116 서 론
Resveratrol (3,5,4’-trihydoxy-trans-stilbene)은 1963년 호장 (虎杖; Polygonum cuspidatum, Polygonaceae)의 건조된 뿌리에
서 최초로 발견되었으며,1) 포도 덩굴(grapevine)에서는 1976년 Langcake와 Pryce에 의해 보고되었다.2) Resveratrol 은 stilbene 구조체(Fig. 1)이며, 주로 포도나무(Vitis vinifera, Vitaceae)의 과실이나 잎 부위에서 진균, 세균 및 바이러 스의 침입, 자외선, 오존 등의 외부 자극에 대항하는 방
Correspondence to:Sang Kook Lee
College of Pharmacy, Ewha Womans University, 11-1 Daehyeon-dong, Seodaemun-gu, Seoul 120-750, Korea
Tel: +82-2-3277-3023, Fax: +82-2-3277-2851 E-mail: [email protected]
Resveratrol 및 그 유도체의 항염증 작용:
새로운 COX-2 및 iNOS 저해제
이화여자대학교 약학대학 박 은 정․이 상 국
Anti-inflammatory Effects of Resveratrol and Its Analogs:
COX-2 and iNOS as Potential Targets
Eun-Jung Park and Sang Kook Lee
College of Pharmacy, Ewha Womans University, Seoul 120-750, Korea
It has been known that resveratrol, a phytoalexin mainly present in grapes, has antioxidant, anti-inflammatory, and cancer chemopreventive activities. One mechanism of its anti-inflammation and cancer prevention is considered to modulate cyclooxygenase-2 (COX-2) and inducible nitric oxide synthase (iNOS) activities. Since COX-2 and iNOS play important roles in inflammation and carcinogenesis, the potential COX-2 and iNOS inhibitors have been considered as anti-inflammatory or cancer chemopreventive agents. In order to discover novel chemopreventive agents, we synthesized about thirty analogs of resveratrol and evaluated their COX-2 and iNOS inhibitory activities with the production of prostaglandin E2 (PGE2) and nitric oxide (NO) in RAW 264.7 cells, respectively. As a result, several compounds exhibited more potent inhibitory activity than resvera- trol. Among them, 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane which showed inhibitory effects on both PGE2
and NO production was further investigated its mechanism of action on COX-2 and iNOS activities and their expressions. This compound suppressed the lipopolysaccharide (LPS)-induced COX-2 and iNOS protein and mRNA expression. In addition, electrophoretic mobility shift assay (EMSA) study revealed that the suppressions of the COX-2 and iNOS gene expression were related to the inhibition of NF-κB binding activity. Further study exhibited that 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane inhibited IκB-α degradation in LPS-stimulated macrophage cells. These findings suggest that 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane might be a promising candidate for developing cancer chemopreventive agent. (Cancer Prev Res 10, 116-123, 2005)
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ Key Words: Cancer prevention, Cyclooxygenase-2 (COX-2), Inducible nitric oxide synthase (iNOS),
Resveratol analogs
책임저자:이상국, ꂕ 120-750, 서울특별시 서대문구 대현동 11-1 이화여자대학교 약학대학
Tel: 02-3277-3023, Fax: 02-3277-2851 E-mail: [email protected]
접수일:2005년 5월 6일, 게재승인일:2005년 6월 20일
어물질(phytoalexin)로 생합성된다.3)
