서 론
폐암은 전세계적으로 사망률이 가장 높은 암종으로서 우리나라 역시 2014년 기준 전체 암으로 인한 사망 중 폐암으로 인한 사망 이 28%를 차지하였다. 그 동안 암의 진단 및 치료기술이 현저히 발전해왔음에도 불구하고 폐암은 조기진단이 어려울 뿐만 아니라 5 년 생존율이 20% 수준에 그쳐 새로운 치료대책이 절실히 요구되는 실정이다1-3). 한의학적으로 폐암은 전신적인 正氣와 陰液 부족 등 虛證을 근본으로 痰飮과 瘀血이 생겨 형성되는 것으로 알려져 있 으며, 실제 중의학에서는 폐암을 치료하는 데 황기와 사삼을 주요
약재로 한 益氣養陰法을 적극적으로 운용해왔다4,5). 황기와 사삼의 항암작용에 대한 전임상연구들을 살펴보면 황기는 면역증강과 신생 혈관억제를 통해 항암활성을 가지며6-10), 사삼은 암세포의 apoptosis 유도, 비장세포 증식유도 및 혈관신생억제 등을 통해 항 암작용을 나타낸다11-15). 이에 본 저자는 인체폐암세포에서 황기와 사삼이 항암 시너지효과를 가지는지 여부를 조사하여 유의한 결과 를 보고하였으나16), 그 기전은 명확히 밝혀지지 않았다.
Apoptosis는 일정한 프로그램에 따라 세포가 능동적으로 죽는 과정으로서 세포막의 blebbing 현상, 세포질과 염색질의 응축, DNA의 절편화 등을 특징으로 한다. Necrosis가 일어난 세포는 팽
H1299 인체폐암세포주에서 활성산소종 생성에 의한 황기와 사삼의 항암 시너지 작용
민태린·박현지·박신형*
동의대학교 한의과대학 병리학교실 & 동의대학교 한의학연구소
Reactive Oxygen Species (ROS) Generation Contributes to the Synergistic Anticancer Effect of Astragalus Membranaceus and Adenophora Triphylla
Var. Japonica in H1299 Human Lung Carcinoma Cells
Tae Rin Min, Hyun Ji Park, Shin Hyung Park*
Department of Pathology & Research Institute of Korean Medicine, Dong-eui University
This study was designed to investigate the mechanism of the synergistic anticancer effect of
Astragalus membranaceus
(AM) andAdenophora triphylla var. japonica
(AT) in H1299 human lung carcinoma cells. A combined treatment of ethanol extract of AM (EAM) and AT (EAT) explosively increased the reactive oxygen species (ROS) generation in H1299 cells compared to the single treatment of each of them. Co-treatment of N-acetyl-L-cysteine (NAC) with EAM and EAT markedly enhanced the cell viability and suppressed apoptosis in H1299 cells, suggesting that ROS generation contributed to the anticancer effect of EAM and EAT. Interestingly, the combined treatment of EAM and EAT down-regulated p-AKT in H1299 cells, which was abrogated by NAC treatment. These results clearly indicated that ROS generation mediated the inactivation of AKT. Co-treatment of LY294002 with EAM and EAT significantly reduced the cell viability at a concentration which EAM and EAT didn’t show any cytotoxicity. In addition, the recovery of cell viability by co-treatment of NAC with EAM and EAT was quite reversed by LY294002 treatment, which confirmed that the inactivation of AKT played a pivotal role in ROS-mediated apoptosis. Taken together, our results demonstrated that the synergistic anticancer effect of EAM and EAT was mediated by ROS generation and inactivation of AKT. We provide a valuable preclinical data for the development of more effective combination of AM and AT to treat lung cancer.keywords :
Astragalus membranaceus
,Adenophora triphylla var. japonica
, Combination therapy, Lung cancer, ROS generation
* Corresponding author
Shin Hyung Park, Department of Pathology, College of Korean Medicine, Dong-eui University, 47227, YangJeong-ro, Busanjin-gu, Busan, Republic of Korea
·E-mail : [email protected] ·Tel : +82-51-850-8646
·Received : 2017/12/15 ·Revised : 2018/05/24 ·Accepted : 2018/06/29
ⓒ The Society of Pathology in Korean Medicine, The Physiological Society of Korean Medicine pISSN 1738-7698 eISSN 2288-2529 http://dx.doi.org/10.15188/kjopp.2018.06.32.3.157
Available online at https://kmpath.jams.or.kr
창하여 터짐으로써 주변 조직에 염증을 일으키는 반면 apoptosis 가 일어난 세포는 식세포의 식작용에 의해 제거되므로 주위에 염증 반응을 일으키지 않는다17,18). 암 치료제 개발에 있어서 암세포에 효율적으로 apoptosis를 일으키는 물질을 탐색하고 그 기전을 밝 히는 것이 중요하나, 현재 사용되고 있는 항암제들은 인체에 대한 독성과 내성 출현 등으로 인해 한계를 드러내고 있는 실정이다19). 따라서 새로운 항암물질을 탐색할 필요성이 대두되며, 이에 인체에 오랫동안 사용한 경험을 바탕으로 안전성과 효과가 검증된 천연물 이나 한약재가 그 후보물질로서 각광받고 있다. 특히 한약에 포함 된 다양한 성분 간의 combination이 단일성분보다 더욱 우수한 효과를 가짐이 보고되면서 단일성분을 주축으로 한 항암제나 표적 항암치료가 한계를 드러내는 현 시점에 한약재 간의 병용효과와 그 기전을 밝히는 연구가 매우 유망한 치료전략이 될 수 있다20-22). Apoptosis를 일으키는 다양한 인자 중에 활성산소종(reactive oxygen species, ROS)을 들 수 있다. 활성산소종은 산소이온과 과산화수소 등을 포함한 화학적으로 반응성이 높은 분자로서 산소 의 정상적인 대사과정에서 생기며, 세포 내 신호전달 및 항상성 유 지에 중요한 역할을 한다. 그러나 자외선이나 고온 환경에 노출되 어 세포 내 활성산소종이 급격히 증가하게 되면 산화적 스트레스로 인해 세포구조를 손상시키면서 apoptosis를 일으킬 수 있다23). 특 히 다양한 천연물 성분이 암세포에 활성산소종을 유발함으로써 항 암활성을 가진다고 알려져 있다. 예를 들어 capsaicin은 췌장암세 포에24), curcumin은 위암세포에25), phenethyl isothiocyanate은 만성골수성백혈병세포에26), piperine은 악성흑색종세포27), deguelin은 폐암세포에 각각 활성산소종을 유발하여 apoptosis를 일으킨다고 보고되어 있다28). 뿐만 아니라 활성산소종을 유발하는 천연물질들은 암세포의 세포주기조절과 전이 억제 및 기존 항암치 료의 효과를 높임으로써 항암활성을 나타낸다29-31).
