Induction of Apoptosis by Ethanol Extract of Cnidium officinale in Human Leukemia U937 Cells through Activation of AMPK
Jin-Woo Jeong
1, Yung Hyun Choi
1,2and Cheol Park
3*
1Department of Biochemistry, Dongeui University College of Oriental Medicine, Busan 614-052, Korea
2Anti-Aging Research Center & Blue-Bio Industry RIC, Dongeui University, Busan 614-714, Korea
3Department of Molecular Biology, College of Natural Sciences and Human Ecology, Dongeui University, Busan 614-714, Korea Received August 7, 2015 /Revised September 10, 2015 /Accepted September 16, 2015
Cnidium officinale, a traditional herb, has diverse beneficial pharmacological activities, such as anti-in-
flammatory, antioxidant, anticancer, and antiangiogenesis effects. However, the cellular and molecular mechanisms of apoptosis by C. officinale are poorly defined. The present study investigated the proa- poptotic effects of water, ethanol, and methanol extract of C. officinale (WECO, EECO, and MECO, re- spectively) in human leukemia U937 cells. The antiproliferative activity of EECO was higher than that of WECO and MECO. The antiproliferative effect of EECO treatment in U937 cells was associated with the induction of apoptotic cell death, including increased populations of annexin-V positive cells, the formation of apoptotic bodies, DNA fragmentation, and increased numbers of cells with a loss of mi- tochondrial membrane potential (MMP, Δψm). EECO-induced apoptotic cell death was associated with upregulation of death receptor 4 (DR4) and down-regulation of cellular inhibitor of apoptosis pro- tein-1 (cIAP-1), Bcl-2, and total Bid. The EECO treatment also induced the proteolytic activation of cas- pases (-3, -8, and -9), and degradation of caspase-3 substrate proteins, such as poly(ADP-ribose) poly- merase (PARP), β-catenin, and phospholipase C-γ1 (PLCγ1). In addition, the EECO treatment effec- tively activated the adenosine monophosphate-activated protein kinase (AMPK) signaling pathway.
However, compound C, a specific inhibitor of AMPK, significantly reduced EECO-induced apoptosis.
These results indicate that AMPK is a key regulator of apoptosis in response to EECO in human leu- kemia U937 cells.
Key words :
AMPK, apoptosis, Cnidium officinale, U937 cells
*Corresponding author
*Tel : +82-51-890-1530, Fax : +82-505-182-6870
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Journal of Life Science 2015 Vol. 25. No. 11. 1255~1264 DOI : http://dx.doi.org/10.5352/JLS.2015.25.11.1255
서 론
세포의 죽음은 necrosis와 apoptosis의 과정으로 유발되는 데 먼저 necrosis는 에너지 비의존적이며, 세포의 팽창, 이온 농도의 변화 및 물의 유입등과 같은 조절되지 않는 상태로 유발되는 세포의 죽음인 반면 programmed cell death라고 불 리는 생리학적 과정인 apoptosis는 개체의 발생단계나 DNA 손상, 바이러스 감염 등에 의한 유전적 조절 하에서 일어나는 정교한 개체의 생존을 위한 방어기전으로서 손상된 세포들의 제거를 위한 중요한 수단이다[10, 27]. 또한 apoptosis는 세포 의 종류와 손상된 형태에 따라 여러 유전자의 증가 또는 감소 등과 같은 상호작용에 의해 유발되며, 세포막 수포화 현상, 염색질 응축, 핵 단편화 현상, 세포 표면에 phosphatidylserine 발현, apoptotic body 형성, 미토콘드리아 변화 및 caspases
활성화 등과 같은 형태적 또는 생화학적인 변화를 동반한다 [13, 21]. 이러한 apoptosis는 개체의 형태형성, 세포의 전환 및 손상된 조직이나 감염된 세포들의 제거 등과 같은 생물학 적 현상과 건강을 유지하는데 중요한 역할을 하지만, apopto- sis 과정이 실패하게 되면 암을 포함하는 여러 가지 질병의 원인이 되므로 암화 과정의 여러 단계에서 암을 치료하는 중 요한 표적이 되고 있다[18].
AMPK는 catalytic α subunit과 regulatory β 및 γ subunit이
heterotrimeric complex로 구성되어 있는 serine/threonine
kinase이며, 에너지 균형을 조절하는 중요한 조절자 및 에너지
센서로서 지질과 포도당 대사의 조절인자로서 당뇨와 비만에
중요한 조절작용을 할 뿐만 아니라 암세포의 증식을 억제하는
것으로도 알려지고 있다[7]. AMPK는 에너지 소모에 의하여
증가된 세포 내 AMP에 의하여 유발된 AMP/ATP ratio 증가
에 따른 γ subunit의 AMP 결합으로 allosteric activation 촉진
에 의한 활성화와 AMPK upstream kinases인 liver kinase B1
(LKB1), Ca
2+/calmodulin- dependent protein kinase kinase
β (CaMKKβ), TGFβ-activated kinase (TAK-1) 등에 의한 α
subunit의 threonine residue Thr172 인산화에 의해 활성화가
유발된다[20, 24, 32]. AMPK의 활성화는 ROS 생성과 더불어
AKT 인산화 억제를 통한 Forkhead box O3a (FoxO3a) 및
proapoptotic protein인 Bim의 발현 증가 또는 tumor sup- pressor인 p53의 Ser15를 인산화 시킴으로서 미토콘드리아의 기능이상을 유발하여 apoptosis를 일으키는 것으로 보고되고 있다[3, 15, 29].
본 연구에 사용한 천궁(Cnidium officinale)은 미나리과 (Umbelliferae)에 속하는 다년생 초본이며, 뿌리나 지하부를 이용하는 약용자원 식물로서 진정, 진통 및 강장 등에 효능이 있어 예로부터 두통, 동통, 추위로 인한 근육의 마비, 빈혈증 및 부인병 등을 치료하는데 사용되어 왔다. 천궁의 근경에는 senkyunolide A~M, ligustilide, butylidene phthalide, cnidi- lide, neocnidilide, tetramethylpyrazine, sedanolide, β-sitos- terol 등과 같은 다양한 기능성 성분을 함유하고 있으며, 대뇌 활동억제, 혈관운동중추, 호흡중추, 척수반사의 흥분작용, 혈 압강하, 심근수축, 혈관확장, 항균작용, 정형, 진정에 효능이 있는 것으로 알려져 있다[22, 26]. 또한 최근 연구에 따르면 천궁은 항염증, 항산화 및 혈관신생 억제뿐 만 아니라 항암효 능도 있는 것으로 보고되고 있다[5, 12, 14, 16]. 현재까지의 연구에서 볼 수 있듯이 천궁은 다양한 약리작용을 가지는 기 능성 물질이지만 인체 혈구암 세포에서 유발하는 세포독성 효능 및 그에 따른 생화학적 기전에 대해서는 명확히 밝혀져 있지 않다.
