LPS로 자극된 RAW 264.7 세포에 대한 제주산 복분자 딸기 추출물의 항염증 효과
박태진1†, 양경월2†, 장성찬1, 김승영1*
Anti-inflammatory Effect of Rubus coreanus Miquel Extract from Jeju Island in LPS-stimulated RAW 264.7 cells
Taejin Park1†, Kyongwol Yang2†, Sungchan Chang1, and Seung-Young Kim1*
Received: 5 July 2019 / Revised: 1 August 2019 / Accepted: 2 August 2019
© 2019 The Korean Society for Biotechnology and Bioengineering
Abstract: Rubus coreabus Miquel are found in the southern part of Korea and their fruits are known to have anticancer, antioxidant and anti-inflammatory properties. However, the suppressive effect of Rubus coreanus Miquel stem and leaf extract (RSL) on the expression of lipopolysaccharide (LPS) - induced inflammatory cytokines and mediators has not been yet examined. So, we evaluated the anti-inflammatory effect of Rubus coreabus stem and leaf extract in LPS-stimulated RAW264.7 cell. In this study, RSL inhibited the generation pro- inflammatory cytokines such as of nitric oxide (NO), prosta- glandin E2 (PGE2), cyclooxygenase-2 (COX-2), inducible NO synthase (iNOS) in a concentration dependent manner without no observable toxicity in RAW 264.7 cells. Our results indicate that RSL could be a new source for the ingredients of cosmetics and pharmaceuticals.
Keywords: Rubus coeanus, lipopolysaccharide, iNOS, COX-2
1. INTRODUCTION
현대인은 서구화된 식생활로 인한 비만과 환경오염 및 스트 레스 등으로 인한 염증 관련 질환이 계속 증가하고 있다 [1].
염증 반응은 본래 손상된 조직과 병원균의 침입으로부터 신 체를 방어하기 위해 가장 먼저 일어나는 원시적인 보호 반응 으로 알려져 있다. 하지만 지속적인 염증 반응은 세포나 조 직의 기능장애 및 괴사를 일으키고 당뇨, 종양, 류머티즘 관 절염 등의 염증 질환으로 발전할 수 있다 [2-5]. 염증반응은 침입으로 인한 조직의 손상에 대해 구조나 기능을 회복시킬 수 있는 유익한 체내 방어기전이다. 그러나 반복되는 염증은 조직 손상의 원인이 되었고 이에 따라 염증을 일으키는 물질 을 찾아 제거하고자 하는 많은 노력이 있었다 [6-8]. 염증반 응에 관여하는 주요 세포로 잘 알려진 macrophage (대식세포) 는 여러 자극뿐만 아니라 면역세포들이 분비하는 cytokine 등에 의해 활성화되어, interleukin-1β (IL-1β), interleukin-6 (IL-6) 및 tumor necrosis factor-α (TNF-α) 와 같은 전염증성 cytokine들의 생산과 NO 및 PGE
2와 같은 염증인자들의 생성 을 촉진한다 [9,10]. 염증과정은 다량의 nitric oxide (NO) 및 prostaglandin E
2(PGE
2) 등 염증 인자가 Nitric Oxide Synthase (NOS) 및 cyclooxygenase (COX)에 의해 형성된다. NO는 혈 압조절 및 응고, 면역기능 등의 역할을 하며 NOS에 의해 형 성된다 [11,12].