본 연구는 H1299 인체 폐암세포주에서 황기와 사삼의 항암 시 너지작용을 보고한 기존 연구16)를 토대로 그 기전을 분석한 결과 활성산소종에 의한 AKT의 인산화 감소 및 apoptosis 유발로 인한 것임을 밝혔다. 이러한 결과는 향후 폐암 치료제로서 황기와 사삼 을 보다 효과적으로 활용할 수 있는 전략을 세우는데 유용한 자료 가 될 것으로 사료되어 이에 보고하는 바이다.
재료 및 방법
1. 재료
본 실험에 사용된 황기 에탄올 추출물(ethanol extract of
Astragali membranaceus
, EAM) 과 사삼 에탄올 추출물(ethanol extract ofAdenophora triphilla var. japonica
, EAT)은 Lee의 연구16)에 기재된 방식으로 제조하였다. 세포 배양에 사용된 RPMI 1640, fatal bovine serum (FBS), penicillin-streptomycin (PS), phosphate-buffered saline (PBS), Trypsin-EDTA는 WelGENE (Seoul, Korea)에서 구입하였다. 단백질 분석을 위한 phospho-AKT 항체는 Cell Signaling Technology (Beverly, MA, USA)에서 구입하였고, 나머지 1차 항체와 2차 항체는 모두 Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA)에서 구입하였다. 별도 표기 하지 않은 모든 시약은 Sigma (St. Louis, MO)에서 구입하였다.
2. 세포 배양
본 연구에 사용된 H1299 인체 폐암세포주는 서울대학교 약학 대학 이호영교수님으로부터 분양받았다. 세포 배양 시 RPMI 1640 을 90%, FBS를 10%, PS를 1%가 되도록 혼합하여 사용하였으며, 세포는 37℃, 5% CO2 조건 하에서 배양하였다.
3. MTT assay
H1299 세포를 2.5×103 cells/well이 되도록 96 well plate에 seeding한 후 overnight으로 안정화시킨 다음 EAM과 EAT를 제 시된 농도로 처리하였다. Inhibitor 실험으로서 NAC과 LY294002 를 처리할 경우 전처리나 후처리하지 않고 EAM, EAT와 동시에 처리하였다. 72 시간 배양 후 tetrazolium bromide salt (MTT)를 0.5 ㎍/㎖ 농도가 되도록 분주하고 2 시간 반응시켰다. 그 후 상층 액을 모두 제거하고 DMSO 100 ㎕를 넣어 각 well의 formazan을 녹인 후 microplate reader (Molecular Devices, Sunnyvale, CA, USA)에서 540 nm로 흡광도를 측정하였다.
4. Trypan blue exclusion assay
H1299 세포를 3×104 cells/well 농도로 12 well plate에 seeding한 후 overnight으로 안정화시키고 EAM (500 ㎍/㎖), EAT (500 ㎍/㎖), NAC (10 mM)을 동시 처리하여 72 시간 배양 하였다. 그 후 세포를 모아 0.1%로 희석된 trypan blue 용액에 부 유하여 hematocytometer로 세포수를 측정하였다. 푸른색으로 염 색된 세포는 dead cell로 판단하여 측정에서 제외하였고, 염색되지 않은 세포만 live cell로 판단하여 측정하였다.
5. 활성산소종 생성 측정
6 well plate에 H1299 세포를 5×104개 seeding한 후 overnight으로 안정화시키고 EAM (500 ㎍/㎖)과 EAT (500 ㎍/
㎖)를 시간별로 처리한 후 배양액에 5-(and 6)-carboxy-2’7’- dichlorodihydrofluorescein diacetate (H2DCFDA)를 10 μM 처 리하여 20분간 37℃, 5% CO2 조건에서 배양시켰다. 염색된 세포 들을 모아 PBS (pH 6.9) 500 ㎕에 부유하여 DNA flow cytometer (BD FACSCalibur)와 CellQuest software를 사용하여 상대적인 활성산소종 생성 변화를 관찰하였다.
6. Cell cycle 분석
60Φ dish에 H1299 세포를 1×105개 seeding한 후 overnight 으로 안정화시키고 EAM (500 ㎍/㎖), EAT (500 ㎍/㎖), NAC (5 mM)을 동시 처리하였다. 72 시간 후 세포들을 모아 80% 에탄올로 4 ℃에서 1 h 고정하였다. 원심분리한 세포를 500 ㎕의 propidium iodide solution (450 ㎕ PBS, 50 ㎕ 0.5 ㎎/㎖
propidium iodide, 1.5 ㎕ 10 ㎎/㎖ RNase A)에 부유하여 30분 간 핵을 염색하였다. 그 후 원심분리시켜 상층액을 제거한 후 500
㎕ PBS (pH 6.9)에 부유하여 DNA flow cytometer와 CellQuest
software를 사용하여 cell cycle을 histogram으로 분석하였다.