따라서 본 연구에서는 천궁의 항암 효과와 그에 따른 생화 학적 기전의 해석을 위하여 인체 혈구암 세포주인 U937 세포 를 대상으로 하여 천궁의 열수 추출물(water extract of C. offi-
cinale, WECO), 에탄올 추출물(ethanol extract of C. officinale,EECO) 및 메탄올 추출물(methanol extract of C. officinale, MECO)이 암세포의 생존율 및 증식에 미치는 영향을 조사하 였고, 이들 중 가장 효능이 좋은 EECO가 apoptosis에 어떠한 영향을 미치는 지를 조사한 결과 유의적인 결과를 얻었기에 이를 보고하는 바이다.
재료 및 방법
실험재료
단백질 분석을 위하여 사용된 TRAIL, DR4, DR5, Fas, FasL Bcl-2, Bcl-X
L, Bax, Bad, Bid, XIAP, cIAP-1, cIAP-2, surviving, caspase-3, caspase-8, caspase-9, PARP, β-catenin, PLCγ1, AMPK, pAMPK, ACC, pACC 및 actin 1차 항체는 Santa Cruz Biotechnology Inc. (Santa Cruz, CA, USA) 및 Cell Signaling Technology (Beverly, MA, USA)에서 구입하였으며, im- munoblotting을 위해 사용된 2차 항체는 Amersham Life Science Corp. (Arlington Heights, IL, USA)에서 구입하였다.
미토콘드리아 활성을 측정하기 위하여 사용된 5,5′, 6,6′- tetrachloro-1,1′,3,3′-tetraethyl-imidacarbocyanine iodide (JC-1)은 Sigma (St. Louis, MO, USA)에서 구입하였으며, DNA flow cytometry를 위하여 사용된 CycleTEST PLUS DNA
REAGENT Kit는 Becton Dickinson (San Jose, CA, USA)에서 구입하여 사용하였다. 또한 caspases의 in vitro 활성 측정을 위한 colorimetric assay kit는 R&D Systems (Minneapolis, MN, USA)에서 구입하였으며, AMPK의 활성을 억제하기 위 하여 사용된 AMPK 억제제인 compound C는 Sigma-Aldrich (St. Luis, MO, USA) 제품을 이용하였다.
시료준비
본 실험에 사용된 천궁은 동의대학교 부속 한방병원에서 제공 받았으며, WECO, EECO 및 MECO를 얻기 위하여 흐르 는 물로 충분히 세척하고 건조시킨 후 잘게 분쇄하였다.
WECO의 경우 건조된 천궁 100 g에 증류수 1 l를 첨가하여 180-200℃로 3시간 가열한 후 상층액만 분리하였고, EECO 및 MECO의 경우 건조된 천궁 100 g에 각각 ethanol 및 methanol 을 첨가하여 60℃, 150 rpm으로 3일간 교반시킨 후 상층액만 분리하였다. 이렇게 준비된 각각의 상층액을 3,000 rpm에서 20분간 원심분리 시켜 찌꺼기를 제거한 다음 Whatman 필터 (No. 2)로 걸러낸 다음 동결 건조 및 감압 농축과정을 통하여 고형성분을 얻어내어 막자사발로 잘게 마쇄하고 밀봉시켜 -7 0℃ 초저온 냉동고에 보관하였다. 실험 시에는 3차 증류수 및 dimethyl sulfoxide (DMSO, Amresco, Solon, OH, USA)를 이 용하여 100 mg/ml의 농도로 만든 다음 이를 적정 농도로 배지 에 희석하여 처리하였다.
세포의 배양
실험에 사용한 인체 혈구암 세포주인 U937 세포는 Amer- ican Type Culture Collection (ATCC, Rockville, MD, USA)에 서 분양 받았으며 90% RPMI-1640 배지(Gibco-BRL, Grand Island, NY, USA)에 10% fetal bovine serum (FBS, Gibco BRL), 2 mM glutamine, 100 U/ml penicillin 및 100 μg/ml streptomycin (Gibco BRL)이 포함된 배지를 사용하여 5%
CO
2, 37℃의 조건하에서 배양하였다. 세포수의 증식에 따른 과밀도 현상을 해소하기 위하여 성장배지의 교환을 매 48시간 마다 실시하여 적정수의 세포를 유지하였다.
Hemocytometer를 이용한 세포 생존율 측정
WECO, EECO 및 MECO 처리에 따른 생존율을 확인하기
위하여 세포 배양용 6 well plate에 U937 세포를 2 × 10
5개/ml
씩 분주한 다음 각각의 시료를 적정농도로 처리하여 배양하였
다. 24시간 경과 후 원심분리하여 상층액을 제거하고 세포를
모은 후 동량의 phosphate-buffered saline (PBS)와 0.5% try-
pan blue solution (Gibco BRL)을 첨가하여 2분 간 염색하고
hemocytometer에 적용하여 염색이 되지 않은 살아있는 세포
의 수를 도립 현미경(inverted microscope, Carl Zeiss, Germa-
ny)을 이용하여 계수하였다.
MTT assay에 의한 세포 증식억제 측정
WECO, EECO 및 MECO 처리에 의한 증식억제의 정도를 확인하기 위하여 MTT assay를 이용하였다. 상기와 동일한 방 법으로 처리된 세포를 대상으로 배지를 제거하고 성장배지와 희석된 0.5 mg/ml 농도의 tetrazolium bromide salt (MTT, Amresco, Solon, OH, USA)를 처리하고 빛을 차단한 다음 동 안 CO
2incubator에서 배양시켰다. 3시간 경과 후 MTT 시약을 제거하고 DMSO를 이용하여 formazan을 모두 녹인 다음 ELISA reader (Molecular Devices, Sunnyvale, CA, USA)를 이용하여 540 nm에서 흡광도를 측정하였다.
도립현미경을 이용한 세포의 성장과 형태의 관찰
EECO 처리에 따른 U937 세포의 성장 정도 및 형태에 어떠 한 변화가 유발되는 지를 확인하기 위하여 세포 배양용 6 well plate에 세포를 2×10
5개/ml의 개수로 well 당 2 ml씩 분주하고 EECO를 적정농도로 희석 처리하여 24시간 동안 배양한 다음 도립현미경을 이용하여 200배의 배율로 각 농도에 따른 성장 정도 및 형태의 변화를 관찰하고 Axio Vision 프로그램을 이 용하여 사진 촬영을 실시하였다.