최근 항암, 항산화, 미백, 면역 활성 등과 같은 생리활성을 갖는 천연물을 찾는 연구가 국내뿐만 아니라 전 세계적으로 활발히 진행되어 기능성 소재를 첨가한 화장품 등이 출시되 고 있다 [13-15]. 복분자 딸기 (Rubus coreanus)는 장미과 낙
1선문대학교 제약생명공학과
1Department of Pharmaceutical Engineering & Biotechnology, Sun- moon University, Asan 31460, Korea
Tel: +82-41-530-2390, Fax: +82-41-530-2939 e-mail: [email protected]
2(주)제주사랑농수산
2Jeju Love Co., Ltd, Jeju 63358, Korea
Research Paper
엽관목에 속하며 중부 이남의 지역과 제주도뿐만 아니라 중 국과 일본에 서식하는 것으로 알려져 있다 [16]. 한방에서는 복분자 딸기의 열매를 복분자라 하며, 다양한 플라보노이드, 페놀 화합물 및 테르페노이드가 포함되어 있다고 알려져 있 으며, 전통적으로 간, 정력 감퇴, 신정보강 및 빈뇨 등을 치료 하는데 사용되었다 [17,18]. 또한, 복분자의 생리활성은 polyphenol에 의한 항산화 활성, 항균활성 및 항염증 작용 등 이 알려져 있다 [19-21]. 그러나 이러한 효능은 열매에 국한 되어 있으며, 복분자 딸기 (잎과 줄기)에 대해서는 아직까지 항염증 효능 및 그 기전에 대해서는 알려진 바가 없다. 따라 서 본 연구에서는 제주산 복분자 딸기의 잎과 줄기를 복합 추출하여 RAW 264.7 cell에서 발생되는 염증 반응에 대한 억 제 효과 및 기능성 소재로서의 가능성을 검토하고자 하였다.
2. MATERIALS AND METHOD
2.1. 복분자 딸기 추출물 제조
본 연구에 사용된 복분자 딸기 (Rubus coreanus Miquel)의 잎 과 줄기는 2018년 제주특별자치도 서귀포시 대정읍에서 2월 에 채집되었으며 제주생물자원(주)으로부터 제공받았다. 복 분자 딸기 (잎과 줄기)는 70% 에탄올을 이용하여 실온에서 24시간 동안 2회에 걸쳐 추출하였으며, 필터로 여과한 후 감 압농축기를 사용하여 농축하였다. 농축된 추출물을 −110
oC 에서 동결 건조 (LABOGENE, KOREA)한 후 Dimethyl sulfoxide (DMSO)에 용해하여 실험에 사용하였다.
2.2. 세포배양
본 연구에서 사용된 RAW 264.7 세포는 Murine macrophage cell line으로 ATCC (American Type Culture Collection)에서 분양받아 10% fetal bovine serum (FBS)와 100 units/mL penicillin-streptomycin 이 함유된 Dulbecco's Modified Eagle Medium (Gibco, USA) 배양액을 사용하여 37
oC incubator (5% CO
2)에서 2~3일 간격으로 계대배양을 실시하였다.
2.3. 복분자 딸기 추출물의 세포 독성 측정
RAW 264.7 cell 을 24 well plate에 3.0 × 10
5cells/mL 만큼 분 주하여 37
oC, 5%, CO
2조건의 incubator에서 18시간 전 배양 한 후 시료와 LPS (1 μg/mL)를 동시 처리하고, 24시간 동안 배양하였다. MTT (Sigma-Aldrich, St.Louis, MO, USA) 를 첨 가하여 37
oC에서 4시간 동안 반응시킨 다음 MTT 용액을 제 거하였다. 여기에 DMSO를 첨가하여 살아있는 세포와의 반 응으로 생긴 침전물을 용해시킨 후 96-well plate에 옮겨 담 아 ELISA reader를 사용하여 570 nm에서 흡광도를 측정하였 다. LDH assay 측정법은 상등액을 채취하여 LDH (Promega, USA) 용액과 반응시킨 후 490 nm에서 흡광도를 측정하였다.
각 시료 군에 대한 평균 흡광도를 측정하였으며, 대조군의 흡광도 측정값과 비교하여 세포독성을 평가하였다.
2.4. 복분자 딸기 추출물의 NO 생성 억제 활성 측정 RAW 264.7 cell을 10% FBS가 첨가된 DMEM 배지를 사용하 여 24 well plate에 3.0 × 10
5cells/mL로 분주하고 37
oC, 5%
CO
2조건의 incubator에서 18시간 전 배양시켰다. 이후 시료 와 LPS (1 μg/mL)를 동시 처리하여 24시간 배양한 후 상층액 100 μL와 Griess 시약 [1% (w/v) sulfanilamide, 0.1% (w/v) naphylethylenediamine in 2.5% (v/v) phosphoric acid] 100 μL 를 96 well plate에서 혼합하여 10분간 실온 암소에서 반응시 킨 후 ELISA reader를 사용하여 540 nm에서 흡광도를 측정 하였다. 생성된 NO의 양은 세포배양액 중에 존재하는 NO
2-의 형태로 측정하였다.