7. Annexin V/PI double staining assay
6 well plate에 H1299 세포를 5×104개 seeding한 후 overnight으로 안정화시키고 EAM (500 ㎍/㎖), EAT (500 ㎍/㎖), NAC (5 mM)을 동시 처리하였다. 72 시간 후 H1299 세포들을 모 아 Annexin V‑FITC apoptosis detection kit (PharMingen, San Diego, California)를 이용하여 제조사의 매뉴얼에 따라 염색 하였다. 15분간 암실에서 세포를 염색한 후 DNA flow cytometer 와 CellQuest software를 사용하여 형광을 분석하였다. Annexin V‑positive cell을 apoptotic cell로 해석하여 수치를 분석하였다.
8. Western blotting
H1299 세포에 EAM (500 ㎍/㎖), EAT (500 ㎍/㎖), NAC (5 mM)을 48 시간 처리한 후 세포를 모아 냉장된 lysis buffer [RIPA buffer (Thermo Scientific, Rockford, IL, USA) + 1X protease inhibitor cocktail (Thermo) + phosphatase inhibitors (1 mM Na3VO4, 100 mM NaF, and 10 mM NaPP)]
에 1 시간 용해시킨 후 Bio-rad 단백질 정량 시약을 이용하여 정 량하였다. 그 후 20 ㎍의 총단백질을 SDS-polyacrylamide gel에 loading하여 전기영동시키고, PVDF membrane (Millipore Corporation, Bedford, MA)에 transfer하였다. 3% bovine serum albumin (BSA, MP Biomedicals Europe, Illkirch, France)으로 1 시간 실온에서 blocking한 후 4℃에서 1차 항체 (1:1000)와 overnight incubation하였다. TBST로 세척하고 상온 에서 2차 항체(1:5000)와 1 시간 반응시킨 후 다시 TBST로 세척하 고 Enhanced Chemilunimoecence (ECL) 용액 (Amersham Life Science Corp., Arlington Height, IL, USA)을 뿌려 암실에서 X-ray film (Agfa-Gevaert, Melbourne, Australia)에 감광시켰다.
9. 통계적 분석
모든 실험 결과는 mean±SD로 표시하였으며, 통계적 유의성 검증은 Student’s t-test를 이용하여 P < 0.05인 것을 유의성 있 다고 판단하였다.
결 과
1. EAM-EAT 병용처리가 활성산소종 생성에 미치는 영향
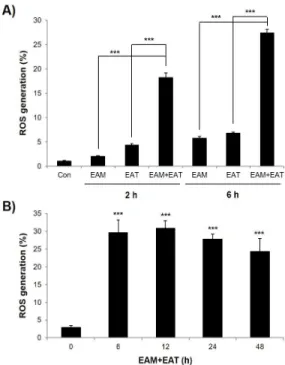
EAM과 EAT의 항암시너지 효과가 활성산소종 발생과 관련있 는지 알아보기 위해 항암시너지 효과가 나타나는 농도16)인 500 ㎍ /㎖에서 EAM과 EAT를 단독 혹은 병용처리한 후 활성산소종 생성 량을 flow cytometry로 조사하였다. 그 결과 EAM과 EAT 단독처 리에 비해 병용처리 시 활성산소종 생성이 급격히 증가하였으며, 시간의존적으로 활성산소종이 증가하여 EAM과 EAT 병용처리 2 h 후보다 6 h 후에 더욱 증가하는 경향을 보였다(Fig. 1A). 이러한 결과는 Lee의 연구16)에서 EAM과 EAT 단독처리에 비해 병합처리 시 apoptosis가 현저히 증가하는 결과와 연관되어 EAM과 EAT의 항암시너지 작용이 활성산소종 발생과 관련있을 가능성을 보여주었
다. 또한 EAM과 EAT를 시간별로 병용처리했을 때 활성산소종은 6 h 후부터 48 h 후까지 지속적으로 발생되어 활성산소종에 의한 signaling pathway가 장시간동안 활성화되었을 것임을 예측할 수 있었다(Fig. 1B).
Fig. 1. Effects of EAM and EAT on ROS generation in H1299 cells. (A) H1299 cells were challenged with single- or combined treatment of EAM (500 ㎍/㎖) and EAT (500 ㎍/㎖) for 2 h or 6 h. (B) H1299 cells were challenged with co-treatment of EAM (500 ㎍/㎖) and EAT (500 ㎍/㎖) for various time periods. (A and B) Cells were stained with H2DCFDA (10 μM) at 37℃ in the dark for 30 min. The cell pellets were washed and resuspended in cold PBS for the evaluation of ROS generation using flow cytometry. Data are expressed as the mean ± S.D. of three independent experiments. Significance was determined by the Student's t-test (*** P <
0.001 vs. respective control).
2. 활성산소종이 EAM-EAT 병용처리에 의한 세포생존율 저하에 미치는 영향
EAM-EAT 병용처리에 의한 세포생존율 저하에 활성산소종이 미치는 영향을 조사하기 위해 EAM-EAT와 함께 항산화제인 NAC 을 처리하였다. 먼저 현미경으로 cell density를 관찰한 결과 EAM-EAT 병용처리에 의해 현저히 감소했던 세포밀집도가 NAC 처리에 의해 다시 증가하는 것을 확인할 수 있었다(Fig. 2A). 또한 MTT assay와 trypan blue exclusion assay로 세포생존율을 측 정한 결과 EAM-EAT 병용처리에 의해 급격히 감소한 세포생존율 이 NAC으로 활성산소종 생성을 억제하자 상당 부분 다시 회복되 는 것을 관찰하였다(Fig. 2B and 2C). 이러한 결과는 활성산소종 이 EAM-EAT 병용처리에 의한 세포생존율 감소에 핵심적인 역할 을 담당함을 의미한다.