DAPI staining에 의한 세포핵의 형태 관찰
Apoptosis가 유발되었을 경우 특이적으로 나타나는 핵의 형태적 변화를 관찰하기 위하여 EECO가 처리된 세포를 모은 다음 fixing solution (3.7% formaldehyde in PBS)으로 10분 동안 고정하였다. 고정된 세포를 PBS에 부유시킨 후 cytospin (Shandon, UK)을 이용하여 slide glass에 부착시키고 0.2%의 Triton X-100 (Amresco, Solon, OH, USA)을 첨가하여 상온에 서 10분간 고정한 다음 2.5 μg/ml 농도의 4',6-diamidino-2- phenylindole (DAPI, Sigma-Aldrich) 용액을 처리하여 상온 에서 15분간 염색하였다. 염색이 끝난 후 형광 현미경(fluores- cene microscope, Carl Zeiss, Germany)을 이용하여 400배의 배율로 핵의 형태 변화를 관찰한 다음 Axio Vision 프로그램 을 이용하여 사진 촬영을 하였다.
DNA 단편화 분석
Apoptosis 유발의 또 다른 증거를 제시하기 위하여 DNA 단편화 현상의 분석을 실시하였다. 상기와 동일한 방법으로 처리된 세포들을 모은 다음 lysis buffer [5 mM Tris-HCl (pH 7.5), 5 mM EDTA, 0.5% Triton X-100]를 첨가하여 상온에서 1시간 동안 처리한 다음 상층액을 회수하였다. 회수된 상층액 에 proteinase K solution (Sigma-Aldrich)을 처리하여 50℃에 서 3시간 동안 반응시킨 후 phenol : chloroform : isoamyl alcohol 혼합 용액(25:24:1, Sigma-Aldrich)을 첨가하고 30분간 회전교반 시킨 다음 원심분리하여 다시 상층액을 분리하였다.
분리된 상층액에 isopropanol (Sigma-Aldrich)과 5 M NaCl를 첨가하여 4℃에서 24시간 동안 반응시킨 후 원심분리시켜
DNA pellet을 추출하였다. 추출된 DNA pellet에 RNase A가 포함된 TE buffer 및 6X gel loading dye (Bioneer, Daejeon, Korea)를 첨가하고 1.6% agarose gel을 이용하여 50 V로 전기 영동 시킨 후 ethidium bromide (EtBr, Sigma-Aldrich)로 염색 하여 DNA 단편화 현상을 확인하였다.
Annexin V-Fluorescein Isothiocyanate (FITC)/
Propidium Iodide (PI) staining에 의한 apoptosis 분석 EECO가 유발하는 apoptosis 정도를 정량적으로 분석하기 위하여 정상 및 EECO가 24시간 동안 처리된 세포들을 모은 다음 원심분리하여 상층액을 제거한 후 PBS를 이용하여 세척 하였다. 준비된 세포를 10 mM HEPES/NaOH, pH 7.4, 140 mM NaCl and 2.5 mM CaCl
2가 포함된 annexin V binding buffer (Becton Dickinson, San Jose, CA, USA)에 부유시킨 다 음 annexin V-FITC 및 propidium iodide (PI, Becton Dickin- son)를 처리하여 암실에서 20분 동안 반응을 시켰다. 반응시킨 세포를 35-mm mesh를 이용하여 단일세포로 분리한 후 DNA flow cytometer (FACSCalibur, Becton Dickinson)에 적용시켜 apoptosis가 유발된 세포(V+/PI–)를 형광반응에 따라 측정 한 후 CellQuest software 및 ModiFit LT 프로그램을 이용하 여 분석하였다.
Mitochondrial membrane potential (MMP, Δψ
m)의 분석
EECO가 유발하는 apoptosis에 있어서 미토콘드리아의 기 능이상이 관여하는지를 확인하기 위하여 MMP 변화 정도를 확인하였다. 정상 및 EECO가 처리된 세포를 모은 다음 PBS에 부유시키고 5,5′, 6,6′-tetrachloro-1,1′,3,3′-tetraethyl-im- idacarbocyanine iodide (JC-1, Sigma) 용액을 첨가하여 상온 에서 20분 동안 반응시켰다. 반응이 끝난 후 원심분리를 이용 하여 상층액을 제거한 다음 차가운 PBS를 첨가하고 35-mm mesh를 이용하여 단일세포로 분리한 후 DNA flow cytometer 에 적용시켜 EECO 처리에 따른 MMP의 변화 정도를 분석하 였다.
Western blot analysis에 의한 단백질 발현 분석
준비된 세포에 lysis buffer [25 mM Tris-Cl (pH 7.5), 250
mM NaCl, 5 mM EDTA, 1% NP-40, 1 mM phenymethy-
lsulfonyl fluoride (PMSF), 5 mM dithiothreitol (DTT)]를 첨가
하여 반응시킨 후 원심분리를 이용하여 상층액에 있는 총 단
백질을 분리하였다. 단백질 농도는 Bio-Rad 단백질 정량 시약
(Bio-Rad, Hercules, CA, USA)을 이용하여 정량 한 다음 동량
의 Laemmli sample buffer (Bio-Rad)를 섞어서 균일한 농도의
단백질 sample을 만들었다. 이렇게 만들어진 단백질 sample
은 sodium dodecyl sulphate (SDS)-polyacrylamide gel을 이
용한 전기영동을 실시하여 분자량에 따라 분리한 다음 nitro-
A B
C
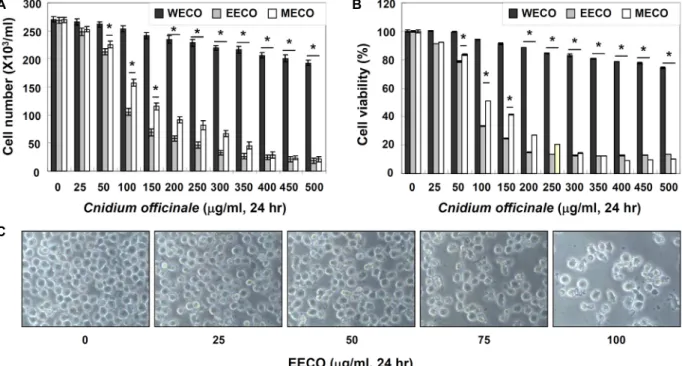
Fig. 1. Growth inhibition and morphological changes of human leukemia U937 cells. Cells were seeded at 2X105/ml in a 6-well plate and treated with variable concentrations of WECO, EECO and MECO. (A) The cells were trypsinized and the viable cells were scored by hemocytometer counts of trypan blue-excluding cells. (B) The growth inhibition was measured by the metabolic-bye-based MTT assay. The data are expressed as the mean ± SD of three independent experiments. The significance was determined by the Student’s t-test (*p<0.05 vs. untreated control). (C) Morphological changes of cells by treatment with EECO were visualized by inverted microscope (Original magnification, 200×).