2.5. Prostaglandin E
2(PGE
2) 생성 억제 활성 측정
RAW 264.7 cell(3.0 × 10
5cells/mL)에 시료와 LPS (1 μg/mL) 를 함유한 450 μL의 배지를 동시에 처리하여 24시간 배양하 였다. 24시간 후 배양 배지를 원심분리(12,000 rpm, 3 min) 하 여 얻어진 상등액의 PGE
2함량을 측정하였다. 모든 시료는 정량 전까지 냉동보관(−20
oC)하여 사용하였다. PGE
2는 mouse enzyme-linked immnunosorbent assay(ELISA) kit(R&D Systems Inc., Minneapolis, MN, USA)를 이용하여 정량하였다.
2.6. 전염증성 cytokines (TNF-α, IL-6, IL-1β) 생성 억제 활 성 측정
RAW 264.7cell 은 1.8 × 10
5cells/mL 로 조절한 후 24 well plate에 분주하고, 37
oC, 5% CO
2incubator에서 18시간 동안 전 배양하였다. 이후 다양한 농도의 시료 50 μL와 LPS (1 μg/
mL) 를 함유한 450 μL의 배지를 동시에 처리하여 24시간 배 양하였다. 이후 배양 배지를 원심분리 (12,000 rpm, 3 min) 하 여 얻어진 상층액의 전염증성 cytokine 생성 함량을 측정하였 다. 전염증성 cytokine은 ELISA kit를 이용하여 정량하였다.
2.7. Western blot analysis
RAW 264.7 cell (3.0×10
6cells/mL)을 18시간 배양한 후 시료와 LPS(1 μg/mL)를 동시 처리하여 24시간 배양하였다. 이후 세포 를 PBS를 이용해 2회 세척하고 lysis buffer [1×RIPA (Upstate Cell Signaling Solution, NY, USA), 1 mM phenylmethylsulfonyl fluoride (PMSF), 1 mM Na
3VO
4, 1 mM NaF, 1 μg/mL aprotinin, 1 μg/mL pepstatin, and 1 μg/mL leupeptin]를 이용해 1시간 동 안 lysis 시킨 후 원심분리 (15,000 rpm, 30 min) 하여 단백질 상층액을 분리하였다. 단백질 농도는 bovine serum albumin (BSA)을 기준으로 Bio-Rad protein assay reagent를 사용하여 정량하였다. 정량한 단백질을 10%의 polyacylamid gel에 전 기영동하고 poly-vinylidene difluoride (PVDF) membrane (Milipore, Burlington, MA, USA)에 200 mA, 2시간 동안 전이 시켰다. 단백질이 전이된 membrane을 5% 탈지분유를 포함 한 0.05% Tween20/Tris-buffered saline (0.05% T/TBS)에 넣 고 상온에서 1시간 blocking 시킨 후, 1차 항체와 반응시켰다.
1차 항체 반응은 iNOS antibody (1:5,000, Calbiochem,
Philadelphia, PA, USA), COX-2 antibody (1:1,000, BD Biosciences
Pharmingen, San Jose, CA, USA), β-actin antibody clone AC- 74 (1:10,000, Sigma, St. Louis, MO, USA)를 이용하여 4
oC에 서 하루 밤 동안 반응시켰다. 1차 항체 반응이 끝난 membrane 은 0.05% T/TBS 용액으로 3회 세척 후 peroxidase-conjugated 된 2차 항체 (Jackson ImmunoResearch, West grove, PA, USA)를 1:5,000 또는 1:10,000으로 희석하여 상온에서 1시간 반응한 뒤 0.05% T/TBS 용액으로 3회 세척하였다. 단백질은 ECL kit (Bio-Rad, USA)를 사용하여 imaging densitometer (model GS-700, Bio-rad, USA) 를 통해 측정하였다.
2.8. 통계처리
모든 실험은 3회 반복하여 측정하였고, 그 결과는 평균값±
표준편차로 나타냈으며 통계적 분석은 SPSS 10.0 프로그램 을 이용하여 각 처리구 간의 유의성 (p<0.05, 0.01)을 검증을 위해 분산분석 (analysis of variance, ANOVA)후 tukey test로 다중비교를 실시하였다.
