NaF-induced Autophagy on SCC25 Human Tongue Squamous Cell Carcinoma Cells
Jin-Mo Kang1, Bo-Young Lee1, In-Ryoung Kim1, Yong-Ho Kim1, Su-Bin Yu1, Hae-Ryoun Park2, and Bong-Soo Park1*
1Department of Oral Anatomy, School of Dentistry, Pusan National University
2Department of Oral Pathology, School of Dentistry, Pusan National University
(received October 15, 2014; revised November 10, 2014; accepted November 24, 2014)
*Correspondence to: Bong-Soo Park, Department of Oral Anatomy, School of Dentistry, Pusan National University, Yangsan, 626-870, Korea.
Tel. : 82-51-510-8242, Fax. : 82-51-510-8241, E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creati- vecommons.org/licenses/by-nc/3.0) which permits unrestricted non- commercial use, distribution, and reproduction in any medium, pro- vided the original work is properly cited.
Fluoride has been accepted as an important material for oral health and is widely used to prevent dental caries in dentistry. However, its safety is still questioned by some.
Autophagy has been implicated in cancer cell survival and death, and may play an important role in oral cancer. This study was undertaken to examine whether sodium fluoride (NaF) modulates autophagy in SCC25 human tongue squamous cell carcinoma cells. NaF demonstrated anticancer activity via autophagic and apoptotic cell death. Autophagic vacuoles were detectable using observed to form by monodansylcadaverine (MDC) and acridine orange (AO).
Analysis of NaF-treated SCC25 cells for the presence of biochemical markers revealed direct effects on the conversion of LC-3II, degradation of p62/SQSTM1, cleavage formation of ATG5 and Beclin-1, and caspase activation. NaF-induced cell death was suppressed by the autophagy inhibitor 3-methyladenine (3-MA). NaF-induced autophagy was confirmed as a pro-death signal in SCC25 cells. These results implicate NaF as a novel anticancer compound for oral cancer therapy.
Key words: NaF, autophagy, apoptosis, human squamous cell carcinoma
Introduction
Fluoride is extensively distributed in water, soils, and the atmosphere, causing constant exposure of human beings and animals to fluoride compounds [1]. The principal sources of ingested fluoride are drinking water or beverages produced with fluoride-containing dental products, and food [1,2].
Fluoride is widely used as a remedy to dental caries, although controversy remains about the safety. While long-term fluoride ingestion for the prevention of dental caries may lead to side effects asserious acute toxicity [3,4]. Nevertheless, fluoride has been considered as an important material for oral health care.
Autophagy is an evolutionary phenomenon, long-lived or damaged proteins and cellular organelles by internal and external stimuli are digested in lysosomes and it is reuse as energy or lead to death within cell [5,6]. However, autophagy also promotes cancer cell survival under conditions of stress, functioning as a defense mechanism in response to various anticancer drugs [7,8]. Therefore, the induction of autophagic cell death by anti-cancer reagents has been recognized to play an important role in cancer therapy [9-11].
Oral squamous cell carcinoma (OSCC) is the most common
193
and persistent malignant tumor of the oral cavity, with nearly 500,000 patients suffering from this disease each year world-wide [12,13]. Three major treatment of oral cancer including surgery, radiation and chemotherapy are currently applied, but it is difficult to cure because successful treatment is not always achieved in patients [14]. Although the OSCC patients are treated with classical treatment modalities, noticeably high mortality rates are still observed [15-17].
Therefore, new therapeutic approaches are being investigated, with the use of natural agents being one of the most promising anti-cancer treatment options.
Treatment with sodium fluoride (NaF) has also been examined in the course of OSCC research, and was shown to cause the death of OSCC cells via apoptosis. However, no reports have yet examined the effects of NaF on autophagic cell death in the SCC25 human tongue squamous carcinoma cells. In the present study, we examined whether NaF can induces autophagic and apoptotic cell death in SCC25 cells, and established the molecular mechanism of association between autophagy and apoptosis.