cellulose membrane (Schleicher and Schuell, Keene, NH, USA)으로 electroblotting에 의해 전이시켰다. 단백질이 전이 된 nitrocellulose membrane에 skim milk 용액을 이용하여 비 특이적인 단백질들에 대한 blocking을 실시하고 1차 및 2차 항체를 처리하여 반응시켰다. 반응이 끝난 후 암실에서 en- hanced chemiluminoesence (ECL) slution (Amersham Life Science Corp.)을 적용시킨 다음 X-ray film에 감광시켜 특정 단백질의 발현양을 분석하였다.
In vitro caspases activity 측정
EECO에 의한 caspases 활성 정도를 확인하기 위하여 vitro caspases activity를 측정하였다. 준비된 세포에서 단백질을 추 출하고 정량한 다음 150 μg의 단백질이 함유된 50 μl의 sample 에 기질 100 μM이 함유된 reaction buffer [40 mM HEPES (pH 7.4), 20% glycerol (v/v), 1 mM EDTA, 0.2% NP-40, 10 mM DTT] 50 μl를 혼합하여 각 sample 당 총 volume이 100 μl가 되게 하였다. 여기에 세 종류의 caspases 기질(caspase-3 : Asp-Glu-Val-Asp (DEVD)-p-nitroaniline (pNA), caspase-8 : Ile-Glu-Thr-Asp (IETD)-pNA, caspase-9 : Leu-Glu-His-Asp (LEHD)-pNA)을 각각 5 μl 씩 첨가하여 37℃, 암실에서 3시간 동안 반응시킨 후 이용하여 405 nm 파장에서 caspases 활성 정도에 따른 변화를 측정하였다.
통계분석
상기의 모든 실험 결과는 SPSS ver. 18.0 통계 프로그램을 이용하여 평균(mean) ± 표준편차(SD)로 나타냈다. 각 실험군 의 분석 항목별 통계의 유의성 검증은 분산분석(Analysis of Vatiance, ANOVA)을 한 후, Student t-test를 이용하여
*p<0.05 수준에서 검증하였다.
결과 및 고찰
U937 세포의 생존율 및 증식에 미치는 WECO, EECO 및 MECO의 영향
U937 세포의 생존율 및 증식에 천궁 추출물인 WECO, EECO 및 MECO가 어떠한 영향을 미치는 지를 확인하기 위하 여 각각의 시료를 적정 농도로 24시간 동안 처리한 후 hemo- cytometer counting 및 MTT assay를 실시하였다. Fig. 1A에서 나타난 바와 같이 생존율 변화를 확인한 결과 WECO 처리군 에서는 큰 변화가 나타나지 않은 반면에 EECO 및 MECO 처 리군에서는 처리농도 증가에 따라 급격한 생존율의 감소가 관찰되었으며 특히 MECO에 비해서 EECO을 처리했을 경우 생존율의 감소가 더욱 강하게 유발되는 것으로 관찰되었다.
또한 Fig. 1B에서와 같이 U937 세포의 증식억제 정도를 확인
한 결과에서도 생존율 감소 현상과 같이 WECO 및 MECO
A B
C
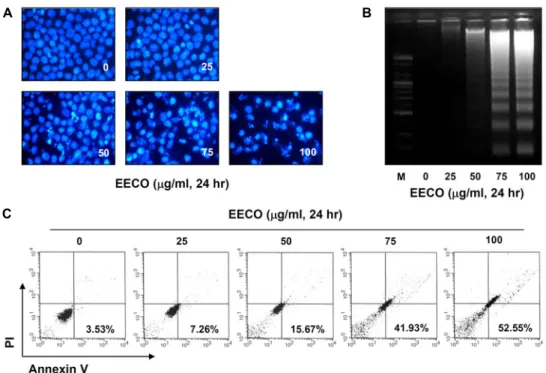
Fig. 2. Induction of apoptosis by EECO in human leukemia U937 cells. (A) The cells were fixed and stained with DAPI solution.
The stained nuclei were photographed with a fluorescence microscope using blue filter. (Original magnification, 400×). (B) The DNA fragmentations were separated on 1.6% agarose gel electrophoresis and visualized under UV light after staining with EtBr. (C) To quantify the degree of apoptosis induced by EECO, cells were stained with FITC-conjugated Annexin V and PI for DNA flow cytometry analysis. Apoptotic cells are determined by counting the % of annexin V+/PI– cells.
처리군에 비해서 EECO 처리군에서의 증식억제 현상이 강하 게 유발된다는 것을 알 수 있었다. 생존율 및 증식억제 현상을 가장 강하게 유발시키는 EECO가 세포의 형태 변화에 어떠한 영향을 미치는 지를 확인한 결과 Fig. 1C에서와 같이 처리 농 도 증가에 따른 전체적인 세포 밀도의 감소와 함께 apoptosis 특징 중 하나인 membrane blebbing 현상을 포함한 심한 형태 적 변형이 관찰되었다. 이상의 결과는 EECO 처리에 따른 생존 율 감소 및 증식 억제 정도와 잘 일치함을 알 수 있었다.
Apoptosis 유발에 미치는 EECO의 영향
EECO 처리에 따른 생존율 감소, 증식 억제 및 형태 변화가 apoptosis 유발과 관련이 있는 지를 확인하기 위하여 핵의 형 태변화, DNA 단편화 현상 및 annexin V-FITC/PI 염색을 실 시하였다. 먼저 핵의 형태변화를 확인하기 위하여 DAPI 염색 을 실시하고 형광현미경으로 관찰한 결과 Fig. 2A에 나타난 바와 같이 정상 배지에서 배양된 세포와는 다르게 EECO 처리 농도 증가에 따라 전체적인 핵의 밀도 감소와 더불어 apopto- sis가 유발된 세포에서 전형적으로 관찰되는 염색질 응축에 의한 apoptotic body의 형성이 관찰되었다. 다음으로 apopto- sis 유발의 또 다른 증거인 DNA 단편화 현상을 관찰하였다.
