결과에서 MPA에 의한 세포 독성 및 세포 자멸사 효과는 Cohn 등(15)의 사람 림프구인 Molt 세포에서 보고한 결과와 일치하였다. MPA는 Jurkat 세포의 생존율을 농도 의 존적 및 시간 의존적으로 저하시켰으며, caspase-3의 활성화 를 유도하였다. 또한 sub-G0/G1 분획의 증가와 핵산의 분절 현상으로 미루어 세포 자멸사 기전에 의해서 매개됨을 확 인 할 수 있었다.
활성 산소종은 활성화된 호중구, xanthine oxidase, 미토콘 드리아 호흡 및 아라키돈산 대사 과정을 통해서 생성된 다.(9) 비록 superoxide dismutase, catalase, glutathione pero- xidase 효소 등이 세포 내에서 생성된 활성 산소종을 제거하 기 위한 생리적 방어 기전으로 작용하지만, 과도한 활성 산 소종은 이들 방어 체계를 초과하여 세포 손상을 초래한 다.(10)
본 연구에서 MPA에 의한 Jukat 세포 자멸사는 세포 내 활성산소종인 H2O2의 생성 증가를 초래하였다. 특히, 세포 내 H2O2는 48시간에 최대의 생성량 증가를 보였다.
세포 자멸사를 조절하는 Bcl-2 family 단백질은 사람의 림 프종(human follicular lymphoma)과 관련된 유전자로서 크게 세포 자멸사 촉진 단백질과 세포 자멸사 억제 단백질로 구
분되어진다.(16) 이 중 Bcl-2는 세포 자멸사를 억제하고, Bax 단백질은 세포 자멸사를 촉진한다.(17) 또한, Bcl-2 family 단백질은 c-terminal이 소수성 막으로 구성되어 미토콘드리 아의 막에 결합할 수 있는데,(18) 다양한 자극 시 Bcl-2나 Bcl- XL의 과발현을 통해 미토콘드리아 막전위(MPT,ㆍΨ m)의 상실과 cytochrome c의 방출을 방지하였다.(19) 반대로 Bax나 Bcl-XS 단백질을 과발현 시켰을 경우에는 cytochrome c의 방출을 촉진하였고, 미토콘드리아 막의 탈전극을 유도 한다.(20) 또한 미토콘드리아의 intermembrane space에 존재 하여 respiratory chain에서의 전자전달자의 역할을 수행하 는,(21) cytochrome c는 세포자멸사 자극에 의하여 세포질로 방출되어 Apaf-1, dATP, procaspase-9과 복합체를 형성하여 caspase-9을 활성화 한다. 이러한 cytochrome c는 아직 충분 히 밝혀져 있지 않지만 Bcl-2의 방출을 억제하고 Bax의 방 출을 촉진한다.(22) 활성화된 caspase-9은 다음 단계의 caspase-3를 활성화 한다.(16) 본 연구 결과에서는 MPA는 Bcl-2의 발현감소와 Bax의 발현 증가 혹은 그에 따른 비율 의 감소를 초래하고 caspase-3를 활성화하여 Jurkat 세포의 세포 자멸사를 촉진하였을 것으로 판단된다. 특히 이러한 세포 자멸사 현상은 MPA가 Jurkat 세포에서 활성산소의 생 성을 유도하여 초래하였다고 판단된다.
결 론
MPA에 의한 세포 독성이 Jurkat 세포에서 농도 및 시간 의존적으로 축적되어 증가함을 시사하였고, MPA에 의한 Jurkat 세포 내 활성 산소 생성을 증가시키고 세포 자멸사 억제 단백질인 Bcl-2의 발현을 감소, 세포 자멸사 촉진 단백 질인 Bax는 증가시킴으로 인하여 세포 자멸사에 기여하였 을 가능성을 시사하였다.
REFERENCES
1) Allison AC, Eugui EM. Immunosuppressive and other effects of mycophenolic acid and an ester prodrug, mycophenolate mofetil. Immunol Rev 1993;136:5-28.
2) Allison AC, Kowalski WJ, Muller CD, Eugui EM. Mecha- nisms of action of mycophenolic acid. Ann N Y Acad Sci 1993;696:63-87.
