Extract from Eucheuma cottonii Induces Apoptotic Cell Death on Human Osteosarcoma Saos-2 Cells via Caspase Cascade Apoptosis Pathway
Chang-Won Kang
1†, Min-Jae Kang
1†, Kyong Rok Kim
1, Nan-Hee Kim
1, Yong Bae Seo
2, Keon-Hee Kang
3, Sang-Ho Kim
3* and Gun-Do Kim
1*
1
Department of Microbiology, College of Natural Sciences, Pukyong National University, Busan 48513, Korea
2
Institute of Fisheries Science, College of Fisheries Science, Pukyong National University, Busan 48513, Korea
3
Yeongsan Skate Co., Ltd., Naju-si 58266, Jeollanamdo, Korea
Received October 12, 2015 /Revised November 10, 2015 /Accepted December 6, 2015
Osteosarcoma (OS) is the most common and malignant bone tumors. Although many types of re- section surgery and experimental agents were developed, median survival and clinical prognosis are poorly investigated. Recently, several researches have reported that Eucheuma cottonii has potent as protective effects of coal dust-induced lung damage via inhibition of malondialdehyde (MDA) and ox- idative stress in bronchoalveolar lavage fluids (BALF). However, anti-cancer effects and specific molec- ular mechanism of extract from Eucheuma cottonii (EE) has not been clearly studied yet. This study evaluated that anti-cancer potential of EE in human osteosarcoma Saos-2 cells. EE indicated cytotox- icity on Saos-2 cells in a dose-dependent manner. Morphological degradation and nucleic con- densation were also observed under the EE treatment. However, it did not significantly affect on non-cancerous kidney HEK-293 cells under the same concentration which is shown cytotoxicity on Saos-2 cells. The phosphorylation of Fas-Associated Death Domain (FADD) and expression of cleaved caspase-8, -7 and -3 were upregulated in a dose-dependent manner. In immunofluorescence staining, expression level of Fas and cleaved PARP were upregulated by EE treatment. Furthermore, treatment of EE induces upregulation of sub G1 phase by flow cytometry analysis. The results demonstrated that EE has a therapeutic potential against osteosarcoma via FADD mediated caspase cascade apopto- sis signal pathway.
Key words : Apoptosis, Eucheuma cottonii, FADD, Saos-2 cells, sub G1 phase
†
Authors contributed equally.
*Corresponding author
*Tel : +82-61-334-0305, Fax : +82-61-336-0585
*E-mail : [email protected] (Sang-Ho Kim)
*Tel : +82-51-629-5619, Fax : +82-51-629-5618
*E-mail : [email protected] (Gun-Do Kim)
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2016 Vol. 26. No. 2. 147~154 DOI : http://dx.doi.org/10.5352/JLS.2016.26.2.147
서 론
뼈는 vascular mineralized tissue에 의해 아주 복잡하고 체 계적으로 유지되고 있으며 근육과 인대, 장기의 지지 및 보호 역할과 미네랄 저장 등의 매우 다양한 기능을 수행하고 있다 [1]. 골육종암은 뼈에 발생하는 암으로 현재 전세계적으로 유 아와 청소년 등 젊은 연령층에서 흔히 나타나며 사망률이 두 번째로 높은 암이다[2, 3]. 현재 가장 많이 쓰이는 치료법으로 는 surgical resection과 methotrexate, doxorubicin, cisplatin 등을 사용한 chemotherapy가 알려져 있고, 이 두 가지 방법의 병행을 통한 치료에 의해 5년 생존율은 70% 정도까지 증가하
였다. 하지만 일부 환자의 경우 chemotherapy의 약효가 제대 로 나타나지 않아 치료가 어려우며, 그로 인해 이를 대체할 새로운 치료 방법에 대한 연구가 계속적으로 요구되는 상황이 다[3-6].