Materials and Methods
Reagents
Acridine orange (AO), monodansylcadaverine (MDC), and 3-[4,5-dimethylthiazol-2-yl]2,5-diphenyl tetrazolium bromide (MTT) were purchased from Sigma (St. Louis, MO, USA).
3-methyladenine (3-MA) was obtained from Calbiochem (La Jolla, CA, USA). Antibodies against Beclin-1, caspase-3, caspase-8, and PARP were purchased from Cell Signaling Technology (Beverly, MA, USA). Antibodies against microtubule-associated protein 1 light chain 3 (LC3) (Sigma, MO, USA) were also used. The autophagy protein 5 (ATG5), caspase-9, GAPDH, and p62/SQSTM1 were purchased from Santa Cruz Biotechnology (Santa Cruz, CA, USA). The mouse anti-rabbit IgG antibody, and rabbit anti-mouse IgG secondary antibodies were purchased from Enzo Life Sciences (Farmingdale, NY, USA). All other chemicals and reagents were purchased from Sigma, unless otherwise specified.
Cell culture
The SCC25 human oral squamous cell carcinoma cell line was purchased from ATCC (Rockville, MD, USA) and maintained as monolayers in Dulbecco's Modified Eagle
Medium: Nutrient Mixture F-12 (DMEM⁄F-12) with supplemented with 10% FBS and 100 µg/ml streptomycin at 37˚C in a humidified atmosphere containing 5% CO2.
Treatment of NaF
The stock solutions of NaF (100 mM) were dissolved in distilled water, and were prepared as needed. NaF stock solution was diluted to the working concentration with media where needed. Cells were cultured about 80% confluence in wells or plates, and then treated to various concentrations of NaF (0-2 mM) for 24 h. Cells grown in medium without NaF served as the control and Earle’s Balanced Salt Solution (EBSS) used as an autophagy-positive control.
MTT assay
SCC25 cells (1 × 104) were seeded and incubated for 24 h in tissue culture-coated 96-well plate. Then, cells were treated with various concentrations of NaF (0-2 mM) for 24-72 h. After treatment, thiazolyl blue tetrazolium bromide (MTT) solution (500 μg/ml) was added to the plates, and they were incubated at 37°C for 4 h. The medium was aspirated and 100 μl DMSO was added for dissolving the formazan crystals. Cell viability was measured by an ELISA reader (Tecan, Männedorf, Switzerland) at the excitatory emission wavelength of 570 nm.
Flow cytometer analysis
After drug treatment, cells (1 × 106) were harvested by trypsinization. For fix the cells, ice cold 95% ethanol and 0.5% Tween 20 were added until a final ethanol concentration of 70%. The fixed cells were pelleted and then washed PBS in 1% BSA. Cells were resuspended in 1 ml PBS containing 20 μg/ml RNase A, incubated on ice for 30 min, and finally added 10 μl propidium iodide (PI) stock solution (1 mg/ml).
After incubation on ice for 2 min, DNA content was measured and analyzed on a CYTOMICS FC500 flow cytometry system (Beckman Coulter, FL, CA, USA) to allow simultaneous estimation of apoptosis. Induction of autophagy in cells, shows acidic vesicular organelles (AVOs), it can be orescentstained red and detected in FL3 red-fluorescence by acridine orange (AO). To find a ratio of NaF induced autophagy, cells were stained with AO (0.1 μg/mL) for 5 min, analyzed using a FACScan flow cytometer. For inhibition of autophagy, cells were pretreated with 1mM 3-MA for 1 h before incubation with NaF for 24 h.
Fluorescence microscopy
5 × 104 cells were grown on coverslips in 24-well plate and treated with NaF. After treatment, cells were stained with monodansylcadaverine (MDC) (0.05 mM) for 30 min and acridine orange (AO) (0.1 μg/mL) for 15 min at 37°C. Acidic vesicular organelles (AVOs) stained with both MDC and AO fluorescent, and cells were observed using a fluorescence microscope (Axioskop, Carl Zeiss, Germany). EBSS was using as a positive control of autophagy.