Resveratrol은 많은 과학자들에 의해 지속적으로 연구 되고 있으며, 대표적인 연구로서는 적포도주가 고지방 식이를 하는 프랑스 및 그리스 지방 사람들의 심혈관계 질환에 의한 사망률을 낮춘다는4,5) ‘French parodox’가 알 려지면서 적포도주의 함유 성분인 resveratrol (0.1∼10 mg/L)4)에 대한 연구도 활발히 이루어졌다. 그 결과, res- veratrol이 항혈전 작용(inhibition of platelet aggregation), 항 산화 작용(free-radical scavenging), endothelin-1 생성 저해 작용, endothelial nitric oxide synthase (eNOS) 유도 작용, high-density lipoprotein (HDL) 생성 증진 작용 및 low- density lipoprotein (LDL) 산화 저해 작용 등을 통해 심혈관 계 보호 효능을 나타내는 것으로 입증되었다.5,6) 또한, 암 예방 및 항암 활성을 나타내는데 암 개시단계(initiation) 에서 resveratrol은 arylhydrocarbon을 epoxydation 및 hydro- xylation화 하여 생체 내 반응성이 높은 물질로 전환하는 phase I 효소 CYP450를 저해한다. 반면, 해독화와 관련된 phase II 효소인 quinone reductase (QR)를 유도함으로써 발 암물질에 의한 DNA 변이를 저해하며, 항산화 활성을 통 해 세포 내에서 생성된 과도한 활성 산소를 소거하여 DNA 손상을 억제한다. 암 촉진단계(promotion)에서는 염 증유발 관련 효소인 cyclooxygenase-2 (COX-2), inducible nitric oxide synthase (iNOS) 및 protein kinase (PK) 등을 저해 하며, 암 진행단계(progression)에서는 HL-60와 같은 미분 화된 암세포의 분화를 유도하고, 다양한 암세포에서의 세포주기 저해 및 세포 사멸을 유도한다.6,7) 실제로, 암 유발 동물 모델(two-stage mouse skin model)에서도 resver- atrol의 암 발생억제 효과가 입증되어, 암예방제로서의 가능성을 제시하였다.8)
본 논문에서는 resveratrol의 위에서 언급한 생리활성 작용 중, 항염증 및 암예방과 관련된 COX-2, iNOS에 대 한 저해 작용에 초점을 맞추어, 합성된 resveratrol 유도체 들의 항염증 및 암예방 효과를 평가하고 새로운 항염증 제 및 암예방제로서의 가능성을 제시하려고 한다.
Cyclooxygenase-2 (COX-2)는 phospholipase A2 (PLA2)에 의 해 세포막 인지질로부터 유리된 arachidonic acid를 prosta-
glandin H2 (PGH2)로 전환하는 과정에 관여하는 효소이 다. 조직 내에서 일정하게 발현되는 COX-1, COX-3와 달 리 COX-2는 성장인자, 염증유발인자, 산화적 스트레스, 외상, 암유전자, 사이토카인 등에 의해 유도되는 효소 (inducible enzyme)이다.9,10) COX-2는 염증 반응에 중요한 역할을 하는 것으로 알려져 있지만, COX-2의 지속적 과 발현은 암을 유발 혹은 악화시키거나, 심혈관계 질병 및 알츠하이머 질병을 유발하는 것으로 알려져 있다.11) 한 편, inducible nitric oxide synthase (iNOS)는 일정하게 발현 되는 neuronal NOS (nNOS) 및 endothelial NOS (eNOS)와 달리, interleukin-1β (IL-1β), tumor necrosis factor-α (TNF- α), lipopolysaccharide (LPS) 등과 같은 염증유발인자 및 암 발생인자에 의해 발현되어, 과량의 nitric oxide (NO)를 생 성한다. 생성된 NO는 프리라디칼의 일종으로 혈관확장 (vasodilation), 비선택적 숙주방어(nonspecific host defense) 등의 작용을 하지만, 지속적 과발현은 만성 염증(chronic inflammation), 천식(asthma), 퇴행성 뇌질환(neurodegenera- tive disease), 다발성 경화증(multiple sclerosis), 관절염(arth- ritis), 암의 진행 및 악화 등을 유발한다.12)
위에서 언급한 바와 같이, resveratrol이 염증 반응 및 암화와 밀접한 관련이 있는 COX-2 및 iNOS의 발현 및 활성화를 저해한다는 기존 연구를 바탕으로 하여, 본 연 구에서는 resveratrol의 구조를 기본으로 hydroxyl 잔기를 methoxy 잔기로 치환하거나 그 위치를 변화시키고, stil- bene 구조 자체에서 benzene을 furane이나 thiophene으로 치환하는 등의 방법으로 합성된 유도체들의 COX-2 및 iNOS 활성 저해능을 평가함으로써 항염증 및 암예방제 로서의 가능성을 제시하고자 한다.13,14)
재료 및 방법 1. 재료
RAW 264.7 마우스 대식세포(TIB-71)는 American Type Culture Collection (Manassas, VA, USA)에서 구입하였다. 세 포 배양에 사용된 Dulbecco’s Modified Eagle’s Medium (DMEM), fetal bovine serum (FBS), antibiotics-antimycotics (peni- cillin G sodium, streptomycin sulfate and amphotericin B), 100 bp DNA ladder는 Invitrogen (Carlsbad, CA, USA)에서 구입 하였다. Prostaglandin E2 표준품 및 PGE2-AchE tracer는 Cayman Chemical (Ann Arbor, MI, USA)에서 구입하였다.