3. 활성산소종이 EAM-EAT 병용처리에 의한 apoptosis 유발에 미 치는 영향
다음으로 활성산소종이 EAM-EAT 병용처리에 의한 apoptosis
유발에 미치는 영향을 조사하였다. 먼저 annexin V/PI double staining 후 flow cytometry로 annexin V-positive cell을 분석 하였다. Apoptosis 초기에는 세포막 내부에 위치하는 phosphatidylserine (PS)이 세포막 외부로 이동하여 annexin V와 결합하므로 annexin V+/PI- 상태의 세포가 증가하고, apoptosis 후기에는 세포막에 pore가 형성되면서 PI에 의해 핵이 염색되므로 annexin V+/PI+ 상태의 세포가 증가한다. 따라서 apoptosis가 일 어난 세포는 stage에 상관없이 공통적으로 annexin V가 양성으로 나타난다32). 이를 바탕으로 데이터를 해석한 결과 EAM-EAT 병용 처리에 의해 현저히 증가한 apoptotic cell population이 NAC 처 리에 의해 control 수준으로 감소하는 것을 확인할 수 있었다 (Fig. 3A). Cell cycle 분석결과 역시 같은 패턴을 보여 apoptotic cell을 의미하는 sub-G1 phase cell population이 EAM-EAT 병 용처리에 의해 증가했다가 NAC 처리에 의해 감소하는 경향을 보 였다 (Fig. 3B). 이러한 결과는 EAM-EAT 병용처리에 의한 apoptosis 유발이 활성산소종 생성에 의한 것임을 보여준다.
Fig. 2. Effects of NAC on cell viability in H1299 cells treated with EAM and EAT. H1299 cells were seeded and treated with EAM (500 ㎍/
㎖) and EAT (500 ㎍/㎖) w/ or w/o NAC (10 mM) for 72 h. (A) The cell density was observed under microscope (×50). (B) The cell viability was evaluated by MTT assay. (C) The viable cells were counted by trypan blue exclusion assay. The blue stained cells were regarded as dead cells and the relative cell viability compared to control cells was evaluated. Data are expressed as the mean ± S.D. of three independent experiments.
Significance was determined by the Student's t-test (*** P < 0.001 vs.
respective control).
4. EAM-EAT 병용처리에 의한 활성산소종 생성이 AKT signaling pathway에 미치는 영향
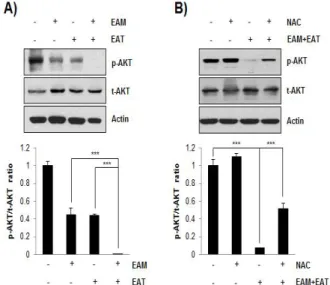
AKT는 암세포의 apoptosis를 억제하여 세포생존에 기여하는 신호분자로서 다양한 암종에서 과활성화되어 있다. 따라서 AKT의 ATP-binding site나 regulatory domain을 조절함으로써 kinase 활성을 억제할 수 있는 약물들이 개발되고 있다33). EAM-EAT 병 용처리에 의한 항암 시너지작용이 AKT signaling pathway 조절 과 관련되어 있는지 확인하기 위하여 웨스턴블롯을 시행한 결과 EAM과 EAT 단독처리 시보다 병용처리 시 AKT의 인산화가 현저
히 줄어드는 것을 확인할 수 있었다(Fig. 4A). 흥미롭게도 EAM-EAT 병용처리에 의해 감소한 AKT의 인산화는 NAC으로 활 성산소종 생성을 차단하자 다시 증가하였다(Fig. 4B). 이를 통해 EAM-EAT 병용처리에 의한 AKT의 활성 감소가 활성산소종 발생 에 의한 것임을 알 수 있었다.
Fig. 3. Effects of NAC on apoptosis induction in H1299 cells treated with EAM and EAT. H1299 cells were seeded and treated with EAM (500
㎍/㎖) and EAT (500 ㎍/㎖) w/ or w/o NAC (10 mM) for 72 h. (A) Annexin V/PI double staining was conducted using a flow cytometer. Annexin V-positive population was determined as apoptotic cells. (B) Cells were stained with PI solution and sub-G1 DNA content was evaluated using a flow cytometer. Data are expressed as the mean ± S.D. of three independent experiments. Significance was determined by the Student's t-test (*** P < 0.001 vs. respective control).
Fig. 4. Effects of EAM and EAT on the phosphorylation of AKT in H1299 cells. (A) H1299 cells were challenged with single- or combined treatment of EAM (500 ㎍/㎖) and EAT (500 ㎍/㎖) for 48 h. (B) H1299 cells were seeded and treated with EAM (500 ㎍/㎖) and EAT (500 ㎍/㎖) w/ or w/o NAC (10 mM) for 48 h. (A and B) Cells were lysed and the cellular proteins were separated by SDS-polyacrylamide gels, transferred onto PVDF membrane. The membrane was probed by the indicated antibodies, and the proteins were visualized by an ECL detection system.
Actin was used as an internal control. The ratio of p-AKT/t-AKT was analysed with Image J software using actin for normalization. Data are expressed as the mean ± S.D. of three independent experiments.
Significance was determined by the Student's t-test (*** P < 0.001 vs.
respective control).