Western blot analysis
2 x 106 cells were harvested using a cell scraper, washed twice with ice-cold PBS, and lysed ice-cold RIPA lysis buffer (Invitrogen, Carlsbad, CA, USA) and incubated on ice for 1 h. The lysates were then centrifuged at 14,000 rpm for 15 min at 4°C. Protein quantitation was measured using a Bradford protein assay (Bio-Rad, Richmond, CA, USA), and aliquots of the lysates (20 μg protein) were resolved by 10%
SDS/PAGE. The gels were transferred on a to polyvinylidene fluoride (PVDF) membranes (Millipore, Billerica, MA, USA).
The membranes were kept with 5% non fat dry milk blocking buffer, and incubated with primary antibodies and secondary antibodies for 1 h. Immunoblotting of potein was accomplished via the SuperSignal West Femto (Pierce, Rockford, IL, USA) improved chemiluminescence substrate, detected with an Alpha Imager HP (Alpha Innotech, Santa Clara, USA).
Statistic analysis
All results were representative of at least 3 independent experiments and are expressed as the mean ± SEM (standard error of the mean). Statistical analyses were performed by using the SPSS version 14.0 statistical software program (SPSS Inc., IL, USA). A one-way ANOVA for multiple comparison was used for statistical analysis.
Results
NaF induces apoptosis in SCC25
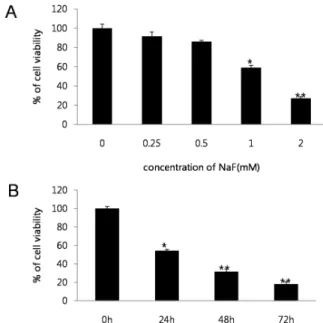
The cytotoxicity of NaF was investigated over a wide concentration range in SCC25 cells. Cells were treated with NaF (0-2 mM) for 24 h, after which viability was assessed using the MTT assay. NaF concentrations from 0 to 2 mM potently induced cell death in SCC25 cells (Fig. 1A).
Treatment with 1 mM NaF resulted in decrease of cell viability
Fig. 1. Cytotoxic effect of NaF on SCC25 cells. NaF treated cell viability was determined by MTT assay. (A) Cells were treated with various concentrations of NaF (0-2 mM) for 24 h.
(B) To investigate cell viability according to the time variation, cells were treated with NaF (1 mM) for 24-72 hours. Data show mean±SEM of three independent experiments. *p < 0.05,
**p < 0.01
Fig. 2. NaF induced apoptotic cell death in SCC25 cells. Cells were treated with NaF (0-2 mM) for 24 h, and the percent of apoptotic cells was determined by FACS.
in a time-dependent manner (Fig. 1B). Thus, the viability of SCC25 cells was decreased in both a dose- and time-dependent manner by NaF treatment. A flow cytometry assay was undertaken to test whether the induction of cell death was mediated via apoptosis. The percentage of sub diploid cells,
indicative of apoptotic cells, was found to increase in a dose-dependent fashion by NaF treatment (Fig. 2).
NaF induced autophagy in SCC25 cells
It was next investigated whether autophagy occurs in the SCC25 cells treated with NaF. When cells were stained with monodansylcadaverine (MDC), a selective marker of autophagic vesicles, the NaF-treated SCC25 cells exhibited staining autophagic vesicles as punctate compared to the control (Fig. 3A). The formation of autophagic vacuoles due to NaF was also confirmed by staining the acidic vesicular organelles (AVOs) with acridine orange (AO). The AVOs, which represent autophagic vacuoles, were clearly formed in the SCC25 cells (red fluorescence). As shown in Fig. 3B, red-stained AVOs were observed following treatment with 1 mM NaF for 24 h. These findings indicate that treatment of NaF is sufficient to promote autophagic response in SCC25 cells, as observed the autophagic vacuoles by MDC and AO staining.
NaF promotes the formation of autophagic and apoptotic molecules in SCC25 cells.