ECL Western blotting detection reagents, rainbow molecular weight marker는 Amersham Pharmacia Biotechnology (Pisca- taway, NJ, USA)에서 구입하였고, anti-goat, anti-mouse, anti-rabbit IgG-HRP antibody, goat polyclonal cyclooxyge- Fig. 1. Structures of stilbene and resveratrol.
nase-2 antibody, rabbit polyclonal iNOS antibody는 Santa Cruz Biotechnology (Santa Cruz, CA, USA)에서 구입하였다.
Mouse monoclonal-actin antibody, TRI reagent, lipopolysac- charide (LPS), dimethyl sulfoxide (DMSO), bicinchoninic acid (BCA)는 Sigma-Aldrich (St. Louis, MO, USA)에서 구입하였 으며, PCR primer로 사용한 oligonucleotide는 Bioneer Cor- poration (Daejon, Korea)에서 합성하였다. Reverse transc- ription (RT) system은 Promega (Madison, WI, USA)에서 구 입하였고, Taq PCR master mix kit은 Qiagen (Valencia, CA, USA)에서 구입하였다.
2. 시료
Resveratrol 및 그 유도체들은 서울대학교 약학대학 김 상희 교수님 연구실에서 제공받았다.
3. 세포배양
RAW 264.7 마우스 대식세포는 FBS (10%)과 antibiotics- antimycotics (100 U/ml penicillin G sodium, 100μg/ml strep- tomycin sulfate and 0.25μg/ml amphotericin B)가 함유된 DMEM 배지에서 37oC, 5% CO2 조건으로 격일마다 계대 배양하였다.
4. Prostaglandin E2 (PGE2) 생성 측정
RAW 264.7 세포를 10% FBS가 함유된 DMEM 배지로 현탁하여 96 well plate의 각 well 당 1×105개씩 넣어 37°C, 5% CO2 배양 조건에서 24시간 동안 배양하였다.
RAW 264.7 세포를 phosphate buffered saline (PBS)으로 1회 세척하고 새로운 DMEM 배지로 교체한 다음 LPS (1μg/
ml)와 시험물질 혹은 시험물질을 녹인 용매(DMSO)를 동 시 처리하여 20시간 동안 배양하였다. 상등액을 희석하 여 PGE2 antibody plate에 가하고 PGE2-AchE trancer를 처리 하여 상온에서 18시간 이상 배양하였다. 배양 후 PGE2
antibody plate를 0.05% Tween in PBS로 1분간 5회씩 세척 한 다음, Ellman’s reagent를 처리하여 상온에서 약 7시간 동안 배양하였다. 405 nm에서 흡광도를 측정한 후, PGE2
표준 용액으로 작성한 검량선에 그 수치를 대입하여 PGE2 생성량을 환산하였다. LPS만을 처리한 군에서의 PGE2 생성량을 기준으로 시험물질 처리군의 PGE2 생성 저해 활성을 non-linear regression analysis를 이용하여 각 시험물질의 IC50 (PGE2 생성을 50% 저해하는 농도)를 결 정하는 방법으로 시험물질 간의 효력을 비교하였다.