Apoptosis는 흔히 programmed cell death라고 알려져 있으 며 잘못된 세포의 증식과 유전적 결함의 축적을 막아 종양의 형성을 막음으로써 다세포 생물의 생존에서 매우 중요한 역할 을 한다고 알려져 있다[7]. Apoptosis는 크게 extrinsic (death receptor) pathway와 intrinsic (mitochondrial) pathway 두 가 지 경로로 나뉜다. Extrinsic (death receptor) pathway는 세포 막 표면에 있는 FAS ligand와 같은 인자에 의해 활성화된다[8.
FAS ligand는 FAS-Associated Death Domain (FADD)을 활성
화시켜 하위인자인 caspase-8을 활성화 형태인 cleaved cas-
pase-8으로 cleavage시킨다[9, 10]. Cleaved caspase-8은 하위
인자인 caspase-7, caspase-3, PARP [poly (ADP-ribose) poly-
merase] 차례대로 cleavage시키며 활성화시키고 cleaved cas-
pase-3와 cleaved PARP에 의해 DNA fragmentation, chroma-
tin condensation, cell shrinkage, membrane blebbing 등이
진행되어 apoptosis가 유도된다[11]. Intrinsic (mitochondrial)
pathway는 mitochondria membrane의 투과성을 조절하는
Bcl family 중 pro-apoptosis protein인 Bid, Bak, Bad, Bax 등 의 발현을 유도하여 mitochondria membrane의 투과성을 증 대시켜 mitochondrial intermembrane space protein인 cyto- chrome c의 방출을 유도한다[12]. Mitochondria membrane의 외부로 방출된 cytochrome c는 세포질에 존재하는 pro-apop- totic protein들과 복합체를 형성하여 하위인자인 caspase들의 활성화를 유도하여 최종적으로 apoptosis를 유도한다[13].
최근 여러 연구를 통해 해조류는 항암, 항염증, 항미생물, 항궤양성 등 다양한 생리활성을 나타내는 천연물을 제공하는 것으로 보고되고 있다. 이러한 특성때문에 현재 항종양 활성 을 가지는 해조류로부터 유래한 천연물의 분리와 이를 활용한 새로운 항암제의 개발을 위한 연구가 진행되고 있다[14]. 홍조 류의 일종인 Eucheuma cottonii는 아주 깨끗한 물에서 서식하 며 매우 빠르게 성장하여 45일 마다 수확이 가능하다[15]. 현재 까지 알려진 E. cottonii의 기능으로는 석탄먼지와 같은 산화적 스트레스요인에 의한 염증으로부터 보호 작용이 있으나, 항암 효능과 그 메커니즘에 대한 구체적인 연구는 아직 이루어지지 않았다[16-18].
본 연구에서는 E. cottonii의 추출물(이하 EE)에 의해 골육종 암 Saos-2 세포에서 나타나는 항암효과가 apoptosis와 관련된 다양한 단백질들의 어떠한 상호작용을 통하여 일어나는지를 WST-1
®cytotoxicity assay, DAPI staining, Western blot anal- ysis, immunofluorescence staining, flow cytometry를 통해 분석하였다. 실험 결과, EE는 골육종암 Saos-2 세포에서 Fas Associated Death Domain (FADD)에 의한 caspase cascade signal pathway의 활성을 통해 apoptosis를 유도함을 확인하 였다.
재료 및 방법
실험재료
본 실험에 사용한 원료인 E. cottonii는 식품용 카라기난 제 조용으로 인도네시아로부터 수입되어 사용하고 있는 원료를 사용하였다. E. cottonii 원료는 흐르는 물에 침지시켜 24시간 동안 계속적으로 물을 흘려주면서 탈염 처리하였고, 탈염이 끝난 원료는 48시간 진공 건조하여 수분을 8%이하로 제거한 후, 분쇄하여(60 mesh 이하) 추출 원료로 사용하였다. E. cotto-
nii 추출액은 알코올 추출, 감압여과 그리고 감압농축 과정을거쳐 준비되었고, 94% Ethyl alcohol (SK Chemical Co., Korea) 을 원료대비 15배를 가하여 상온에서 24시간 동안 교반하여 얻었다. 추출액은 다시 정성용 여과지(No.2, Advantec, Toyo Roshi Kaisha, Japan)로 진공펌프(N 840.3 FT.18, KnF, Freiburg, Germany)를 이용하여 감압 여과한 뒤, 농축기 (N-1200A, EYELA, Japan)를 이용하여 농축, 동결 건조시켜 실험에 사용하였다.