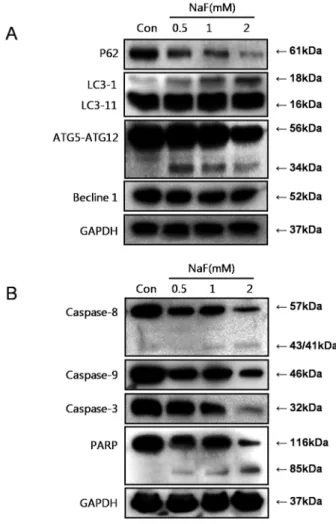
Herein, NaF was also examined for the ability to induce autophagy in SCC25 cells through observation of various autophagy markers, including ATG5, Beclin-1, p62/SQSTM1, and LC3. LC3, the only reliable marker of autophagosomes, was increased in the NaF-treated SCC25 cells in a dose-dependent manner. In contrast, the level of p62/SQSTM1, a protein degraded by autophagy, and Beclin-1 were reduced
Fig. 3. NaF showed induction of autophagy in SCC25 cells.
Cells were seeded in 24-well plates and NaF (1 mM) was treatment 24 h later. NaF induced autophagy was using MDC and acridine orange staining, it was analyzed by fluorescence microscopy. EBSS was used as a positive control of autophagy.
Fig. 4. Autophagy and apoptosis-related proteins were activated by NaF treatement. NaF treatment were performed at various concentrations (0-2 mM) in SCC25 cells. Autophagy-related proteins such as p62/SQSTM1, LC3, ATG5, Beclin-1 and apoptosis–related proteins such as caspase-8, caspase-9, caspase-3, PARP were measured by western blot analysis.
in the NaF-treated SCC25 cells (Fig. 4A). The mechanism of NaF-induced cell death were further investigated by examining the levels of PARP cleavage and the cleavage of procaspase to active caspase-8, caspase-9 and caspase-3, markers of apoptotic activity in SCC25 cells. As shown in Fig. 4B, NaF induced the activation of caspase-8, caspase-9, and caspase-3, also producing 85 kDa processed PARP fragment.
3-MA inhibits NaF-induced apoptosis and autophagy The role of NaF-induced autophagy in SCC25 was further clarified by investigating the effects of treatment with 3-methyladenine (3-MA) as known autophagy inhibitors, on the NaF-treated SCC25 cells. To confirm whether 3-MA can inhibit autophagy, we determined the cell viability of NaF-treated cell lines after pretreatment with 1 mM 3-MA for
Fig. 5. NaF-induced autophagy was usually inhibited by 3-MA in SCC25 cells. (A) Cells were pretreated with 3-MA (1 mM) for 1 h, and then exposed to NaF (1 mM ) for 24 h. Cell viability were analyzed by the MTT assay. (B) Cells were stained with MDC (a) and acridine orange (b), observed with a fluorescence microscope. The ratio of red fluorescence was quantified using FACS flow cytometry (c). Data show mean±SEM of three independent experiments. *p < 0.05.
1 h. Pretreatment with 3-MA slightly increased the viability of the NaF-treated cells (Fig. 5A). 3-MA also inhibited the induction of AVOs by NaF. This was confirmed by AO staining by measurement of the red-to-green fluorescence ratio by FACS (Fig. 5B). Therefore, NaF induced autophagy and 3-MA inhibited this process, resulting in reduced cell death.
In conclusion, these results suggest that NaF induces autophagy and regards death signals leading to apoptosis in SCC25 cells.
Discussion
Human beings and animals are constantly exposed to fluoride compounds, at greater levels than ever due to the increased use of fluorides in industry, medicine, and dentistry [1,18]. Sodium fluoride (NaF), the most widely used material of fluorides, showed no significant cytotoxicity in OSCC cells
at a low dose in the present study. However, NaF has been reported to induce apoptosis via the intrinsic pathway in HL-60 cells at relatively high doses [19-21]. The possible connection between NaF-induced apoptotic cell death and glycolysis [22] or autophagy [23] has also been reported.
Autophagy is widely known as an important process in cell physiology, for both cell survival and death [24]. It starts with the elimination of cytoplasmic organelles in a double-membrane vacuole, an autophagosome, after which they are delivered to a degradative organelle, the vacuole/
lysosome, for breakdown and eventual recycling of the resulting macromolecules. Because numerous recent studies have shown increased autophagic activity to be associated with cell death [10,25], autophagy is now considered as a type of cell death [26]. One of the recent studies reported that autophagy may play a critical role as a protective response in ameloblasts as a result of NaF-induced cell stress. Therefore, the necessity of performed more studies emerged, as the molecular mechanism between NaF-induced apoptosis and autophagy has not been clearly identified.