5. EAM-EAT 병용처리에 의한 AKT 인산화 억제가 항암 시너지 작용에 미치는 영향
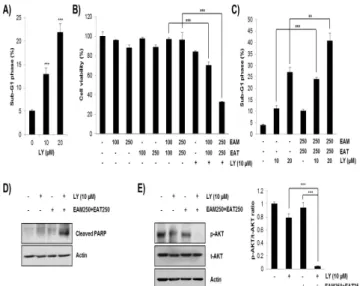
AKT 인산화 억제만으로도 H1299 세포주에 세포증식억제와 apoptosis가 유발되는지 확인하기 위하여 LY294002를 농도별로 처리한 후 cell cycle 분석을 시행하였다. 그 결과 농도의존적으로 H1299 세포주의 sub-G1 phase cell population이 증가하는 것을 확인함으로써 AKT 인산화 억제가 apoptosis를 유발하여 세포증식 을 억제함을 알 수 있었다(Fig. 5A) 다음으로 EAM-EAT 병용처리 에 의해 억제된 AKT의 인산화가 항암 시너지 작용에 미치는 영향 을 조사하기 위해 AKT 저해제인 LY294002를 처리한 후 MTT assay로 세포생존율을 측정하였다. 그 결과 항암 시너지 작용이 유 의하게 나타나지 않았던 농도인 EAM과 EAT 각 100 ㎍/㎖과 250
㎍/㎖ 처리군에서 LY294002를 함께 처리하자 세포생존율이 현저 히 감소하는 것을 확인할 수 있었다(Fig. 5B). 이러한 세포증식억제 가 apoptosis 유발에 의한 것인지 확인하기 위하여 cell cycle 분 석을 시행한 결과 LY294002와 EAM-EAT의 병용처리에 의해 sub-G1 phase cell population이 대폭 증가하였으며(Fig. 5C), 이 와 함께 apoptosis marker 단백질인 cleaved-PARP의 발현이 증 가하는 것을 확인하였다(Fig. 5D). 이러한 결과는 LY294002 단독 처리에 의해서는 약하게 감소하였던 p-AKT의 발현이 LY294002와 EAM-EAT의 병용처리에 의해 현저히 감소하는 결과와 함께 AKT 의 인산화 수준이 H1299 세포주의 apoptosis 유발과 밀접하게 연 관되어 있음을 보여주었다(Fig. 5E). 따라서 EAM-EAT 병용처리가 apoptosis 유발을 통해 항암 시너지 작용을 나타내는 데 AKT의 인산화 억제가 중요한 역할을 함을 알 수 있었다.
Fig. 5. Effects of AKT inhibition on the cell viability in H1299 cells. (A) H1299 cells were treated with various concentration of LY294002 for 72 h.
Cells were stained with PI solution and sub-G1 DNA content was evaluated using a flow cytometer. (B-E) H1299 cells were challenged with indicated concentration of EAM and EAT w/ or w/o LY294002 for 72 h. (B) The cell viability was evaluated by MTT assay. (C) Cells were stained with PI solution and sub-G1 DNA content was evaluated using a flow cytometer. (D) The expression of cleaved-PARP was detected by western blot. Actin was used as an internal control. (E) The expression of p-AKT and t-AKT was detected by western blot. Actin was used as an internal control. The ratio of p-AKT/t-AKT was analysed with Image J software using actin for normalization. Data are expressed as the mean ± S.D. of three independent experiments. Significance was determined by the Student's t-test (** P < 0.01, *** P < 0.001 vs. respective control).
다음으로 EAM-EAT 병용처리에 의한 AKT의 인산화 억제가 활성산소종 생성 및 세포사멸과 관련 있는지 다시 한 번 확인하기 위해 NAC과 LY294002를 함께 처리한 후 MTT assay를 시행하였 다. 그 결과 EAM-EAT 병용처리에 의해 현저히 감소한 세포생존 율이 NAC 처리에 의해 회복되고, 이는 다시 LY294002 처리에 의 해 EAM-EAT 병용처리군 수준으로 감소하는 것을 확인할 수 있었 다(Fig. 5B). 활성산소종 생성 억제에 의해 상승한 세포생존율이 AKT 활성 저해에 의해 다시 감소하는 것은 결국 활성산소종 생성 에 의한 EAM과 EAT의 항암 시너지 작용이 AKT의 활성 억제에 의해 매개됨을 의미하는 것이다.
고 찰
본 연구는 저자들이 앞서 보고한 EAM과 EAT의 항암 시너지 작용이 나타나는 기전을 밝히는 데 주안점을 두었다. 그 결과 EAM 과 EAT 단독처리에 비해 병용처리 시 활성산소종 생성이 현저히 증가함을 관찰하였고(Fig. 1), NAC으로 활성산소종 생성을 차단하 자 EAM-EAT 병용처리에 의한 세포생존율 감소 및 apoptosis 유 발이 저해되는 것을 확인함으로써 EAM과 EAT의 항암 시너지 작 용이 활성산소종 생성에 의한 것임을 알 수 있었다(Fig. 2 and 3).
흥미롭게도 EAM-EAT 병용처리는 AKT의 인산화를 억제하였고, NAC 처리에 의해 p-AKT의 발현이 다시 증가되었으므로 활성산소 종 생성이 AKT의 인산화를 저해함을 알 수 있었다(Fig. 4). 또한 LY294002를 통한 AKT의 활성 억제는 세포생존율을 감소시켰으므 로 EAM과 EAT의 항암 시너지 작용은 활성산소종 생성을 통한 AKT 인산화 억제와 관련 있음을 확인하였다(Fig. 5).