3. RESULTS AND DISCUSSION
3.1. 복분자 딸기 추출물의 세포 독성 측정
RAW 264.7 세포에 대한 복분자 딸기의 잎과 줄기 추출물 (RSL)의 세포독성을 알아보기 위해 MTT와 LDH 분석법을 이용하여 세포 생존율을 조사하였다. 그 결과 RSL 추출물은 모든 농도에서 무처리군과 비교 시 유의적인 차이를 보이지 않 아 세포의 생존율에 영향을 주지 않음을 확인 하였다 (Fig. 1).
따라서 RSL 추출물은 처리한 농도 내에서 세포독성이 없는 것으로 사료되어 추후 실험에서 위의 농도를 사용하여 실험 을 진행하였다.
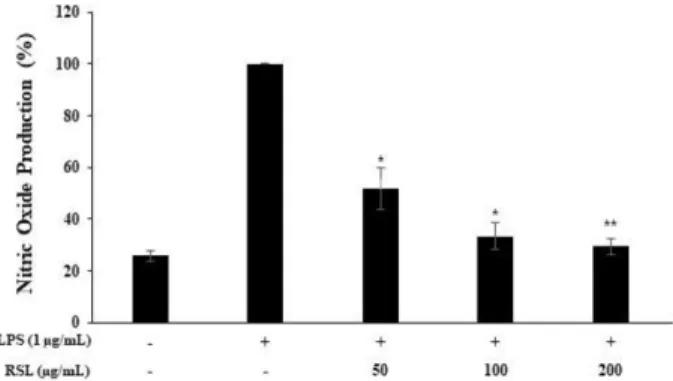
3.2. 복분자 딸기 추출물의 NO 생성 억제 활성
염증을 유발하는 원인이 되는 활성산소 Nitric oxide (NO)의 생성에 RSL 추출물이 어떠한 영향을 미치는지에 대해서 조
사하였다 (Fig. 2). RAW 264.7 세포에 RSL 추출물 (50, 100, 200 μg/mL)과 LPS (1 μg/mL)를 동시 처리하여 NO 생성량을 Griess 시약을 이용하여 세포 배양액에 존재하는 NO
2-의 양 을 측정하였다. 그 결과 LPS 단독 처리군과 비교했을 시 RSL 추출물은 50 μg/mL 농도에서 NO 생성을 48%, 100 μg/
mL 농도에서는 66%, 200 μg/mL에서는 71%로 농도 의존적 으로 NO의 생성량을 감소시키는 것으로 나타났다 (Fig. 1).
따라서 RSL 추출물은 대식세포의 NO의 생성을 감소시킴으 로써 염증 발생에 의한 질환 유발을 효과적으로 제어할 수 있을 것으로 사료된다.
3.3. 복분자 딸기 추출물의 PGE
2생성 억제 활성
COX-2 유래 PGE
2는 면역 관련 세포의 활성을 유도해서 염 증반응을 일으키며 Th2 세포의 활성을 유도해 염증성 cytokine을 과도하게 생성하는 원인으로 알려져 있다 [22].
RSL 추출물을 상기의 NO 실험과 동일한 농도로 처리하여 RAW264.7 세포에서 PGE
2의 생성 억제 활성을 측정하였다.
측정 결과, LPS가 첨가되어 염증이 유발된 군과 비교했을 시 RSL 추출물이 첨가된 각각의 처리 농도 (50, 100, 200 µg/
mL)에서 농도 의존적으로 PGE
2의 생성 억제 효과가 있는 것 을 확인할 수 있었다(Fig. 3). 특히 200 μg/mL 농도에서는 RSL 추출물의 활성이 55%로 PGE
2생성을 억제 하는 것으로 나타났다. 따라서 RSL 추출물은 LPS로 인해 생성량이 증가 된 PGE
2를 감소시킴으로써 항염증 효과를 나타내는 것으로 확인하였다.