Previous studies in our laboratory also showed that NaF induced cell death in SCC25 cells via apoptosis. The effect of NaF in the autophagic process of SCC25 has not yet been determined. The present data demonstrated that the NaF-treated SCC25 cells had decreased viability, and underwent cell death via apoptosis and autophagy (Fig. 1, 2).
The autophagic effects of NaF in SCC25 cells were confirmed via AO and MDC staining. NaF induced the formation of cytoplasmic vacuoles and acidic vesicular organelles (AVOs) in the SCC25 cells (Fig. 3A, B).
The NaF-treated SCC25 cells were also analyzed for the presence of biochemical markers of autophagy, including p62/SQSTM1, LC3, ATG5-ATG12 complex and Beclin-1.
NaF treatment directly affected the conversion of LC-3Ⅱ, degradation of p62/SQSTM1 and full-length Beclin-1, and cleavage formation of the ATG5-ATG12 complex (Fig. 4A).
Several previous studies reported that a relationship may exist between LC3 and p62/SQSTM1, which is selectively degraded in cells undergoing autophagy [27-30]. Autophagy mediates a nonspecific bulk degradation pathway responsible for the degradation of the majority of long-lived proteins and some organelles. ATG5-ATG12 conjugation systems are necessary for the formation of the autophagosome [31].
Beclin-1 (Bcl-2-interacting protein-1) is a key protein in autophagy signaling, functioning with Vps34, UVRAG,
AMBRA-1, and Barkor to assemble the PI3KC3 complex during initiation of autophagosome formation [32-34]. Several studies using different cell types and stimuli recently described that caspase-mediated cleavage of Beclin-1 and ATG proteins enhances apoptosis [35-38]. The results obtained herein showed that NaF led to the degradation of caspase-8, caspase-9, and caspase-3, and assumed a decisive role on Beclin-1 (Fig. 4B). The role of autophagy in SCC25 was further clarified by demonstration that the NaF-induced cell death was suppressed by 3-MA, an inhibitor of autophagy. This result strongly implies that NaF-induced autophagy is a pro-death rather than a pro-survival signal (Fig. 5A, B).
In the present study, we reported for the first time that treatment with NaF induced autophagic cell death in SCC25 cells. NaF induced autophagy as a pro-death signal prior to apoptosis in SCC25, hence, treatment of oral cancer with NaF could be a potentially useful therapeutic strategy. Finding the molecular mechanisms behind autophagic cell death induced by NaF is expected to be useful information for the development possibility as a new therapeutic potential against malignant oral cancer.
Acknowledgments
This work was supported by a 2-Year Research Grant of Pusan National University
Conflict of interest
The authors declare that they have no competing interest.
References
1. Ozsvath D. Fluoride and environmental health: a review.
Rev Environ Sci Biotechnol. 2009;8:59-79.
2. Petersen PE, Lennon MA. Effective use of fluorides for the prevention of dental caries in the 21st century: the WHO approach. Community Dent Oral Epidemiol. 2004;32:
319-21.
3. Li Y. Fluoride: safety issues. J Indiana Dent Assoc.
1993;72:22-26.
4. Lee JH, Jung JY, Jeong YJ, Park JH, Yang KH, Choi NK, Kim SH, Kim WJ. Involvement of both mitochondrial- and
death receptor-dependent apoptotic pathways regulated by Bcl-2 family in sodium fluoride-induced apoptosis of the human gingival fibroblasts. Toxicology. 2008;243:340-347.
5. Yorimitsu T, Klionsky DJ. Autophagy: molecular machinery for self-eating. Cell Death Differ. 2005;12 (Suppl 2):
1542-1552.