세포배양
실험에 사용한 인간 골육종암세포 Saos-2, 인간 위암세포 AGS, 인간 간암세포 SK-Hep1, 인간 뇌교모세포종 U87MG, 인간 정상 신장세포 HEK-293는 American Tissue Culture Collection (ATCC, Rockville, MD, USA)에서 분양 받아 사용 하였다. AGS 세포는 RPMI 1640 (Cellgro Mediatec, Inc., Mannassas, USA) 배지, SK-Hep 1과 U87MG는 Minimum Essential Medium (MEM) (Cellgro Mediatec, Inc.) 배지, Saos-2와 HEK-293 세포는 Dulbecco’s Modified Eagle’s Medium (DMEM) (Cellgro, Mediatech, Inc.) 배지를 사용하여 배양하였다. 각 배지는 10% heat inactivated fetal bovine se- rum (Cellgro Mediatec, Inc.)과 100 μg/ml Penicillin and 100 μg/ml Streptomycin (Hyclone, Logan, UT, USA)을 첨가하여 사용하였으며 37℃, 5% CO
2조건 하에서 배양하였다.
세포 증식 억제 효과와 형태적 변화 분석
E. cottonii 추출물이 Saos-2 세포의 증식에 미치는 영향은
WST-1
Ⓡassay로 확인하였다. Saos-2 세포를 96-well micro- plate의 각 well에 1×10
4cells씩 분주하고 24시간 배양한 후, 추출물을 추출물을 100 μg/ml, 300μg/ml 농도로 각각 처리한 후 24시간 동안 배양하였다. 24시간 이후 EZ-Cytox Cell Viability Assay Solution WST-1
Ⓡ(2-(4-iodophenyl)-3(40n- itrophenyl)-5-(2,4-disulfophenyl)-2H-tetrazolium, monosodi- um salt) (Daeil Lab Service, Jong-ro, Korea)을 각 well 에 10 μl씩 넣고 37℃, 5% CO
2의 조건에서 4시간 동안 반응시킨 후 Microplate reader (Molecular Devices, Silicon Valley, CA, USA)를 이용하여 460 nm의 파장에서 흡광도를 측정하였다.
E. cottonii 추출물의 처리에 따른 세포의 형태적 변화는 추출물
처리 24시간 후 도립 현미경 CKX41 (Olympus, Tokyo, Japan) 을 이용하여 200배, 400배의 배율로 관찰하였다.
Western blot analysis에 의한 단백질 발현 분석
단백질 발현량의 변화를 분석하기 위해서 Saos-2 세포에
E. cottonii 추출물을 주어진 농도로 24시간 동안 처리한 후 세포를 회수하였다. 회수된 세포는 ice-cold lysis buffer [50 mM
Tris-Cl (pH 7.5), 150 mM NaCl, 1 mM DTT, 0.5% NP-40,
1% Triton x-100, 1% Deoxycholate, 0.1% SDS and proteinase
inhibitors (PMSF, EDTA, Aprotinin, Leupeptin, Prostatin A)
(Intron biotechnology, Gyeonggi, Korea)]를 첨가하여 4℃에
서 30분간 반응시킨 후 14,000 rpm으로 20분간 원심분리하여
그 상등액을 회수하였다. 상등액에 포함된 단백질의 농도는
bovine serum albumin (BSA)을 standard로 Protein Quantifi-
cation Kit (CBB solution
Ⓡ) (Dojindo Molecular Technologies,
Rockville, MD, USA)를 이용하여 측정하였다. 각각의 단백질
sample은 Laemmli sample buffer (Bio-Rad Laboratories, Inc.,
Hercules, CA, USA)와 섞어 5분간 끓인 후 식혀서, SDS-poly-
acrylamide gel electrophoresis (SDS-PAGE)로 분리하였다.