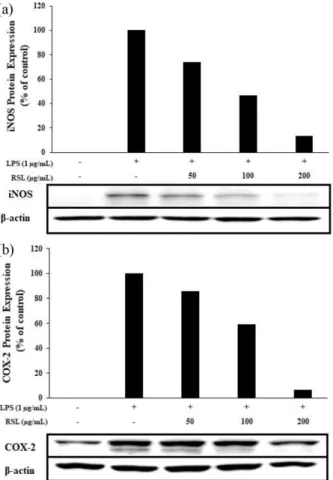
3.4. iNOS 및 COX-2 발현 억제
LPS에 의해 자극을 받은 NF-κB는 핵 내로 이동하여 COX-2 및 iNOS와 같은 여러 염증관련 효소의 전사를 활성화하며, iNOS 및 COX-2에서 유래한 NO와 PGE
2는 급성 및 만성 염 증의 발변 기전에 중추적인 역할을 한다 [23]. 따라서 본 실
Fig. 1. Effects of Rubus coeanus stem and leaf extract on cellviability in RAW 264.7 cells. The production of nitric oxide was assayed in the culture medium of cells stimulated with LPS (1 μg/
mL) for 24 h in the presence of Rubus coreanus extract (50, 100, and 200 μg/mL). Cytotoxicity was determined by LDH assay and MTT assay. Result is expressed as percentages compared to the respective values obtained for the control.
Fig. 2. Inhibitory effects of Rubus coeanus stem and leaf extract on nitric oxide production in RAW 264.7 cells. The production of nitric oxide was assayed in the culture medium of cells stimulated with LPS (1 μg/mL) for 24 h in the presence of Rubus coreanus extract (50, 100, and 200 μg/mL). Cytotoxicity was determined by MTT assay. Result is expressed as percentages compared to the respective values obtained for the control. Data represent the means±SD with three separate experiments. *p<0.05; **p<0.01.
험에서는 RSL 추출물에 의한 염증의 억제가 어떠한 경로에 의한 것인지 확인하기 위해 RAW264.7 세포를 LPS로 자극
한 후 RSL 추출물을 농도별로 처리하여 각 신호 전달물질의 발현 정도를 관찰하였다. 실험 결과, 무첨가군과 비교하여 LPS (1 µg/mL)에 의해 생성이 증가된 iNOS 단백질 발현양이 RSL 추출물을 처리했을 때 농도 의존적으로 크게 감소한 것 을 확인하였다 (Fig. 4(a)). 또한, LPS에 의해 다량 생성된 COX-2의 단백질 발현 양 역시 RSL 추출물에 의해 농도 의 존적으로 감소시키는 것을 확인하였다 (Fig. 4(b)). 이로써 RSL 추출물에 의한 염증 억제가 NF-κB의 신호 전달과정을 통해 이루어지고 있음을 확인하였으며, 이러한 결과는 NO, PGE
2,iNOS, COX-2의 발현을 효과적으로 감소시킴으로써 항염증 효과를 나타내는 것임을 시사했다.
Fig. 3. Inhibitory effects of Sechium edule extract on PGE2
production in RAW 264.7 cells. The production of PGE2 was assayed in the culture medium of cells stimulated with LPS (1 μg/
mL) for 24 h in the presence of Rubus coreanus extract (50, 100, and 200 μg/mL). Result is expressed as percentages compared to the respective values obtained for the control. Data represent the means±SD with three separate experiments. *p<0.05.
Fig. 4. Inhibitory effects of Rubus coreanus extract on the protein level of iNOS and COX-2 expression in LPS-stimulated RAW 264.7 cells. Cells (1.0 × 106 cells/mL) were pre-incubated for 18 h, and then treated with LPS (1
µ
g/mL) and Sample for 24 h. The protein levels of iNOS, COX-2 were analyzed by western blot. β-actin was used as the control.Fig. 5. Inhibitory effects of Rubus coreanus extract on production of pro-inflammatory cytokines in RAW 264.7 cells. The production of (a) TNF-α, (b) IL-6, and (c) IL-1β was assayed in the culture medium of cells stimulated with LPS (1 μg/mL) for 24 h in the presence of Rubus coreanus extract (50, 100, and 200 μg/mL).
Results are expressed as a percentage of the control. Data represent the means±SD with three separate experiments. *p<0.05.