6. Codogno P. Autophagy in cell survival and death. J Soc Biol. 2005;199:233-241.
7. Zhang JQ, Li YM, Chen XH, Liu T, Chen YT, He WT, Zhang QB, Liu SY. Autophagy is involved in anticancer effects of matrine on SGC-7901 human gastric cancer cells. Oncol Rep. 2011;26:115-124.
8. Michaud M, Martins I, Sukkurwala AQ, Adjemian S, Ma Y, Pellegatti P, Shen S, Kepp O, Scoazec M, Mignot G, Rello-Varona S, Tailler M, Menger L, Vacchelli E, Galluzzi L, Ghiringhelli F, di Virgilio F, Zitvogel L, Kroemer G. Autophagy-dependent anticancer immune responses induced by chemotherapeutic agents in mice.
Science 2011;334:1573-1577.
9. Giannopoulou E, Antonacopoulou A, Matsouka P, Kalofonos HP. Autophagy: Novel Action of Panitumumab in Colon Cancer. Anticancer Res. 2009;29:5077-5082.
10. Gozuacik D, Kimchi A. Autophagy as a cell death and tumor suppressor mechanism. Oncogene. 2004;23:2891-2906.
11. Guo XL, Li D, Hu F, Song JR, Zhang SS, Deng WJ, Sun K, Zhao QD, Xie XQ, Song YJ, Wu MC, Wei LX.
Targeting autophagy potentiates chemotherapy-induced apoptosis and proliferation inhibition in hepatocarcinoma cells. Cancer Lett. 2012;320:171-179.
12. Clayman GL, Ebihara S, Terada M, Mukai K, Goepfert H.
Report of the Tenth International Symposium of the Foundation for Promotion of Cancer Research: Basic and clinical research in head and neck cancer. Jpn J Clin Oncol. 1997;27:361-368.
13. Shen J, Huang CH, Jiang L, Gao F, Wang Z, Zhang YY, Bai JP, Zhou HM, Chen QM. Enhancement of cisplatin induced apoptosis by suberoylanilide hydroxamic acid in human oral squamous cell carcinoma cell lines. Biochem Pharmacol. 2007;73:1901-1909.
14. Suzuki M, Endo M, Shinohara F, Echigo S, Rikiishi H.
Enhancement of cisplatin cytotoxicity by SAHA involves endoplasmic reticulum stress-mediated apoptosis in oral squamous cell carcinoma cells. Cancer Chemother Pharmacol. 2009;64:1115-1122.
15. Bell RB, Kademani D, Homer L, Dierks EJ, Potter BE.
Tongue cancer: Is there a difference in survival compared with other subsites in the oral cavity? J Oral Maxil Surg.
2007;65:229-236.
16. Lo WL, Kao SY, Chi LY, Wong YK, Chang RCS.
Outcomes of oral squamous cell carcinoma in Taiwan after surgical therapy: Factors affecting survival. J Oral Maxil Surg. 2003;61:751-758.
17. Shintani S, Li CN, Mihara M, Klosek SK, Terakado N, Hino S, Hamakawa H. Anti-tumor effect of radiation response by combined treatment with angiogenesis inhibitor, TNP-470, in oral squamous cell carcinoma. Oral Oncol. 2006;42:66-72.
18. Hansen KJ, Clemen LA, Ellefson ME, Johnson HO.
Compound-specific, quantitative characterization of organic fluorochemicals in biological matrices. Environ Sci Technol. 2001;35:766-770.
19. Song JS, Lee HY, Lee E, Hwang HJ, Kim JH. Cytotoxicity and apoptosis induction of sodium fluoride in human promyelocytic leukemia (HL-60) cells. Environ Toxicol Phar. 2002;11:85-91.
20. Anuradha CD, Kanno S, Hirano S. Fluoride induces apoptosis by caspase-3 activation in human leukemia HL-60 cells. Arch Toxicol. 2000;74:226-230.
21. Anuradha CD, Kanno S, Hirano S. Oxidative damage to mitochondria is a preliminary step to caspase-3 activation in fluoride-induced apoptosis in HL-60 cells. Free Radical Bio Med. 2001;31:367-373.