분리된 단백질을 함유한 acrylamide gel을 nitrocellulose membrane (PALL Life Sciences, MI, USA)으로 electro-trans- fer 시킨 후 5% skim milk를 함유한 PBST buffer (135 mM Sodium chloride, 2.7 mM Potassium chloride, 4.3 mM Sodium phosphate, 1.4 mM Potassium dihydrogen phos- phate, 0.5% Tween-20)으로 상온에서 2시간 정도 반응시켜 비 특이적인 단백질들에 대한 blocking을 실시하였다. 그 다음 PBST로 15분 정도 세척한 후 1차 항체[anti-FAS, anti-FADD, anti-p-FADD (Ser 194), anti-cleaved caspase-8, anti-cleaved caspase-7, anti-cleaved caspase-3, anti-PARP, anti-cleaved PARP, anti-GAPDH (glyceraldehyde-3-phosphate dehydro- genase)] (Cell Signaling Technology Inc., Beverly, MA, USA) 를 처리하여 4℃에서 overnight 시켰다. 1차 항체 반응이 끝난 후 PBST로 세척하고 처리된 1차 항체에 대해 적절한 2차 항체 (horseradish peroxidase-coupled anti-rabbit lgG or anti- mouse lgG) (Cell Signaling Technology Inc.)를 PBST buffer 에 넣은 용액을 이용하여 상온에서 1시간 반응시켰다. 반응 후 다시 PBST로 세척하고 enhanced chemiluminescent (ECL
Ⓡ
) detection solution (Pierce, Rockford, lL, USA)을 처리한 다음 X-ray film에 감광시켜 특정 단백질의 발현양상을 비교 분석하였다.
Immunofluorescence staining
Cover-glass bottom dish에 자란 Saos-2 세포를 4,6-Diami- dimo-2-Phenylindole Dihydrochloride Hydrate (DAPI;
Roche, Germany)으로 15분간 37℃에서 반응시켜 핵을 대비 염색하였다. 1x PBS (phosphate-buffered saline) buffer로 한 번 세척한 후 4% formaldehyde (Junsei Chemical Ltd., Tokyo, Japan)를 이용하여 세포를 고정시켰다. PBS로 5분씩 3번 세척 한 후, blocking과정으로 blocking solution [1 dish 당: Distil- led water 199 μl, 10x PBS 25 μl, Normal rabbit serum 12.5 μl, Normal mouse serum 12.5 μl (Santa Cruz Biotechnology Inc., Santa Cruz, CA, USA), Triton X-100 0.75 μl]을 넣고 상온 에서 한 시간 동안 암반응시켰다. Blocking 후 세척 과정 없이 확인하고자 하는 단백질(FAS, cleaved PARP)과 결합하는 1차 antibody (Cell Signaling Technology Inc.)와 β-actin antibody (mouse; Cell Signaling Technology Inc.)를 넣은 antibody di- lution buffer [1 dish 당: Distilled water 0.9 ml, 10x PBS 0.1 ml, 5% BSA 0.05 g (Bioshop, Canada), Triton X-100 3 μl]를 넣어 4℃에서 overnight 시켰다. 2차 antibody 처리 전 세척과 정은 1x PBS로 5분씩 1차, 2차 세척을 한 후 high salt PBS [50 ml 당: Distilled water 45 ml, 10x PBS 5 ml, NaCl 1.17 g (Sigma-Aldrich, Inc., MO, USA)]로 3차 세척을 하였다. 세척 후 2차 antibody (mouse, rabbit 0.4 μl씩; Cell Signaling Technology Inc.)를 넣은 antibody dilution buffer에서 1시간
30분간 상온에 반응시켰다. 그 후 1차, 3차 세척은 1x PBS로 시행하고 2차 세척은 high salt PBS로 시행하였다. 세척 과정 이 끝난 후 Prolong Gold
®Antifade Reagent (Invitrogen, Eugene, OR, USA)을 cover glass 위에 떨어뜨려 형광이 유지 되게 한 후, slide glass 위에 올려 형광 현미경(Carl Zeiss LSM 710 Confocal Laser Scanning Microscope Carl Zeiss, Jena, Germany)으로 1,000배의 배율에서 관찰하였다.