활성산소종은 암의 발전과 진행에 있어 양방면의 작용을 한다 고 알려져 있다. 소량의 활성산소종은 세포의 항상성을 유지하고 세포증식을 일으키며 혈관신생과 암 전이를 촉진하여 암을 악화시 킨다. 반면 과량의 활성산소종은 암세포에 산화적 스트레스를 야기 하여 DNA 손상 및 apoptosis를 일으키므로 암을 억제하는 작용을 한다34). 따라서 암 치료전략을 세울 때 활성산소종의 대조적인 작 용을 매개하는 signaling pathway를 파악하는 것이 중요하다. 본 연구에서는 EAM과 EAT의 병용처리에 의해 발생한 활성산소종이 H1299 인체 폐암세포주에 apoptosis를 일으켜 암을 억제하는 작 용을 했음을 알 수 있으며, 이 과정을 매개하는 신호전달경로가 AKT pathway임을 밝혔다. 기존 연구에 따르면 다양한 drug가 활 성산소종 생성을 통해 AKT 활성을 저해한다고 보고되어 있다. 예 를 들어 대황에 함유되어 있는 emodin은 폐암 세포주에서 활성산 소종을 발생시켜 미토콘드리아의 기능을 억제하는 동시에 AKT와 ERK의 활성을 저해한다35). 계피의 성분인 β-Caryophyllene는 활 성산소종 발생과 PI3K/AKT/mTOR/S6K1 경로 저해를 통해 전립 선암과 유방암 세포에서 apoptosis를 일으킨다고 알려져 있다36). 강황의 주성분인 curcumin은 신장암 세포에서37), 15d-PGJ238)와 2-Methoxyestradiol39)은 공통적으로 혈액암 세포주에서 활성산소 종 생성을 통해 AKT의 불활성화 및 apoptosis를 일으킨다. 또한 histone deacetylase inhibitor와 perifosine이 혈액암 세포주에서 활성산소종 생성 및 AKT 활성 억제를 통해 항암 시너지 작용을
나타낸다는 보고40)도 있어 본 연구의 시너지 기전과 동일한 양상을 보여주었다.
황기와 사삼의 활성산소종 생성에 관한 기존 연구를 살펴보면 황기와 당귀 및 과루인으로 구성된 SH003이 자궁경부암 세포와 전 립선암 세포에서 apoptosis 유발과는 별개로 활성산소종을 발생시 킨다는 보고41,42)가 있다. 이와 반대로 황기의 활성산소종 소거 작 용에 있어서 황기 추출물의 항산화효능이 보고되어 있으며43), 황기 의 유효성분이 골수유래 중간엽줄기세포에서 미토콘드리아의 활성 산소종 축적을 막아 세포의 기능장애 및 apoptosis를 억제한다는 보고가 있다44). 또한 황기의 성분인 astragaloside Ⅳ가 ROS-NFκ B 경로를 저해함으로써 혈관의 기능장애를 회복한다는 보고45), 황 기의 3가지 성분이 산화성 스트레스로 손상받은 간세포를 회복시킨 다는 보고46), 황기의 다당류 성분이 심근세포의 PI3K/AKT 경로를 활성화하여 doxorubicin의 활성산소종 생성으로 인한 심장 독성을 감소시킨다는 보고47) 등은 황기가 오히려 산화적 스트레스를 감소 시켜 세포보호작용을 함을 보여준다. 이는 황기가 활성산소종을 발 생시킬 것인지 소거할 것인지 여부가 대상 세포와 약물처리환경에 따라 달라질 수 있음을 시사한다. 사삼은 항산화작용이 보고48)되어 있을 뿐 암세포에서 활성산소종을 발생시키는지 여부는 아직까지 연구된 바가 없다. 따라서 기존 연구를 종합해보면 황기와 사삼 단 독으로는 활성산소종을 발생시킬 것으로 기대하기 어려우며, 본 연 구의 실험결과 역시 EAM과 EAT 단독처리만으로는 활성산소종 생 성이 미미한 수준으로 관찰되었다. 그러나 EAM-EAT 병용처리에 의해 활성산소종 발생량이 현저히 증가했으며, 이는 apoptosis 유 발로 이어져 EAM과 EAT의 항암 시너지 작용에 기여한 것으로 판 단된다. 다만 NAC 처리에 의해 세포생존율과 apoptosis 유발이 control 수준으로 완벽히 회복되지 않았으므로 EAM과 EAT가 활 성산소종 발생 뿐만 아니라 다른 신호전달경로를 조절하여 이들이 협동적으로 apoptosis를 일으킬 가능성도 존재한다.
본 연구는 황기와 사삼의 항암 시너지 작용의 기전을 밝힘으로 써 향후 폐암을 치료하기 위해 황기와 사삼을 응용할 때 유용한 전 임상 근거를 제공하며, AKT 저해제와 같은 항암제를 함께 투여했 을 때 보다 탁월한 효과를 거둘 수 있을 것임을 제안한다. 그러나 활성산소종이 AKT의 인산화를 억제하는 기전은 추후 더욱 연구되 어야 할 부분으로 사료된다.
결 론
H1299 인체폐암세포주에서 EAM과 EAT의 항암 시너지 효과 가 나타나는 기전을 조사하여 다음과 같은 결론을 얻었다. EAM과 EAT 단독처리에 비해 병용처리 시 H1299 세포의 활성산소종 생성 이 현저히 증가하였다. 활성산소종은 AKT의 인산화를 저해하여 H1299 세포의 apoptosis를 유발하였으므로 EAM과 EAT의 항암 시너지 작용은 활성산소종 생성과 AKT 활성 억제에 의해 매개되는 것으로 판단된다.
이상의 결과는 황기와 사삼의 병용처리에 의해 폐암에 대한 항 암효과를 높일 수 있는 기전을 명확히 제시하며, AKT 저해제와 같 은 항암제와의 combination을 시도할 수 있는 전임상 근거를 제
공한다. 향후 새로운 폐암 치료제 개발에 있어서 황기와 사삼 이외 에 다양한 한약재의 combination effect와 그 기전을 연구해야 할 필요가 있을 것으로 사료된다.
감사의 글
본 연구는 한국연구재단의 일반연구자지원사업(신진연구지원사 업)의 지원에 의해 수행되었음(과제번호 NRF-2016R1C1B2015076).
References
1. Siegel RL, Miller KD, Jemal A. Cancer statistics. CA: A Cancer Journal for Clinicians. 2016;66(1):7-30.
2. National Cancer Information Center. Available from:
http://www.cancer.go.kr/mbs/cancer.