3.5. 전염증성 cytokines(TNF-α, IL-6, IL-1β) 생성 억제 TNF-α, IL-1β 및 IL-6는 염증반응을 조절하는 대표적 pro- inflammatory cytokine으로 알려져 있으며, 이러한 cytokine은 면역세포의 활성, 증식 및 분화를 조절하는 것으로 알려져 있 다 [23]. 따라서 본 실험에서는 LPS로 자극한 RAW 264.7 세 포에 RSL 추출물을 처리한 후 IL-1β, IL-6, TNF-α의 생성량을 측정하여 각각의 발현량을 조사하였다 (Fig. 5). LPS에 의해 RAW264.7 세포의 IL-1β, IL-6, TNF-α의 생성량이 유의적으 로 증가된 것을 확인하였으며, RSL 추출물 (50, 100, 200 μg/
mL) 처리군에서는 LPS에 의해 증가된 IL-1β, IL-6의 생성량 이 농도 의존적으로 억제된 것을 확인하였다 (Fig. 5(a), 5(c)).
특히, RSL 추출물 처리군의 200 μg/mL 농도에서는 IL-1β와 IL-6 분비량이 각각 50% 와 70% 이상으로 억제됨을 확인 하 였다. 반면, TNF-α의 생성능은 LPS처리군과 비교했을 때 억 제능이 미비하게 관찰되는 것으로 보아 RSL 추출물이 TNF-α 생성 기작에는 크게 관여하지 않는 것으로 사료된다 (Fig.
5(a)). 그러나 RSL 추출물은 IL-1β와 IL-6 생성량 감소에는 뛰 어난 효과가 있는 것으로 보아 RSL 추출물은 cytokine의 발현 에 관여함으로써 염증반응을 억제할 수 있음을 시사한다.
4. CONCLUSION
본 연구에서는 RSL 추출물의 항염 효과를 확인하기 위해 Nitric oxide (NO) 저해 활성, Prostaglandin E
2생성 저해 활성 과 전염증성 cytokines 생성 억제, iNOS 및 COX-2 단백질 발 현 억제 효과를 측정하였다. RSL 추출물은 RAW264.7 세포 에 대하여 전 농도에서 세포독성이 관찰되지 않았다. 또한, RSL 추출물은 LPS 자극에 의한 NO 생성을 농도 의존적으로 억제 했다. 게다가 PGE
2생성 억제 활성 측정 결과 LPS 자극 으로 분비된 PGE
2생성을 억제하였으며 전염증성 cytokine 인 IL-1β, IL-6의 분비량을 농도 의존적으로 억제하였다. 대 표적인 염증 관련 신호 전달 경로인 NF-kB 관련 유전자의 발 현을 검토한 결과 RSL 추출물에 의해 iNOS 및 COX-2의 발 현이 유의적으로 억제 되었다. 따라서 RSL 추출물은 천연 항염증 기능성 화장품 소재로의 가능성을 제시하는 결과로 사료된다.
Acknowledgements
본 연구는 산업통상자원부와 한국산업기술진흥원이 지원하 는 광역협력권산업 육성사업으로 수행된 연구결과입니다 (P0004692).
REFERENCES
1. Vlassara, H. (2005) Advanced glycation in health and disease role
of the modern environment. Ann. N.Y. Acad. Sci. 1043: 452-460.
2. Medzhitov, R. (2008) Origin and physiological roles of inflamma- tion. Nature. 454: 428-435.
3. Kaplanski, G., V. Marin, F. Montero-Julian, A. Mantovani, and C.
Farnarier (2003) IL-6: a regulator of the transition from neutrophil to monocyte recruitment during inflammation. Trends Immunol.
24: 25-29.
4. Hofseth, L. J and L. Ying (2006) Identifying and defusing weap- ons of mass inflammation in carcinogenesis. Biochim. Biophys.
Acta. 1765: 74-84.
5. Jeong, H. R., M. S. Sung, Y. H. Kim, H. M. Ham, Y. M. Choi, and J. S. Lee (2012) Anti-Inflammatory Activity of Salvia plebeia R.
Br. Leaf through Heme Oxygenase-1 Induction in LPS-Stimu- lated RAW264.7 Macrophages. J. Korean Soc. Food Sci. Nutr. 41:
888-894.
6. Lee, E. J., E. Y. Whang, K. Whang, I. S. Lee, and S. A. Yang (2009) Anti-allergic effect of Zizania latifolia turcz Extracts.
Korean J. Food Sci. Tecnol. 41: 717-721.