Flow cytometry 분석
Trypsin-EDTA 처리를 통해 세포를 회수 한 후 70% ethanol 을 처리하여 4℃에서 overnight 시켰다. 이 후 세포를 PBS buf- fer로 풀어 준 후 0.2 mg/ml의 RNase A가 첨가된 PBS buffer 를 처리하여 37℃에서 1시간 동안 반응시켰다. 반응 후 40 μg/
ml의 propidium iodide를 첨가한 후 실온에서 30분 동안 암반 응 시켰다. Sub-G1 phase의 비율은 flow cytometer BD FACS Calibur (Becton-Dickinson, Mountain View, CA, USA)를 이 용하여 분석하였다.
통계분석
실험의 측정 결과는 SPSS ver. 12.3 통계프로그램을 이용하 여 평균(mean) ± 표준편차(SD)로 나타내었으며, 각 실험군 사 이의 통계학적 유의성 검정은 분산분석(Analysis of Variance, ANOVA) 후student’s test를 통해 P value가 0.05 미만(*p<
0.005)인 경우 유의성이 있는 것으로 평가하였다.
결과 및 고찰
E. cottonii 추출물의 정상 및 암세포 증식에 대한 효과
E. cottonii 추출물(EE)이 정상 및 암세포의 증식에 미치는영향을 알아보기 위해 다양한 농도의 EE를 정상 세포인 HEK-293 신장세포주와 다양한 암 세포주(위암세포 AGS, 간 암세포 SK-Hep-1, 교모세포종 U87MG, 골육종암 세포 Saos-2) 에 각각 처리한 후 WST-1
®assay를 통해 각 세포들의 세포생 존율을 분석하였다. EE의 처리농도가 100 μg/ml, 200 μg/ml, 300 μg/ml로 증가함에 따라, Saos-2 세포의 생존율이 각각 78%, 57%, 44%로 감소하는 것을 확인할 수 있었다. 반면 AGS, SK-Hep1, U87MG 세포주들의 생존율 감소는 관찰되지 않았 다. 또한 HEK-293 세포의 생존율은 Saos-2 세포와 같은 처리 농도에서 그 생존율이 크게 감소하지 않음을 관찰할 수 있었 다(Fig. 1). 이를 통하여 EE는 골육종암 Saos-2 세포에 상대적 으로 특이하게 세포생존율을 억제하는 효능을 나타낸다는 것 을 확인하였다.
E. cottonii 추출물 처리에 의한 세포의 형태 변화 및 nucleic condensation 관찰
EE의 처리에 의한 Saos-2 세포의 증식 억제 효과가 Saos-2
Fig. 1. The cytotoxicity of extracts from E.
cottonii on cell proliferation. Each of Saos-2, AGS, SK-Hep1, U87MG and HEK-293 cells were treated with in- dicated concentration of the extracts from E. cottonii for 24 hr. Each bar represents the mean ± SD (n=3).
Asterisks are significant differences from control by ANOVA (*p<0.005).
A
B
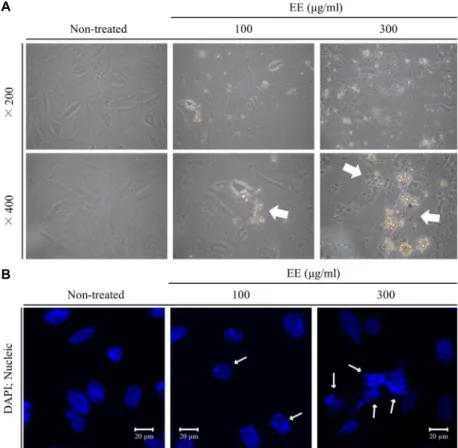
Fig. 2. The extracts from E. cottonii induced morphological changes in Saos-2 cells. (A) Saos-2 cells were treated with the extracts in indicated concen- trations for 24 hr, and imaged cell morphology under a phase contrast inverted microscope at ×200 and
×400 magnifications respectively. (B) Fragmented nucleus in Saos-2 cells were observed followed by DAPI staining. Each arrows are indicated the apoptosome in Saos-2 cells.