3. Liu LK, Li YF, Wang XX. Exploraion of the Pathogenesis and Therapeutic Methods of Lung Cancer.
China Journal of Basic Medicine In Traditional Chinese Medicine. 2004;10(5):75.
4. Geng G, Sun FY. Efficacy of Yiqi Yangyin in the treatment of advanced non-small cell lung cancer. Nei Mongol Journal of Traditional Chinese Medicine.
1991;4:4-5.
5. Ma CY, Tang YJ. Application of Yiqi Yangyin in lung cancer. Journal of Shandong college of traditional chinese medicine. 1996;20(3):178-9.
6. Chu DT, Wong WL, Mavligit GM. Immunotherapy with Chinese medicinal herbs I. Immune restoration of local xenogeneic graft-versus-host reaction in cancer patients by fractionated Astragalus membranaceus in vitro. J Clin Lab Immunol. 1988;25:119-23.
7. Shao BM, Xu W, Dai H, Tu P, Li Z, et al. A study on the immune receptors for polysaccharides from the roots of Astragalus membranaceus, a Chinese medicinal herb. Biochem Biophys Res Commun.
2004;320(4):1103-11.
8. Li Q, Bao JM, Li XL, Zhang T, Shen XH. Inhibiting effect of Astragalus polysaccharides on the functions of CD4+CD25 highTreg cells in the tumor microenvironment of human hepatocellular carcinoma.
Chin Med J. 2012;125(5):786-93.
9. Auyeung KK, Woo PK, Law PC, Ko JK. Astragalus saponins modulate cell invasiveness and angiogenesis in human gastric adenocarcinoma cells. J Ethnopharmacol.
2012;141(2):635-41.
10. Shao P, Zhao LH, Zhi C, Pan JP. Regulation on maturation and function of dendritic cells by Astragalus mongholicus polysaccharides. Int Immunopharmacol.
2006;6:1161-6.
11. Yu X, Li L, Han CJ, Zhang QZ. Effects of Codonopsis lanceolata total sanponin on apoptosis in HepG2 cells. J Jilin Univ (Medicine Edition). 2011;6:41.
12. So JN, Kim JH. Effects of Codonopsis lanceolata Extracts on bFGF-induced Angiogenesis in vitro. Korean Journal of Biotechnology and Bioengineering.
2003;18(1):25-9.
13. Byeon SE, Lee YG, Cho JY. Regulatory effects of Codonopsis lanceolata on gene expression of GM-CSF in macrophage-like cells. J Ethnopharmacol. 2009;123:185-9.
14. Wang L, Xu ML, Hu JH, Rasmussen SK, Wang MH.
Codonopsis lanceolata extract induces G0/G1 arrest and apoptosis in human colon tumor HT-29 cells involvement of ROS generation and polyamine depletion.
Food Chem Toxicol. 2011;49:149-54.
15. Lee KW, Jung HJ, Park HJ, Kim DG, Lee JY, et al.
Beta-Dxylopyranosyl-(1→3)-beta-D-glucuronopyranosyl echinocystic acid isolated from the roots of Codonopsis lanceolata induces caspase-dependent apoptosis in human acute promyelocytic leukemia HL-60 cells. Biol Pharm Bull. 2005;28:854-9.
16. Lee JM, Kim HJ, Choi YH, Chi GY, Park SH. Synergistic Anticancer Effect of Astragalus Membranaceus and Adenophora Triphylla Var. Japonica in H1299 Human Lung Carcinoma Cells. J Physiol & Pathol Korean Med.
2017;31(2):111-7.
17. Norbury C.J., Hickson I,D. Cellular responses to DNA damage. Annu Rev Pharmacol Toxicol. 2001;41:367-401.
18. Elmore S. Apoptosis: a review of programmed cell death. Toxicol Pathol. 2007 ;35(4):495-516.
19. Skeel RT. Handbook of Cancer Chemotherapy.
Lippincott Willians & Wilkins. 1999a; 34-55.
20. Wang L, Zhou GB, Liu P, Song JH, Liang Y, et al.
Dissection of mechanisms of Chinese medicinal formula Realgar-Indigo naturalis as an effective treatment for promyelocytic leukemia. Proc Natl Acad Sci USA.
2008;105(12):4826-31.
21. Ma XH, Zheng CJ, Han LY, Xie B, Jia J, et al.
Synergistic therapeutic actions of herbal ingredients and their mechanisms from molecular interaction and network perspectives. Drug Discov Today.
2009;14(11-12):579-88.
22. Kim HU, Ryu JY, Lee JO, Lee SY. A systems approach to traditional oriental medicine. Nat Biotechnol.
2015;33(3):264-8.
23. Devasagayam TP, Tilak JC, Boloor KK, Sane KS, Ghaskadbi SS, et al. Free radicals and antioxidants in
human health: current status and future prospects. J Assoc Physicians India. 2004;52:794-804.
24. Zhang R, Humphreys I, Sahu RP, Shi Y, Srivastava SK.
In vitro and in vivo induction of apoptosis by capsaicin in pancreat ic cancer cells is mediated through ROS generation and mitochondrial death pathway. Apoptosis.
2008;13(12): 1465-1478.
25. Zou P, Zhang J, Xia Y, Kanchana K, Guo G, et al. ROS generat ion mediates the ant i-cancer effects of WZ35 via act ivat ing JNK and ER st ress apoptot ic pathways in gastric cancer. Oncotarget. 2015;6(8): 5860-76.
26. Wang Y, Wei S, Wang J, Fang Q, Chai Q. Phenethyl isothiocyanate inhibits growth of human chronic myeloid leukemia K562 cells via react ive oxygen species generation and caspases. Mol Med Rep.
2014;10(1): 543-9.