5. Nitric oxide (NO) 생성 측정
RAW 264.7 세포를 phenol red가 없는 10% FBS가 함유
된 DMEM에 현탁하여 24 well plate의 각 well 당 5×105 개씩 넣어 37°C, 5% CO2 배양 조건에서 24시간 동안 배 양하였다. RAW 264.7 세포에 LPS (1μg/ml)와 시험물질 혹은 DMSO를 동시 처리하여 20시간 배양한 후 상등액 100μl를 Griess reagent (0.1% naphthylethylenediamine 용액 과 1% sulfanilamide in 5% H3PO4 용액) 180μl과 반응시켜 540 nm에서 흡광도를 측정하였다. 검량선은 sodium nit- rite 용액을 이용하여 작성하였고, 이를 이용하여 흡광도 평균을 nitrite 양으로 환산하였다. LPS만을 처리한 군에 서의 nitrite 양을 기준으로 하여 시험물질 처리군의 NO 생성 저해 활성을 non-linear regression analysis를 이용하여 각 시험물질의 IC50 (NO 생성을 50% 저해하는 농도)를 결정하는 방법으로 시험물질 간의 효력을 비교하였다.
6. 단백질 발현 분석(Western blotting)
RAW 264.7 세포에 시험물질과 LPS (1μg/ml)를 처리하 여 10분 또는 16시간 동안 배양한 후 cell lysate를 만들었 다. 단백질 정량은 BCA 정량법을 이용하였고, 동량(40μg) 의 단백질을 SDS-polyacrylamide gel 전기영동으로 분리하 였다. SDS-polyacrylamide gel에 분리된 단백질을 PVDF membrane으로 옮긴 후 PBS-T (0.1% Tween 20 in PBS)로 5분간 2회 세척하고 5% skim milk in PBS-T에 넣어 상온 에서 1시간 동안 blocking 하였다. 다음, primary antibody 를 1:1,000∼1:2,000 정도가 되도록 PBS에 녹인 3%
skimmed milk로 희석하여 membrane과 함께 상온에서 2∼
3시간 배양하였다. Membrane을 PBS-T로 5분간 3회 세척 한 후 HRP-conjugated secondary antibody를 1:1,500∼1:
2,000으로 희석하여 membrane과 함께 상온에서 2∼3시 간 배양하였다. 이를 PBS-T로 5분간 3회 세척한 후 ECL western blotting detection reagents를 처리하여 생성된 luminescence를 X-ray film에 감광해 확인하였다.
7. mRNA 발현 분석(reverse transcription-poly- merase chain reaction; RT-PCR)
RAW 264.7 세포에 시험물질과 LPS (1μg/ml)를 처리한 후 5시간 동안 배양하여 Chomczynski 및 Sacchi가 제안한 방법으로 TRI reagent를 이용하여 RNA를 분리하였다. 1μg 의 RNA를 avian myeloblastosis virus (AMV)의 역전사효소 (reverse transcriptase)를 이용하여 cDNA를 합성하였으며, 생성된 cDNA에 관심 유전자에 선택적인 primer 및 Taq polymerase를 넣고 PCR을 수행하여 유전자를 증폭하였 다. PCR반응이 끝난 후에는 증폭된 DNA를 2% agarose gel에 전기영동 하였고, SYBR Gold로 염색한 다음 Alpha ImagerTM을 이용하여 DNA band를 확인하였다. PCR 반응
에 사용된 primer는 Table 1에 정리하였다.
8. Electrophoretic mobility shift assay (EMSA) 분석
RAW 264.7 세포에 시험물질을 1시간 동안 전처리한 다음 LPS (1μg/ml)를 가하여 1시간 동안 배양하였다. 세 포에 lysis buffer를 1 ml 씩 가하고 원심분리한 후 생성된 pellet을 NP-40이 함유되지 않은 lysis buffer로 세척해 준 다음, nuclease extract buffer를 가하여 14,000 rpm에서 15분 동안 원심 분리한 다음 상등액을 분주하여 -70°C 냉동 고에 보관하였다. Bradford assay 정량법으로 단백질을 정 량한 다음, 5μg의 단백질을 poly (dI-dC) 2μg 및 5’에 32P
로 labeling된 NF-κB consensus oligonucleotide (100,000 cpm)와 상온에서 25분 동안 반응시켜 DNA-단백질 복합 체를 형성하도록 하였다. 생성된 복합체는 5% polyacryla- mide gel을 이용하여 200 V에서 1시간 동안 전기영동 하 였고, 전개된 gel을 건조시킨 다음 70°C에서 X-ray film에 감광하여 확인하였다.