DNA 단편화는 apoptosis가 일어난 세포들에서 나타나는 전 형적인 현상으로서 핵 내의 endonuclease가 활성화되어 chro- mosomal DNA를 단편화시킴으로서 나타나는 것으로 알려져
있다[31]. 이를 위하여 정상 및 EECO가 함유된 배지에서 자란 세포를 대상으로 총 DNA를 추출하여 agarose gel 전기영동으 로 조사한 결과는 Fig. 2B에 나타난 바와 같다. 결과에서 볼 수 있듯이 EECO 25 μg/ml 처리군까지는 DNA 단편화 현상이 나타나지 않았지만 50 μg/ml 처리군에서 약한 DNA 단편화 현상이 관찰되기 시작하여 75 μg/ml 처리군에서부터 DNA 단편화 현상이 강하게 나타나는 것으로 조사되었다. 이러한 결과들을 살펴볼 때 EECO는 U937 세포에서 apoptosis를 유발 한다는 것을 알 수 있었으므로 이러한 apoptosis 유발의 정량 적인 분석을 위하여 annexin V-FITC/PI로 염색한 후 DNA flow cytometry 분석을 실시하였다. Fig. 2C에 나타난 바와 같이 정상배지에서 자란 세포의 apoptosis 유발 빈도는 약 3.5% 정도였지만 EECO 처리농도 증가에 따라 apoptosis 유발 빈도가 증가하여 최고 농도인 100 μg/ml 처리군에서는 약 52.5%의 세포에서 apoptosis가 유발되는 것으로 나타났다. 이 상의 결과를 살펴볼 때 U937 혈구암세포에서 EECO 처리에 의한 생존율 감소, 증식 억제 및 형태 변화는 apoptosis 유발과 밀접한 연관이 있음을 알 수 있었다.
MMP (Δψ
m)에 미치는 EECO의 영향
EECO가 유발하는 apoptosis에 있어서 미토콘드리아가 어
떠한 역할을 하는지를 확인하기 위하여 MMP를 조사하기 위
하여 EECO를 처리하고 dual-emission fluorescent dye인 JC-1
Fig. 3. Effects of EECO treatment on mitochondrial membrane potential (MMP, Δ
ψ
m) in human leukemia U937 cells.Cells were treated with EECO for 24 hr to follow the extent of apoptosis by determination of MMP using JC-1 reagent. Cells were analyzed on a DNA flow cytometer.
Dot plots of red vs. green fluorescence show live cells with intact MMP and dead cells with lost MMP, respectively.
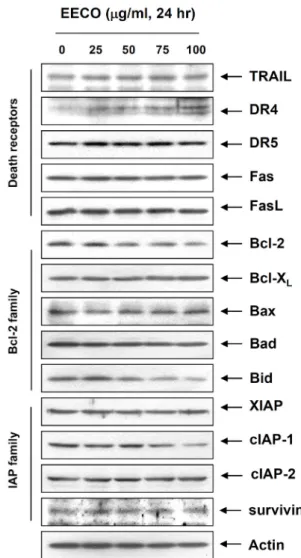
Fig. 4. Expression levels of apoptosis-related proteins by EECO treatment in human leukemia U937 cells. The cells were treated with indicated concentration of EECO for 24 hr.
The cells were lysed and then equal amounts of cell ly- sates (30 μg) were separated on SDS-polyacrylamide gels and transferred to nitrocellulose membranes. The mem- branes were probed with the indicated antibodies and the proteins were visualized using an ECL detection system. Actin was used as an internal control.
으로 염색한 후 DNA flow cytometer를 이용하여 조사하였다.
Fig. 3에 나타난 바와 같이 EECO를 처리하지 않았을 경우에는 정상적인 MMP를 가진 세포는 약 93.2%, 손상된 MMP를 가진 세포는 약 6.8%로 나타났지만 EECO 처리농도 증가에 따라서 MMP 손상 정도가 증가하였으며, 최고 농도인 100 μg/ml 처 리군에서는 약 74.5% 정도의 세포에서 MMP의 손상이 유발되 는 것으로 나타났다. 이상의 결과에서 EECO에 의하여 유발되 는 apoptosis에는 MMP의 손상을 통한 미토콘드리아 기능 이 상이 관여하고 있다는 것을 알 수 있었다.
Apoptosis 조절 단백질의 발현에 미치는 EECO의 영향 일반적으로 apoptosis는 extrinsic pathway 및 intrinsic pathway가 관여하며, 이러한 과정은 여러 가지 유전자들의 조절에 의하여 유발된다[9]. Extrinsic pathway를 통한 apop- tosis 유발에는 세포막에 존재하는 death receptor인 DR4, DR5 및 Fas에 apoptotic ligand인 TRAIL 및 FasL가 결합함으 로서 caspase-8을 활성화시켜 하위단계에 있는 caspase-3의 활 성화에 직접적으로 관여하거나 Bid의 단편화를 통한 trun- cated Bid (tBid)의 형성을 유발시킴으로서 intrinsic pathway 를 통한 apoptosis를 유발하는 것으로 알려져 있다[17, 23]. 한 편 Intrinsic pathway에 의한 apoptosis 유발에 중요한 역할을 하는 Bcl-2 family는 mitochondria 외막에 존재하고 있으며, Bcl-2 및 Bcl-
XL과 같은 apoptosis를 억제하는 anti-apoptotic member와 Bax 및 Bad와 같이 apoptosis를 유발하는 pro- apoptotic member로 구성되어 있다. 이들은 서로 dimer를 형 성하고 있지만 균형이 깨어지게 되면 mitochondria의 기능 이상을 유발함으로서 mitochondria로 내막에 존재하고 있는 cytochrome c가 세포질로 방출되어 하위단계에 있는 유전자
들을 조절하여 apoptosis가 유발된다[2, 19]. 또한 IAP family
의 경우에는 caspases와 직접적으로 결합함으로서 caspases의
활성을 억제함으로서 apoptosis를 억제하는 것으로 알려져 있
으므로 IAP family의 발현 억제는 caspases의 활성을 억제하
지 못함으로서 apoptosis가 유발되는데 중요한 역할을 하게
된다[4]. 따라서 EECO에 의하여 유발되는 apoptosis에 이들
유전자가 어떠한 영향을 미쳤는지를 확인한 결과 Fig. 4에 나
타난 바와 같이 extrinsic pathway에 관여하는 death receptor
인 DR4의 발현 증가 및 intrinsic pathway에 관여하는 Bcl-2의
발현 감소가 유발되었으며, 활성화된 caspase-8에 의하여 단편
화되어 MMP 손실 유발에 중요한 역할을 하는 것으로 알려진
A B
Fig. 5. Activation of caspases and the degradation of the caspase-3 substrate protein by EECO treatment in human leukemia U937 cells. The cells were treated with indicated concentration of EECO for 24 hr. (A) The cells were lysed and then equal amounts of cell lysates (30 μg) were separated on SDS-polyacrylamide gels and transferred to nitrocellulose membranes. The membranes were probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B) Caspases (-3, -8, and -9) activities were determined using in vitro caspases assay kits obtained from R&D according to the protocol of the manufacturer. The data are expressed as the mean ± SD of three independent experiments.