세포의 외형 변화와 어떠한 관련성이 있는지 알아보기 위해 도립현미경을 이용하여 세포의 모양 변화를 관찰하였다. EE를 처리하지 않은 대조군에서는 세포의 외형 붕괴가 나타나지 않았고 세포의 외부와 세포막이 구분이 될 만큼 깔끔한 형태 를 유지하는 반면, EE의 처리농도가 100 μg/ml, 300 μg/ml로 각각 증가한 실험군에서는 세포 외형의 붕괴와 세포질에서 apoptosome이 증가함을 확인할 수 있었다(Fig. 2). 세포에서 apopotsis가 유발되면 세포 외형의 붕괴, apoptosome, nucleic fragmentation 등의 전형적인 특성이 나타난다는[19] 보고를 토대로 DAPI staining을 통한 Saos-2 세포 내에서의 핵 형태 변화를 형광현미경으로 관찰하였다. Saso-2 세포에 EE를 각각
0 μg/ml, 100 μg/ml, 300 μg/ml의 농도로 24시간 처리한 후 DAPI solution으로 staining하여 세포를 관찰한 결과, apopto- sis의 전형적 특성인 nucleic fragmentation이 처리 농도가 증 가함에 따라 더 확연히 나타난다는 것을 확인 할 수 있었다 (Fig. 2) [20, 21]. 이러한 결과를 통하여 EE는 Saos-2 세포에 처리시 세포의 apoptosis를 유도하여 세포의 증식을 억제하는 것으로 예측할 수 있었다.
E. cottonii 추출물의 처리에 따른 apoptosis 관련 단백질 의 발현량 변화
Fig. 2의 결과와 같이 EE는 Saos-2 세포에 대하여 농도 의존
A
B
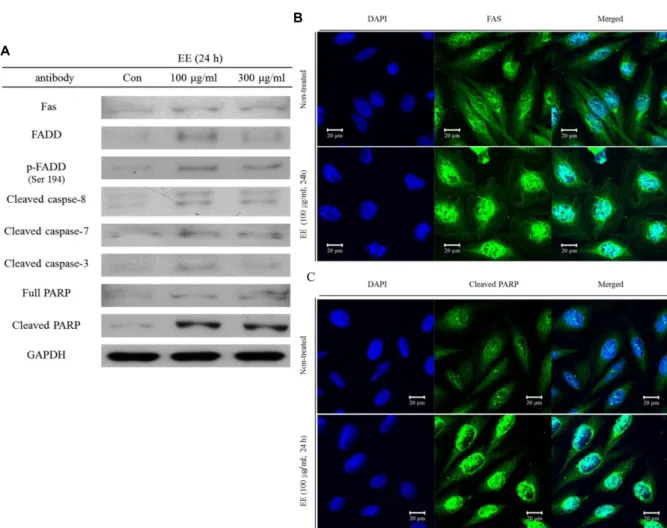
Fig. 3. The extracts from E. cottonii induced FADD mediated apoptosis pathway. (A) Saos-2 cells were treated with 100 μg/ml and 300 μg/ml of the extracts for 24 hr respectively and isolated proteins were analyzed by Western blot analysis with indicated antibodies. GAPDH was used as internal control. Saos-2 cells were treated with the extracts and examined by immunofluorescence staining to detect the expression levels of (B) FAS and (C) cleaved PARP using a confocal laser scanning microscope.