9. 통계처리
본 연구에서 얻어진 결과에 대해서는 SPSS를 이용하여 각 실험군마다 평균과 표준편차를 계산하고, 군 간의 유 의성을 p<0.01 수준에서 t-test를 실시하여 검증하였다.
Table 1. Sequences of target gene-specific primers used in the PCR
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Target gene Sequences
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Mouse iNOS Sense 5'-ATGTCCGAAGCAAACATCAC-3'
Antisense 5'-TAATGTCCAGGAAGTAGGTG-3'
Mouse COX-2 Sense 5'-GGAGAGACTATCAAGATAGTGATC-3'
Antisense 5'-ATGGTCAGTAGACTTTTACAGCTC-3'
Mouse β-actin Sense 5'-TGTGATGGTGGGAATGGGTCAG-3'
Antisense 5'-TTTGATGTCACGCACGATTTCC-3'
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Fig. 2. Inhibitory effects of resveratrol and its analogs on LPS-induced PGE2production (IC50
values were indicated in brackets). RAW 264.7 cells (5×105 cells/ml) were plated in 96 well plate and incubated for 24 h. The attached cells were washed and incubated in the fresh medium containing LPS (1μg/ml) and serially diluted test compounds. After incubation for 20 h, PGE2
content was determined by enzyme immunometric assay. The percent inhibition was expressed as [1-(PGE2 level of test compounds/PGE2 levels of vehicle treated-control)]×100.
결과 및 고찰
1. Resveratrol 유도체의 PGE2 생성 저해능
RAW 264.7 마우스 대식세포에 LPS와 시험물질(res-
veratrol 및 그 유도체)를 처리하여, LPS에 의하여 유도, 생성되는 COX-2 및 iNOS에 대한 시험물질의 저해 활성 을 평가하였다. 우선, COX-2에 대한 저해 활성은 유도된 COX-2에 의해 생성된 PGH2가 하위 효소에 의해 발현되 는 대표적인 prostaglandin류인 PGE2의 생성량을 측정함 으로써 평가하였다. 그 결과, phenyl ring에 4-methoxy 기 를 가진 styrylheterocycle (compound 1c∼3c)이 phenyl ring 에 3,5-dimethoxy 기를 가진 styrylheterocycle (compound 4c∼
9c)보다 뛰어난 활성을 보였다. 반면, aryl기에 methoxy 기 가 결합된 phenyl ring으로 치환될 경우(compound 10c, 11c) resveratrol에 비해서는 PGE2 생성 저해능이 우수하였 으나, styrylheterocycle을 가진 물질보다는 활성이 저하됨 을 알 수 있었다(Fig. 2). Fig. 3에 제시된 바와 같이 활성 이 우수한 유도체 중 하나인 compound 5c는 농도 의존적 으로 LPS에 의해 유도되는 PGE2 생성을 저해하는 것으 로 나타났다.
2. Resveratrol 유도체의 NO 생성 저해능
Resveratrol 및 그 유도체가 LPS에 의해 유도되는 iNOS 에 어떠한 영향을 미치는지를 확인하기 위하여 iNOS에 의해 유도되는 NO 생성에 대한 시험물질의 저해 활성을
Fig. 4. Inhibitory effects of resveratrol and its analogs on LPS- induced NO production (IC50
values were indicated in brackets). RAW 264.7 cells (5×105 cells/ml) were plated in 24 well plate and incubated for 24 h. The attached cells were washed and incubated in the fresh medium containing LPS (1μg/ml) and serially diluted compound 2n. After incubation for 20 h, NO content was determined by Griess reaction. The percentage inhibition was expressed as [1-(NO level of test compounds/NO levels of vehicle treated control)]×100.