The significance was determined by the Student’s t-test (*p<0.05 vs. untreated control).
Bid 단백질의 경우에도 EECO 처리에 의하여 total Bid의 발현 이 감소하는 것으로 나타났으므로 단편화된 형태인 tBid의 증 가가 유발되었다는 것을 예상할 수 있었다. 또한 IAP family의 경우에는 XIAP, cIAP-2 및 survivin의 발현에는 큰 변화가 관 찰되지 않았지만, cIAP-1의 발현 감소가 관찰되었다. 이상의 결과를 살펴볼 때 EECO는 extrinsic pathway 및 intrinsic pathway가 모두 관여함으로서 apoptosis를 유발한다는 것을 알 수 있었다.
EECO 처리에 따른 caspases의 활성 및 기질단백질의 발 현변화
정상적인 세포에서 핵과 미토콘드리아 외막에 불활성 형태 인 proenzyme 형태로 존재하는 caspases는 caspase-8 및 -9과 같은 initiator caspase와 caspase-3, -6 및 -7과 같은 effector caspase로 구분되어지며, 다양한 세포 내외부 자극에 의하여 initiator caspases가 활성화되면 하위단계에 존재하는 effector caspases가 활성화 됨으로서 여러 종류의 기질 단백질들을 분 해함으로서 apoptosis를 유발하는 것으로 알려져 있다[6].
Caspases 활성화에 의하여 분해가 유발되는 대표적인 기질 단백질로는 PARP, β-catenin 및 PLC-γ1이 알려져 있으며, 이 들 중 PARP는 DNA의 결함을 인지하는 domain을 가지고 있 으므로 DNA 수선에 관여함으로서 세포의 유전적 안정성 유 지에 중요한 역할을 하지만 분해가 유발되면 DNA 수선 기능 을 상실하게 되는 것으로 알려져 있다[30]. 또 다른 기질 단백
질인 β-catenin은 세포들 사이의 연접에 직접적으로 관여함으 로서 세포의 골격 유지 및 다양한 부착성 세포의 전사 조절에 중요한 역할을 담당하며, PLC-γ1은 세포 증식에 중요한 역할 을 하는 것으로 알려져 있다[1, 8]. 따라서 EECO 처리에 의한 apoptosis 유발에 있어서 이들 유전자들이 어떠한 영향을 미 치는 지를 확인하였다. 먼저 caspases의 발현 정도를 확인한 결과 Fig. 5A에서 나타난 바와 같이 EECO 처리에 의하여 불활 성형인 pro-caspase-3, -8 및 -9 단백질의 발현은 감소하는 것으 로 나타났으나 활성형 단백질 발현은 나타나지 않았다. 따라 서 in vitro caspases activity assay를 통하여 이들 caspases의 활성 정도를 직접 분석한 결과, Fig. 5B에서 나타난 바와 같이 caspase-3, -8 및 -9의 활성이 EECO 처리 농도 의존적으로 증 가하여 최고 농도인 100 μg/ml 처리군에서는 각각 4.9배, 2.7 배 및 3.1배 증가하는 것으로 나타났다. 다음으로 활성화된 caspase-3에 의하여 특이하게 분해가 일어나는 기질 단백질의 발현에 미치는 EECO의 영향을 조사한 결과 Fig. 5A에 나타난 바와 같이 PARP, β-catenin 및 PLC-γ1의 발현 감소 또는 단편 화 현상이 관찰되었다. 이상의 결과를 살펴볼 때 인체 혈구암 세포인 U937 세포에서 EECO가 유발하는 apoptosis는 cas- pase-8 및 -9의 활성화에 따른 caspase-3의 활성화 및 기질단백 질들의 단편화가 중요한 역할을 한다는 것을 알 수 있었다.
EECO에 의한 apoptosis 유발에서 AMPK의 역할
AMPK는 지질과 포도당 대사의 조절인자로서 당뇨병 및
A B C
D E
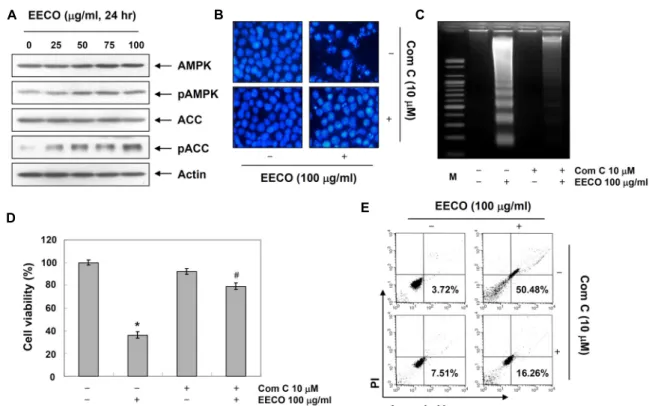
Fig. 6. Effects of AMPK inhibitor, compound C, on the EECO-induced in human leukemia U937 cells. (A) The cells were treated with EECO for 24 hr. Equal amounts of cell lysate were resolved by SDS-polyacrylamide gels, transferred to nitrocellulose membranes, and probed with the indicated antibodies. Proteins were visualized using an ECL detection system. Actin was used as an internal control. (B-E) The cells were pretreated with compound C for 1 hr and then treated with EECO for 24 hr. (B) After staining with DAPI solution, the nuclei were observed under a fluorescent microscope (Original magnification, 400×). (C) The DNA fragmentations were separated on 1.6% agarose gel electrophoresis and visualized under UV light after staining with EtBr. (D) Cell viability was determined by MTT assays. Each point represents the mean ± SD of three independent experiments. The significance was determined using Student’s t-test (*p<0.05 vs. untreated control; #p<0.05 vs. EECO-treated cells). (E) The cells were stained with FITC-conjugated Annexin V and PI for DNA flow cytometry analysis.
비만을 포함한 대사성 질환의 발생에 중추적인 역할을 담당하 고 있으므로 대사성 질환의 약물 타겟으로 많은 연구가 이루 어져 왔지만 최근에는 암세포의 증식을 억제하는 것으로도 알려지고 있으므로 이와 관련된 연구가 진행되고 있다[7, 11].