3) Allison AC, Eugui EM. Mycophenolate mofetil and its mecha- nisms of action. Immunopharmacology 2000;47:85-118.
4) Natsumeda Y, Carr SF. Human type I and II IMP dehydro- genases as drug targets. Ann N Y Acad Sci 1993;696:88-93.
5) Allison AC, Eugui EM. Purine metabolism and immunos- uppressive effects of mycophenolate mofetil (MMF). Clin Transplant 1996;10:77-84.
6) Gerber DA, Bonham CA, Thomson AW. Immunosuppressive agents: recent developments in molecular action and clinical application. Transplant Proc 1998;30:1573-9.
7) Cohen JJ. Apoptosis. Immunol Today 1993;14:126-30.
8) Cheon SH, Sung NH. Apoptosis, Genetics Vol.2. Seoul: World Science Co; 1998.
9) Xiong Y, Zhang H, Beach D. Subunit rearrangement of the cyclin-dependent kinases is associated with cellular transfor- mation. Genes 1993;7:1572-83.
10) Gao Z, Huang K, Xu H. Protective effects of flavonoids in the roots of Scutellaria baicalensis Georgi against hydrogen peroxide-induced oxidative stress in SH-SY5Y cells. Phar- macol Res 2001;43:173-8.
11) Halliwell B, Gutteridge J.M.C. Protection aganist oxidants in biological systems: the superoxide theory of oxygen toxicity.
In Textbook of Surgery: Free radicals in biology and medicine.
2nd Ed. Oxford: Clarendon Press; 1989. p.86-9.
12) Lee WA, Gu L, Miksztal AR, Chu N, Leung K, Nelson PH.
Bioavailability improvement of mycophenolic acid through amino ester derivatization. Pharm Res 1990;7:161-6.
13) Sollinger HW. Mycophenolate mofetil for the prevention of acute rejection in primary cadaveric renal allograft recipients.
U.S. Renal Transplant Mycophenolate Mofetil Study Group.
Transplantation 1995;60:225-32.
14) Nagai M, Natsumeda Y, Konno Y, Hoffman R, Irino S, Weber G. Selective up-regulation of type II inosine 5'-monophosphate dehydrogenase messenger RNA expression in human leukemias.
Cancer Res 1991;51:3886-90.
15) Cohn RG, Mirkovich A, Dunlap B, Burton P, Chiu SH, Eugui E, et al. Mycophenolic acid increases apoptosis, lysosomes and lipid droplets in human lymphoid and monocytic cell lines.
Transplantation 1999;68:411-8.
16) Gross A, McDonnell JM, Korsmeyer SJ. Bcl-2 family members and the mitochondria in apoptosis. Genes Dev. 13:1899-911, 1999. Vander Heiden, M.G., and Thompson, C.B. Bcl-2 proteins: inhibitors of apoptosis or regulators of mitochondrial homeostasis? Nat Cell Biol 1999;1:E209-16.
17) Kroemer G, Dallaporta B, Resche-rigon M. The mito- chondrial death/life regulator in apoptosis and necrosis. Nature Med 1998;6:614-20.
18) Reed JC. Bcl2 and the regulation of programmed cell death.
J Cell Biol 1994;124:1-6.
19) Herr I, Wilhelm D, Bohler T. Activation of CD95 (APO-1/
Fas) signaling by ceramide mediates cancer therapy-induced apoptosis. EMBO 1997;16:6200-8.
20) Li PF, Dietz R, Von Harsdorf R. P53 regulates mitochondrial membrane potential through reactive oxygen species and induces cytochrome c-independent apoptosis blocked by Bcl-2.
EMBO J 1999;18:6027-36.
21) Heibein, J.A., Barry, M., Motyka, B. Granzyme B-induced loss of mitochondrial inner membrane potential (Delta Psi m) and cytochrome c release are caspase independent. J Immuno- logy 1999;163:4683-93.
22) Lemos FB, Ijzermans JN, Zondervan PE. Differential expre- ssion of heme oxygenase-1 and vascular endothelial growth factor in cadaveric and living donor kidneys after ischemia- reperfusion. J Am Soc Nephrol 2003;14:3278-87.