Fig. 3. Dose-dependent inhibition of compound 5c on LPS- induced PGE2 production. Data were represented by mean±SD.
*These values were significantly different compared to LPS- treated control (p<0.01).
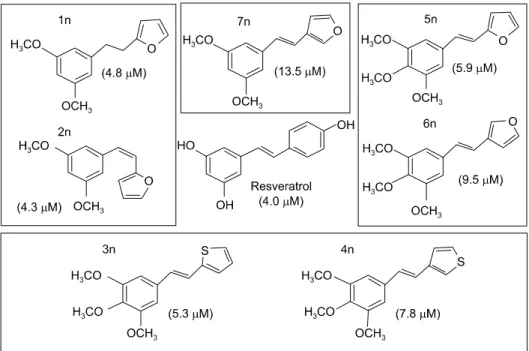
Griess 반응을 이용하여 확인하였다. 그 결과 phenyl ring 에 3,4,5-trimethoxy 기를 가진 styrylheterocycle (compound 3n∼6n)이 3,5-dimethoxy 혹은 4-methoxy 기를 가진 물질 들(data not shown)보다 상대적으로 뛰어난 NO 생성 저해 능을 나타내었다. 3,5-dimethoxy 혹은 4-methoxyphenyl 기 를 가진 물질 중에서는 furanyl 유도체(compound 1n, 2n, 7n)만이 활성을 나타내었다. 특이하게 Z-, E- 이성체는 NO 생성 저해 활성에 거의 영향을 주지 않은 것으로 밝 혀졌다(Fig. 4). NO 생성 저해 효능 물질 중 대표적인 compound 2n의 농도 의존적인 NO 생성 저해 활성을 Fig.
5에 제시하였다.
3. 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane 의 COX-2 및 iNOS 단백질 발현 저해 작용
PGE2 생성 및 NO 생성 저해 활성이 우수하였던 2-[2- (3,5-dimethoxy-phenyl)-vinyl]-furane (Fig. 6; compound 5c in Fig. 5; compound 2n in Fig. 6)에 대하여, COX-2 및 iNOS의 단백질 발현에는 어떠한 영향을 주는지 확인하기 위해 Western blotting을 시행하였다. 실험 결과, 시험물질은 최 고 농도인 21.7μM에서 COX-2 및 iNOS의 발현을 강력
하게 저해하였으며, 농도 의존적인 저해 활성을 보였다 (Fig. 7).
4. 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane 의 COX-2 mRNA 발현 저해 작용
2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane이 COX-2의 mRNA 발현에 미치는 영향을 RT-PCR 법을 통해 확인하였다.
LPS에 의해 RAW 264.7 세포에서의 COX-2 mRNA의 발 현이 증가하였고, 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane 이 COX-2의 mRNA 발현을 농도 의존적으로 저해하는 것으로 나타났다(Fig. 8).
5. 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane 의 유전자 발현 조절 인자에 미치는 영향 평가
2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane가 COX-2 및 iNOS 의 유전자 발현 저해 효능이 있음을 확인함에 따라, 이들 유전자의 발현에 관여하는 transcription factor인 NF-κB가
Fig. 5. Dose-dependent inhibition of compound 2n on LPS- induced NO production.
*These values were significantly different compared to LPS-treated control (p<0.01).
Fig. 6. Chemical structure of 2-[2-(3,5-dimethoxy-phenyl)- vinyl]-furane.
Fig. 8. Inhibitory effect of 2-[2-(3,5-dimethoxy-phenyl)-vinyl]- furane on LPS-induced COX-2 mRNA expression. RAW 264.7 cells (5×105 cells/ml) were plated in 60 mm dish and incubated for 24 h, and then treated with LPS (1μg/ml) and a test compound for 5 h. Cells were lysed, and total RNA (1μg) was used in the RT-PCR. PCR products were separated on a 2%
agarose gel, stained with SYBR Gold, and visualized under UV illumination. β-actin was used as an internal standard.