최근 연구에 따르면 AMPK는 암세포에서 cyclin dependent kinase inhibitor (CKI)인 p21을 유도하여 세포분열을 억제시 킬 뿐 만 아니라 Bcl-2 family의 발현을 조절하거나 p53의 인산 화를 유발시킴으로서 미토콘드리아의 기능이상을 유발하여 apoptosis를 일으키는 등 여러 경로를 통하여 인체 암세포의 억제에 관여하는 것으로 보고되어 있다[15, 25, 28]. 따라서 EECO에 의한 apoptosis 유발에서 AMPK의 활성화가 관여하 는 지를 알아보기 위하여 AMPK 및 AMPK의 하위단계에서 작용하는 ACC의 발현 변화를 살펴보고 AMPK의 활성을 억 제하였을 경우 EECO에 의하여 유발되는 변화에 어떠한 영향 을 미치는 지를 확인하였다. 먼저 AMPK 및 ACC의 발현 변화 를 확인한 결과 Fig. 6A에 나타난 바와 같이 EECO 처리에 의하여 AMPK 및 ACC의 인산화가 현저하게 증가되는 것으 로 나타났으므로 EECO에 의한 apoptosis 유발에 있어서
AMPK의 활성화가 관여한다는 것을 예상할 수 있었다. 다음 으로 AMPK 억제제인 compound C를 선처리하여 AMPK 경 로를 억제하였을 경우 EECO에 의한 apoptosis 유발에 어떠한 영향을 미치는 지를 조사한 결과 Fig. 6B~E에 나타낸 바와 같 이 EECO처리에 의하여 유발된 핵의 형태 변화, DNA 단편화 현상, 증식억제 및 annexin-V positive 세포의 증가 등이 com- pound C 선처리에 의하여 비교하여 현저하게 회복되는 것으 로 나타났다. 이러한 결과를 살펴볼 때 EECO는 AMPK의 활 성화를 직접적으로 유발한다는 것을 알 수 있었으며, 이러한 AMPK의 활성화는 apoptosis 조절 단백질들의 발현에 영향을 줌으로서 EECO에 의한 apoptosis를 억제한다는 것을 예상할 수 있으므로 추가적인 연구를 통하여 이와 관련된 구체적인 기전을 확인할 필요가 있을 것으로 판단된다.
이상의 결과를 종합해 보면 인체 혈구암세포인 U937 세포
에서 EECO에 의하여 유발되는 apoptosis는 DR4의 발현 증가
에 의한 caspase-8의 활성화와 연계된 Bid의 단편화 및 Bcl-2의
발현감소에 의한 미토콘드리아 기능 이상을 유발함으로서
caspase-9 및 -3의 활성화에 의한 기질단백질들의 단편화 등과
같은 전형적인 extrinsic 및 intrinsic pathway를 동시에 경유 하는 multiple apoptotic pathway에 의하여 조절되며, 이러한 과정에서 AMPK의 활성화가 중요한 역할을 한다는 것을 알 수 있었다.
감사의 글
이 논문은 2015년도 정부(미래창조과학부)의 재원으로 한 국연구재단의 지원을 받아 수행된 기초연구사업임(No. 2014 R1A1A1008460).
References
1. Bae, S. S., Perry, D. K., Oh, Y. S., Choi, J. H., Galadari, S.
H., Ghayur, T., Ryu, S. H., Hannun, Y. A. and Suh, P. G.
2000. Proteolytic cleavage of phospholipase C-gamma1 dur- ing apoptosis in Molt-4 cells. FASEB J. 14, 1083-1092.
2. Brunelle, J. K. and Letai, A. 2009. Control of mitochondrial apoptosis by the Bcl-2 family. J. Cell Sci. 122, 437-441.
3. Chen, S., Xiao, X., Feng, X., Li, W., Zhou, N., Zheng, L., Sun, Y., Zhang, Z. and Zhu, W. 2012. Resveratrol induces Sirt1-dependent apoptosis in 3T3-L1 preadipocytes by acti- vating AMPK and suppressing AKT activity and survivin expression. J. Nutr. Biochem. 23, 1100-1112.
4. de Almagro, M. C. and Vucic, D. 2012. The inhibitor of apoptosis (IAP) proteins are critical regulators of signaling pathways and targets for anti-cancer therapy. Exp. Oncol.
34, 200-211.
5. de la Cruz, J., Kim, D. H. and Hwang, S. G. 2014. Anti cancer effects of Cnidium officinale Makino extract mediated through apoptosis and cell cycle arrest in the HT-29 human colorectal cancer cell line. Asian Pac. J. Cancer Prev. 15, 5117-5121.
6. Fiandalo, M. V. and Kyprianou, N. 2012. Caspase control:
protagonists of cancer cell apoptosis. Exp. Oncol. 34, 165-175.
7. Fogarty, S. and Hardie, D. G. 2010. Development of protein kinase activators: AMPK as a target in metabolic disorders and cancer. Biochim. Biophys. Acta. 1804, 581-591.
8. Fukuda, K. 1999. Apoptosis-associated cleavage of beta-cat- enin in human colon cancer and rat hepatoma cells. Int. J.
Biochem. Cell Biol. 31, 519-529.
9. Fulda, S. and Debatin, K. M. 2006. Extrinsic versus intrinsic apoptosis pathways in anticancer chemotherapy. Oncogene 25, 4798-4811.
10. Han, S. I., Kim, Y. S. and Kim, T. H. 2008. Role of apoptotic and necrotic cell death under physiologic conditions. BMB Rep. 41, 1-10.
11. Hardie, D. G., Hawley, S. A. and Scott, J. W. 2006. AMP-acti- vated protein kinase--development of the energy sensor concept. J. Physiol. 574, 7-15.
12. Jeong, J. B., Ju, S. Y., Park, J. H., Lee, J. R., Yun, K. W., Kwon, S. T., Lim, J. H., Chung, G. Y. and Jeong, H. J. 2009.
Antioxidant activity in essential oils of Cnidium officinale ma-
kino and Ligusticum chuanxiong Hort and their inhibitory effects on DNA damage and apoptosis induced by ultra- violet B in mammalian cell. Cancer Epidemiol. 33, 41-46.