적으로 세포의 증식을 억제하며, 이러한 EE의 Saos-2 세포에 대한 증식 억제 효과는 apoptosis의 유도에 의해 발생될 수 있다는 것을 확인할 수 있었다. 따라서 Western blot 분석을 통하여 apoptosis와 관련된 단백질들의 발현량 변화를 확인하 였다. Extrinsic (Death receptor) apoptosis는 Tumor-Necrosis Factor (TNF), Fas ligand (CD95L), TNF-Related Apoptosis- Inducing Ligand (TRAIL) 같은 ligand들이 세포의 외막에 존 재하는 death receptor에 결합하여 발생한다고 알려져 있다.
[22-24]. 이들 ligand와 death receptor와의 결합은 Fas-Associ- ated Death Domain (FADD)의 recruitment를 유도하며 하위 인자인 caspase-8를 cleaved caspase-8으로 분절시켜 활성화 를 유도한다[25, 26]. 활성화 형태의 cleaved caspase-8은 순차 적으로 하위인자인 caspase-7, caspase-3을 cleavage시켜 cas- pase cascade signal pathway의 활성화를 유도한다[27].
Caspase는 cysteine-containing aspartate-specific protease family로 세포가 정상적으로 성장 및 생존할 때에는 핵과 세포
질에 영향을 주지 않는 불활성화 형태인 pro-enzyme 형태로
존재하고 있다[28]. 그러나 외부의 death signal이나 endoplas-
mic reticulum stress 등이 발생하게 되면 세포는 활성화된 형
태의 cleaved caspase-3를 유도하여 apoptosis의 전형적인 특
징인 DNA fragmentation, chromatin condensation, cell
shrinkage, membrane blebbing을 일으키게 되고[20], DNA
손상 시 수선에 관여하는 중합효소인 PARP (Poly ADO-
Ribose Polymerase)까지 cleavage시켜 cell cycle arrest를 유도
하는 것으로 알려져 있다[20, 29]. 이러한 연구 보고들을 토대
로 Extrinsic (Death receptor) apoptosis와 관련된 단백질들의
발현량을 분석하였다. Saos-2 세포에 EE를 농도별로 24시간
처리한 다음 주요 단백질들의 변화량을 확인한 결과 FADD의
인산화와 발현량이 증가함을 확인할 수 있었다. 또한 FADD의
하위인자인 cleaved caspase-8, -7, -3 그리고 최종적으로
cleaved PARP의 발현량까지 EE의 처리 농도가 증가함에 따
라 의존적으로 증가함을 확인할 수 있었다(Fig. 3A). 이러한
Fig. 4. The extracts from E. cottonii increased sub-G1 phase in Saos-2 cells. Saos-2 cells were treated with the extracts in indicated concentrations for 24 hr, and sub-G1 DNA contents were measured by FACS analysis.
Fig. 5. Schematic diagram of FADD mediated apoptosis pathway induced by the extracts from E. cottonii in Saos-2 cells.
결과는 immunofluorescence staining을 통해 caspase cascade signal pathway의 주요 인자인 FAS와 cleaved PARP의 세포 내 발현량이 EE를 처리함에 따라 증가되어진다는 것을 현미 경하에서 추가적으로 확인할 수 있었다(Fig. 3B, 3C). Fig. 3의 결과를 통하여 EE는 Saos-2 세포에서 extrinsic (death re- ceptor) pathway 중 하나인 FADD에 의한 caspase cascade signal pathway의 활성화를 통해 apoptosis를 유도한다는 것 을 확인하였다.