7. An, S. G. (2003) Atopic dermatitis and moisturizer. Update in Der- matology for dermatologists 1: 21-25.
8. Ji, J. D., Y. H. Lee, and G. G. Song (2004) Prostaglandin E2 (PGE2): Roles in immune responses and inflammation. J. Rheu- matic Dis. 11: 307-316.
9. Iwalewa, E. O., L. J. McGaw, V. Naidoo, and J. N. Eloff (2007) Inflammation: the foundation of diseases and disorders. A review of phytomedicines of south african origin used to treat pain and inflammatory conditions. Afr. J. Biotechnol. 6: 2868-2885.
10. Shao, J., Y. Li, Z. Wang, M. Xiao, P. Yin, et al. (2013) A novel naphthalimide derivative, exhibited anti-inflammatory effects via targeted-inhibiting TAK1 following down-regulation of ERK1/2- and p38 MAPK-mediated activation of NF-κB in LPS-stimulated RAW264.7 macrophages. Int. Immunopharmacol. 17: 216-228.
11. Garthwaite, J. (2010) New insight into the functioning of nitric oxide receptive guanylyl cyclase: physiological and pharmacologi- cal implications. Mol. Cell Biochem. 334: 221-232.
12. Crane, B. R., A. S. Arvai, D. K. Ghosh, C. Wu, E. D. Getzoff, et al. (1998) Structure od nitric oxide synthase oxygenase dimer with pterin and substrate. Sci. 279: 2121-2126.
13. Kim, Y. S., S. J. Lee, J. W. Hwang, E. H. Kim, P. J. Park, and J. H.
Jeong (2012) Anti-Inflammatory effects of extracts from Ligus- trum ovalifolium H. Leaves on RAW264.7 macrophages. J.
Korean Soc. Food Sci. Nutr. 41: 1205-1210.
14. Kim, H. J., M. S. Ahn, G. H. Kim, and M. H. Kang (2006) Antiox- idant and antimicrobial activity of Pleurotus eryngii extracts pre- pared from different aerial part. Kor. J. Food Sci Technol. 38: 799- 804.
15. Lee, S. Y., J. C. Yang, and B. A. Kim (2017) A study on the anti- oxidative effects of Zostera marina and its application in cosmat- ics. JKAIS 18: 534-544.
16. Yuk, C. S. (1990) Coloured Medicinal Plants of Korea, pp. 275.
Academy Publish-ing Co., Ltd. Seoul, Korea.
17. Bae, G. H. (2000) The Medicinal Plants of Korea, pp. 231. Kyo- hak Publishing Co., Ltd, Seoul, Korea.
18. Kim, M. S., G. C. Pang, and M. W. Lee (1997) Flavonoids from the leaves of Rubus coreanus. Yakhak Hoeji. 41: 1-6.
19. Kim, T. G., S. Y. Kang, K. K. Jung, J. H. Kang, E. Lee, et al.
(2001) Antiviral activities of extracts isolated from Terminalis chebula Retz., Sanguisorba officinalis L., Rubus coreanus Miq., and Rheum palmatum L. against hepatitis B virus. Phytother. Res.
15: 718-720.
20. Choi, J., K. T. Lee, J. Ha, S. Y. Yun, C. D. Ko, et al. (2003) Antino- ciceptive and anti-inflammatory effects of Niga-ichigoside F1 and 23-hydroxytormentic acid obtained from Rubus coreanus. Biol.
Pharm. Bull. 26: 1436-1441.
21. Cha, H. S., M. S. Park, and K. M. Park (2001) Physiological activ-
ities of Rubus coreanus Miquel. Korean J. Food Sci. Technol. 33:
409-415
22. Jang, S. I., C. S. Jun, K. C. Kwak, M. S. Bae, J. H. Lee, et al.
(2006) Evaluation of korean phytomedicinal plants on inhibition of prostaglandin E2 (PGE2) production and cyclooxygenase-2 (COX- 2) in LPS-stimulated U937 cells. Korean J. Oriental Physiol.
Pathol. 20: 455-459.
23. Baldwin, A. S. Jr. (1996) The NF-kappa B and I kappa B proteins:
new discoveries and insights. Annu. Rev. Immunol. 14: 649-683.