27. Fofaria NM, Kim SH, Srivastava SK. Piperine causes G1 phase cell cycle arrest and apoptosis in melanoma cells through checkpoint kinase-1 activation. PLoS One.
2014;9(5):e94298.
28. Xu H, Li X, Ding W, Zeng X, Kong H, et al. Deguelin induces the apoptosis of lung cancer cells through regulat ing a ROS driven Akt pathway. Cancer Cell Int, 2015;15:25-33.
29. Zhang J, Zhu X, Li H, Li B, Sun L, et al. Piperine inhibits proliferat ion of human osteosarcoma cells via G2/M phase arrest and metast asis by suppressing MMP-2/-9 expression. Int Immunopharmacol.
2015;24(1):50-8.
30. Tak JK, Lee JH, Park JW. Resveratrol and piperine enhance radiosensit ivity of tumor cells. BMB Rep.
2012;45(4):242-6.
31. Denis I, Cellerin L, Gregoire M, Blanquart C. Cisplatin in combination with Phenethyl Isothiocyanate (PEITC), a potential new therapeutic strategy for malignant pleural mesothelioma. Oncotarget. 2014;5(22):11641-52.
32. van Engeland M, Nieland LJ, Ramaekers FC, Schutte B, Reutelingsperger CP. Annexin V-affinity assay: a review on an apoptosis detection system based on phosphatidylserine exposure. Cytometry. 1998;31(1):1-9.
33. Franke TF, Hornik CP, Segev L, Shostak GA, Sugimoto C. PI3K/Akt and apoptosis: size matters. Oncogene.
2003;22(56):8983-98.
34. Kardeh S, Ashkani-Esfahani S, Alizadeh AM.
Paradoxical action of reactive oxygen species in creation and therapy of cancer. Eur J Pharmacol.
2014;735:150-68.
35. Su YT, Chang HL, Shyue SK, Hsu SL. Emodin induces
apoptosis in human lung adenocarcinoma cells through a reactive oxygen species-dependent mitochondrial signaling pathway. Biochem Pharmacol. 2005;70(2):229-41.
36. Park KR, Nam D, Yun HM, Lee SG, Jang HJ, et al. β -Caryophyllene oxide inhibits growth and induces apoptosis through the suppression of PI3K/AKT/mTOR/S6K1 pathways and ROS-mediated MAPKs activation. Cancer Lett. 2011;312(2):178-88.
37. Woo JH, Kim YH, Choi YJ, Kim DG, Lee KS, et al.
Molecular mechanisms of curcumin-induced cytotoxicity: induction of apoptosis through generation of reactive oxygen species, down-regulation of Bcl-XL and IAP, the release of cytochrome c and inhibition of Akt. Carcinogenesis. 2003;24(7):1199-1208.
38. Shin SW, Seo CY, Han H, Han JY, Jeong JS, et al.
15d-PGJ2 induces apoptosis by reactive oxygen species-mediated inactivation of Akt in leukemia and colorectal cancer cells and shows in vivo antitumor activity. Clin Cancer Res. 2009;15(17):5414-25.
39. Gao N, Rahmani M, Dent P, Grant S. Oncogene.
2-Methoxyestradiol-induced apoptosis in human leukemia cells proceeds through a reactive oxygen species and Akt-dependent process. 2005;24(23):3797-809.
40. Rahmani M, Reese E, Dai Y, Bauer C, Payne SG, et al.
Coadministration of histone deacetylase inhibitors and perifosine synergistically induces apoptosis in human leukemia cells through Akt and ERK1/2 inactivation and the generation of ceramide and reactive oxygen species.
Cancer Res. 2005;65(6):2422-32.
41. Lee KM, Lee K, Choi YK, Choi YJ, Seo HS, et al.
SH003‑induced G1 phase cell cycle arrest induces apoptosis in HeLa cervical cancer cells. Mol Med Rep.
2017;16(6):8237-44.
42. Choi YJ, Choi YK, Lee KM, Cho SG, Kang SY, et al.
SH003 induces apoptosis of DU145 prostate cancer cells by inhibiting ERK-involved pathway. BMC Complement Altern Med. 2016;16(1):507.
43. Jung TK, Kim MJ, Lim KR, Yoon KS. Moisturizing and Anti-oxidation Effect of Astragalus membranaceus Root Extract. Journal of the society of cosmetic scientists of Korea. 2006;32(3):193-200.
44. Yang F, Yan G, Li Y, Han Z, Zhang L, et al. Astragalus Polysaccharide Attenuated Iron Overload-Induced Dysfunction of Mesenchymal Stem Cells via Suppressing Mitochondrial ROS. Cell Physiol Biochem.
2016;39(4):1369-79.
45. Xu C, Tang F, Lu M, Yang J, Han R, et al.
Astragaloside IV improves the isoproterenol-induced vascular dysfunction via attenuating eNOS uncoupling-mediated oxidative stress and inhibiting ROS-NF-κB pathways. Int Immunopharmacol.
2016;33:119-27.
46. Li J, Han L, Ma YF, Huang YF. Inhibiting effects of three components of Astragalus membranaceus on oxidative stress in Chang Liver cells. Zhongguo Zhong Yao Za Zhi. 2015;40(2):318-23.
47. Cao Y, Ruan Y, Shen T, Huang X, Li M, et al.
Astragalus polysaccharide suppresses doxorubicin- induced cardiotoxicity by regulating the PI3k/Akt and p38MAPK pathways. Oxid Med Cell Longev.
2014;2014:674219.
48. Min SH, Han HS, Lee YJ. Study on the Anti-oxidative Effects of Adenophorae Radix, Codonopsis lanceolatae Radix and Glehniae Radix Cum Rhizoma on Liver Cells Isolated from Oxidatively Stressed Rat. The Korea journal of herbology. 2009;24(3):109-19.