+ - + + +
LPS (1 g/ml)µ
Compound ( M)µ 0 0 21.7 10.9 5.4 COX 2-
β-actin
Marker
Fig. 7. Inhibitory effect of 2-[2-(3,5-dimethoxy-phenyl)-vinyl]- furane on LPS-induced COX-2 and iNOS protein expressions.
RAW 264.7 cells (5×105 cells/ml) were plated in 60 mm dish and incubated for 24 h, and then treated with LPS (1μg/ml) and a test compound for 16 h. After incubation, cells were lysed, and protein (40μg) was applied on a 8% SDS-polyacrylamide gel.
The level of iNOS or COX-2 protein expression was examined by Western blot analysis.
+ - + + +
LPS (1 g/ml)µ
Compound ( M)µ 0 0 21.7 10.9 5.4 iNOS
COX 2-
β-actin
DNA와 결합하는 데에 이 물질이 어떠한 영향을 주는지 EMSA로 확인하였다. 그 결과, 시험물질은 NF-κB와 DNA의 결합을 농도 의존적으로 저해하는 것을 확인하 였다(Fig. 9). 다음으로, 본 시험물질의 NF-κB에 대한 저 해 활성이, 세포질에서 NF-κB와 공존하는 IκB의 분해를 막아 NF-κB의 핵 내 이동을 저해한 것에서 기인하는지 를 알아보기 위해, IκB단백질의 분해 정도를 확인해 보 았다. IκB는 LPS를 처리하였을 때 10여분 정도 경과하면 최대로 분해 되었고, 따라서 10분 후의 IκB 단백질 분해 정도를 확인한 바, 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane 은 최고 농도인 21.7μM에서 IκB의 분해를 강력하게 저 해하는 것을 확인 할 수 있었다(Fig. 10).
결 론
최근 의료 기술의 발달로 인한 평균 수명의 증가, 산업 화에 따른 환경오염, 생활 방식의 변화 등으로 암 발생률 이 증가하고 있는 추세이다. 따라서 암 발생 이전에 암을 예방하자는 ‘화학적 암예방’에 대한 관심이 커지고 있으 며 특히 천연물로부터 유래한 물질의 암예방 활성에 대 한 연구가 활발히 이루어지고 있다. 만성적인 염증 반응
으로 인하여 암화가 촉진될 수 있다는15) 여러 연구 결과 들에 따라, 염증 유발 인자인 COX-2 및 iNOS의 활성을 저해하는 물질 또한 암예방제로서 역할을 할 수 있을 것 으로 여겨진다. 본 논문에서는 resveratrol의 항염증 및 암 예방 효능을 바탕으로 하여 resveratrol보다 활성이 우수 한 물질을 도출하기 위하여 여러 유도체들을 합성하여 이들의 COX-2 및 iNOS에 대한 저해 활성을 검색하였으 며, 효능물질인 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane의 COX-2 및 iNOS 저해 작용기전을 제시하였다. 이러한 연 구는 보다 우수한 효능을 지닌 항염증제 및 화학적 암예 방 물질의 개발에 기여할 수 있을 것으로 사료된다.
감사의 글
본 연구는 이화여자대학교 약학연구소의 연구비 지원 으로 수행되었으며 이에 감사드립니다.
참 고 문 헌
1) Nonomura S, Kanagawa H, Makimoto A. Chemical cons- tituents of polygonaceous plants. I. Studies on the components of Ko-jo-kon (Polygonum cuspidatum SIEB et ZUCC). Yakugaku Zasshi 83, 983-988, 1963.
2) Langcake P, Pryce RJ. The production of resveratrol by Vitis vinifera and other members of Vitaceae as a response to infec- tion or injury. Physiol Plant Pathol 9, 77-86, 1976.
3) Jeandet P, Douillet-Breuil AC, Bessis R, Debord S, Sbaghi M, Adrian M. Phytoalexins from the vitaceae: biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J Agric Food Chem 50, 2731-2741, 2002.
4) Kopp P. Resveratrol, a phytoestrogen found in red wine. A possible explanation for the conundrum of the ‘French para- dox’? Eur J Endocrinol 138, 619-620, 1998.