E. cottonii 추출물에 의한 세포주기의 변화
Western blot 분석과 immunofluorescence staining을 이용 하여 EE가 Saos-2 세포에서 FADD에 의한 caspase cascade signal pathway를 통해 apoptosis를 유도함을 확인하였다
(Fig. 3). Apoptosis가 유발되는 동안 세포는 cytoskeleton col-
lapse, chromatin condensation, DNA fragmentation과 같은
다양한 세포의 생명유지에 치명적인 현상이 나타나게 된다
[19]. 또한 세포 내에서의 정상적인 DNA 수선 기능이 작동되
지 않아 세포주기가 진행되지 못하고 멈추게 되어 sub G1의
비율이 높게 나타나게 되는 것으로 알려져 있다[30, 31]. 이러
한 보고들을 토대로 EE를 Saos-2 세포에 처리하게 되면 apop-
tosis가 유도되고 이와 동시에 DNA 수선 기능에 문제가 생길
경우 sub G1 비율은 증가 할 것으로 예측할 수 있다. 이러한
세포주기의 변화를 확인하기 위해 EE를 Saos-2 세포에 각각
100 μg/ml, 300 μg/ml로 24시간 처리하고 DNA에 결합하는
형광물질인 Propidium Iodide (PI)로 DNA를 staining한 후
flow cytometry로 세포주기의 변화를 분석하였다. 앞서 예측
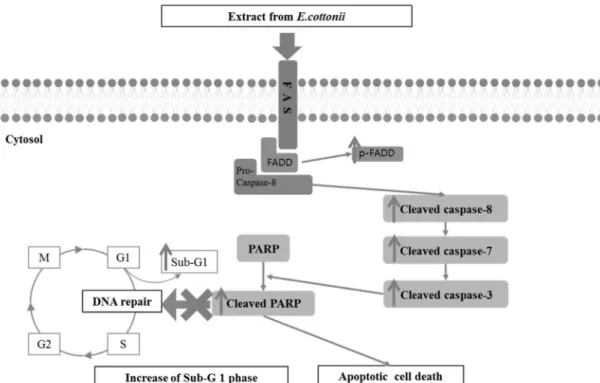
한바와 같이, EE의 처리 농도가 증가함에 따라 Saos-2 세포의 sub-G1 phase가 각각 8.97%, 18.97%로 증가함이 확인되었다 (Fig. 4). 이 결과는 EE의 처리가 Saos-2 세포에서 apoptosis를 유도함과 동시에 세포주기 sub-G1의 비율을 증가시킨다는 것 을 증명하고 있다. 실험의 모든 결과들을 종합하면, EE는 Saos-2 세포에서 FADD에 의한 caspase cascade signal path- way를 활성화시키고, 최종적으로 DNA 수선 단백질인 PARP 를 cleaved PARP로 cleavage시켜 apoptosis를 유도한다고 할 수 있으며 Fig. 5를 통해 그 작용기작을 설명할 수 있다.
과학기술의 발달과 더불어 과거에 비해 골육종 치료에 보다 더 발전된 surgical resection과 다양한 chemotherapy가 이용 되고 있지만, 수술 후 남게 되는 후유증이나 약물 반응성의 제한적인 점 등 여러 가지 문제점으로 인해 천연물유래 항암 치료제의 개발에 대한 관심은 꾸준히 증대되고 있는 상황이다 [32, 33]. 따라서 본 연구에서는 EE를 골육종암의 새로운 천연 물유래 치료제로서 활용할 수 있는지 그 활용 가능성을 알아 보고자 항암 활성 및 작용기전을 확인하였다. EE는 Saos-2 세 포에서 FADD 활성에 의한 caspase cascade signal pathway의 활성을 유도하고 PARP의 cleavage를 증가시켜 apoptosis를 유도하는 것으로 확인되었으며, 이러한 분자적 항암 작용기전 은 골육종암 치료에 대한 새로운 천연물유래 항암제 후보 물 질을 발굴하는 과정의 기초 자료로 사용될 것으로 사료된다.
또한 이러한 기초 기전연구를 바탕으로 실질적인 골육종암 치료제로서의 사용 가능성을 확인하기 위해서는 추가적인 생 리 활성 성분의 순수분리 및 in vivo에서의 추가 실험 등이 수행되어야 할 것이다.
감사의 글
본 연구는 산업통상자원부와 한국산업기술진흥원의 지역 특화산업육성사업으로 수행된 연구결과입니다.
References