22. Otsuki S, Morshed S, Chowdhury S, Takayama F, Satoh T, Hashimoto K, Sugiyama K, Amano O, Yasui T, Yokote Y.
Possible link between glycolysis and apoptosis induced by sodium fluoride. J Dent Res. 2005;84:919-923.
23. Suzuki M, Bartlett JD. Sirtuin1 and autophagy protect cells from fluoride-induced cell stress. Biochim Biophys Acta.
2014;1842:245-255.
24. Lum JJ, Bauer DE, Kong M, Harris MH, Li C, Lindsten T, Thompson CB. Growth factor regulation of autophagy and cell survival in the absence of apoptosis. Cell 2005;120:
237-248.
25. Tsujimoto Y, Shimizu S. Another way to die: autophagic programmed cell death. Cell Death Differ 2005;12:
1528-1534.
26. Levine B, Yuan J. Autophagy in cell death: an innocent convict? J Clin Invest. 2005;115:2679-2688.
27. Chen LH, Loong CC, Su TL, Lee YJ, Chu PM, Tsai ML, Tsai PH, Tu PH, Chi CW, Lee HC, Chiou SH. Autophagy inhibition enhances apoptosis triggered by BO-1051, an N-mustard derivative, and involves the ATM signaling pathway. Biochem Pharmacol. 2011;81:594-605.
28. Ichimura Y, Komatsu M. Selective degradation of p62 by autophagy. Semin Immunopathol. 2010;32:431-436.
29. Ichimura Y, Kominami E, Tanaka K, Komatsu M.
Selective turnover of p62/A170/SQSTM1 by autophagy.
Autophagy 2008;4:1063-1066.
30. Pankiv S, Clausen TH, Lamark T, Brech A, Bruun JA, Outzen H, Overvatn A, Bjorkoy G, Johansen T.
p62/SQSTM1 binds directly to Atg8/LC3 to facilitate degradation of ubiquitinated protein aggregates by autophagy. J Biol Chem. 2007;282:24131-24145.
31. Jounai N, Takeshita F, Kobiyama K, Sawano A, Miyawaki A, Xin KQ, Ishii KJ, Kawai T, Akira S, Suzuki K, Okuda K. The Atg5 Atg12 conjugate associates with innate antiviral immune responses. Proc Natl Acad Sci USA.
2007;104:14050-14055.
32. Fimia GM, Stoykova A, Romagnoli A, Giunta L, Di Bartolomeo S, Nardacci R, Corazzari M, Fuoco C, Ucar A, Schwartz P, Gruss P, Piacentini M, Chowdhury K, Cecconi F. Ambra1 regulates autophagy and development of the nervous system. Nature 2007;447:1121-1125.
33. Kihara A, Kabeya Y, Ohsumi Y, Yoshimori T. Beclin- phosphatidylinositol 3-kinase complex functions at the trans-Golgi network. Embo Rep. 2001;2:330-335.
34. Sun QM, Fan WL, Zhong Q. Regulation of Beclin 1 in autophagy. Autophagy 2009;5:713-716.
35. Betin VMS, Lane JD. Caspase cleavage of Atg4D stimulates GABARAP-L1 processing and triggers mitochondrial targeting and apoptosis. J Cell Sci. 2009;122:2554-2566.
36. Cho DH, Jo YK, Hwang JJ, Lee YM, Roh SA, Kim JC.
Caspase-mediated cleavage of ATG6/Beclin-1 links apoptosis to autophagy in HeLa cells. Cancer Lett. 2009;
274:95-100.
37. Djavaheri-Mergny M, Maiuri MC, Kroemer G. Cross talk between apoptosis and autophagy by caspase-mediated cleavage of Beclin 1 Oncogene 2010;29:1717-1719.
38. Wirawan E, Vande Walle L, Kersse K, Cornelis S, Claerhout S, Vanoverberghe I, Roelandt R, De Rycke R, Verspurten J, Declercq W, Agostinis P, Vanden Berghe T, Lippens S, Vandenabeele P. Caspase-mediated cleavage of Beclin-1 inactivates Beclin-1-induced autophagy and enhances apoptosis by promoting the release of proapoptotic factors from mitochondria. Cell Death Dis. 2010;1:e18.