5) Stojanović S, Brede O. Elementary reactions of the antioxidant action of trans-stilbene derivatives:resveratrol, pinosylvin and Fig. 9. Inhibitory effect of 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-
furane on DNA binding activity of NF-κB in LPS stimulated RAW 264.7 cells. RAW 264.7 cells (5×105 cells/ml) plated in 100 mm dish and incubated for 24 h, and then pretreated with a test compound for 1 h. After incubation, LPS (1μg/ml) was treated for an additional 1 h. Cells were lysed and nuclear extracts were prepared, and 5μg of nuclear extracts were used in the EMSA.
DNA-protein complexes were separated on 5% polyacrylamide gel. After 1h, the gel was vacuum-dried, and autographed at 70oC for 9∼12 h.
+ - + + +
LPS (1 g/ml)µ
Compound ( M)µ 0 0 40 15 5
NF B DNA binding
-κ -
Fig. 10. Inhibitory effect of 2-[2-(3,5-dimethoxy-phenyl)-vinyl]- furane on LPS-induced IκB-α degradation in cultured RAW 264.7 cells. RAW 264.7 cells were pretreated with various concentration of a test compound for 1h, and then stimulated with LPS (1μg/ml) for 10 min (Western blotting).
+ - + + +
LPS (1 g/ml)µ
Compound ( M)µ 0 0 21.7 10.9 5.4 l Bκ -α
β-actin
4-hydroxystilbene. Phys Chem Chem Phys 4, 757-764, 2002.
6) Savouret JF, Quesne M. Resvertarol and cancer: a review.
Biomed Pharmacother 56, 84-87, 2002.
7) Cao Y, Cao R, Brakenhielm E. Antiangiogenic mechanisms of diet-derived polyphenols. J Nutri Biochem 13, 380-390, 2002.
8) Jang M, Cai L, Udeani GO, Slowing KV, Thomas CF, Beecher CWW, Fong HHS, Farnsworth NR, Kinghorn AD, Mehta RG, Moon RC, Pezzuto JM. Cancer chemopreventive activity of resveratrol, a natural product derived from grapes. Science 275, 218-220, 1997.
9) Vane JR, Bakhle YS, Botting RM. Cyclooxygenase 1 and 2.
Annu Rev Pharmacol Toxicol 38, 97-120, 1998.
10) Hla T, Sishop-Bailey D, Liu CH, Schaefers HJ, Trifan OC.
Cyclooxygenase-1 and 2 isoenzymes. Int J Biochem Cell Biol 31, 551-557, 1999.
11) Hata AN, Breyer RM. Pharmacology and signaling of prosta- glandin receptors: multiple roles in inflammation and immune modulation. Pharmacology & Therapeutics 103, 147-166, 2004.
12) Lala PK, Chakraborty C. Role of nitric oxide in carcinogenesis and tumor progression. Lancet Oncol 2, 149-156, 2001.
13) Lee SK, Park EJ, Lee E, Min HY, Kim EY, Lee T, Kim S.
Styrylheterocycles as a novel class inhibitor of cyclooxygenase- 2-mediated prostaglandin E2 production. Bioorg Med Chem Lett 14, 2105-2108, 2004.
14) Lee SK, Min HY, Huh SK, Kim EY, Lee E, Song S, Kim S. Styrylheterocycles: a novel class of inhibitors on lipopoly- saccharide-Induced nitric oxide production. Bioorg Med Chem Lett 13, 3689-3692, 2003.
15) Li Q, Withoff S, Verma IM. Inflammation-associated cancer:
NF-κB is the lynchpin. Trends Immunol 26, 318-325, 2005.


![Fig. 10. Inhibitory effect of 2-[2-(3,5-dimethoxy-phenyl)-vinyl]- 2-[2-(3,5-dimethoxy-phenyl)-vinyl]-furane on LPS-induced IκB-α degradation in cultured RAW 264.7 cells](https://thumb-ap.123doks.com/thumbv2/123dokinfo/5455469.240263/7.892.479.801.158.254/inhibitory-effect-dimethoxy-phenyl-dimethoxy-induced-degradation-cultured.webp)