13. Jin, Z. and El-Deiry, W. S. 2005. Overview of cell death sig- naling pathways. Cancer Biol. Ther. 4, 139-163.
14. Kwak, D. H., Kim, J. K., Kim, J. Y., Jeong, H. Y., Keum, K. S., Han, S. H., Rho, Y. I., Woo, W. H., Jung, K. Y., Choi, B. K. and Choo, Y. K. 2002. Anti-angiogenic activities of Cnidium officinale Makino and Tabanus bovinus. J.
Ethnopharmacol. 81, 373-379.
15. Lee, D. H., Lee, T. H., Jung, C. H. and Kim, Y. H. 2012.
Wogonin induces apoptosis by activating the AMPK and p53 signaling pathways in human glioblastoma cells. Cell.
Signal. 24, 2216-2225.
16. Lee, S. H., Lee, J. H., Oh, E. Y., Kim, G. Y., Choi, B. T., Kim, C. and Choi, Y. H. 2013. Ethanol extract of Cnidium officinale exhibits anti-inflammatory effects in BV2 microglial cells by suppressing NF-κB nuclear translocation and the activation of the PI3K/Akt signaling pathway. Int. J. Mol.
Med. 32, 876-882.
17. Li, H., Zhu, H., Xu, C. J. and Yuan, J. 1998. Cleavage of BID by caspase 8 mediates the mitochondrial damage in the Fas pathway of apoptosis. Cell 94, 491-501
18. Lopez-Beltran, A., Maclennan, G. T., de la Haba-Rodriguez, J., Montironi, R. and Cheng, L. 2007. Research advances in apoptosis-mediated cancer therapy: a review. Anal. Quant.
Cytol. Histol. 29, 71-78.
19. Martinou, J. C. and Youle, R. J. 2011. Mitochondria in apop- tosis: Bcl-2 family members and mitochondrial dynamics.
Dev. Cell 21, 92-101.
20. Momcilovic, M., Hong, S. P. and Carlson, M. 2006. Mamma- lian TAK1 activates Snf1 protein kinase in yeast and phos- phorylates AMP-activated protein kinase in vitro. J. Biol.
Chem. 281, 25336-25343.
21. Okada, H. and Mak, T. W. 2004. Pathways of apoptotic and non-apoptotic death in tumour cells. Nat. Rev. Cancer 4, 592- 603.
22. Ozaki, Y., Sekita, S. and Harada, M. 1989. Centrally acting muscle relaxant effect of phthalides (ligustilide, cnidilide and senkyunolide) obtained from Cnidium officinale Maki- no. Yakugaku Zasshi 109, 402-406.
23. Peter, M. E. and Krammer, P. H. 2003. The CD95 (APO-1/
Fas) DISC and beyond. Cell Death Differ. 10, 26-35.
24. Shen, Q. W., Zhu, M. J., Tong, J., Ren, J. and Du, M. 2007.
Ca2+/calmodulin-dependent protein kinase kinase is in- volved in AMP-activated protein kinase activation by al- pha-lipoic acid in C2C12 myotubes. Am. J. Physiol. Cell Physiol. 293, C1395-1403.
25. Tiainen, M., Vaahtomeri, K., Ylikorkala, A. and Mäkelä, T.
P. 2002. Growth arrest by the LKB1 tumor suppressor: in- duction of p21(WAF1/CIP1). Hum. Mol. Genet. 11, 1497- 1504.
26. Tomoda, M., Ohara, N., Gonda, R., Shimizu, N., Takada, K., Satoh, Y. and Shirai, S. 1992. An acidic polysaccharide having immunological activities from the rhizome of Cnidium officinale. Chem. Pharm. Bull. (Tokyo) 40, 3025-3029.
초록:천궁 에탄올 추출물의 AMPK 활성화를 통한 U937 인체 혈구암세포의 apoptosis 유발
정진우
1․최영현
1,2․박 철
3*
(1동의대학교 한의과대학 생화학교실, 2동의대학교 항노화연구소 및 Blue-Bio 소재개발센터, 3동의대학교 자연생활
과학대학 분자생물학과)
천궁(C. officinale)은 예로부터 민간처방 약재로 사용되었으며, 항염증, 항산화, 항암 및 신생혈관억제 등의 효능 을 가지는 것으로 알려져 있다. 하지만 혈구암세포에서 apoptosis 유발과 관련된 분자생물학적 기전에 대해서는 명확히 밝혀져 있지 않다. 본 연구에서는 인체 혈구암세포인 U937 세포에서 천궁의 열수, 에탄올 및 메탄올 추출 물(WECO, EECO 및 MECO)이 유발하는 항암효과 및 항암기전을 조사하였다. 먼저 WECO, EECO 및 MECO가 유발하는 증식억제 정도를 조사한 결과 EECO가 가장 뛰어난 효능을 가진다는 것을 알 수 있었으며, 이러한 현상 이 apoptosis 유발에 의한 것임을 annexin-V 염색, apoptotic body 형성, DNA 단편화 및 MMP 소실 등을 통하여 확인하였다. EECO 처리에 의한 apoptosis 유발에는 DR4의 발현 증가와 함께 cIAP-1, Bcl-2 및 total Bid의 발현 감소가 관여하였으며, caspases-3, -8 및 -9의 활성화와 함께 caspases-3의 기질 단백질인 PARP, β-catenin 및 PLC γ1의 단편화도 관찰되었다. 또한 EECO는 AMPK signaling pathway를 활성화시키는 것으로 나타났으며, AMPK 억제제인 compound C를 이용하여 AMPK의 활성을 억제하였을 경우 EECO에 의하여 유발되었던 apoptosis가 현저하게 감소되는 것으로 나타났다. 이상의 결과를 살펴볼 때 인체 혈구암세포인 U937 세포에서 EECO에 의하 여 유발되는 apoptosis는 AMPK가 중요한 조절자로서 작용하는 것으로 생각된다.
27. Vilches Troya, J. 2005. Understanding cell death: a challenge for biomedicine. An. R. Acad. Nac. Med. (Madr) 122, 631-656.
28. Wu, S. J., Huang, G. Y. and Ng, L. T. 2013. γ-Tocotrienol induced cell cycle arrest and apoptosis via activating the Bax-mediated mitochondrial and AMPK signaling path- ways in 3T3-L1 adipocytes. Food Chem. Toxicol. 59, 501-513.
29. Yi, B., Liu, D., He, M., Li, Q., Liu, T. and Shao, J. 2013.
Role of the ROS/AMPK signaling pathway in tetrame- thylpyrazine-induced apoptosis in gastric cancer cells.
Oncol. Lett. 6, 583-589.
30. Yinjun, L., Jie, J., Weilai, X. and Xiangming, T. 2004.
Homoharringtonine mediates myeloid cell apoptosis via up- regulation of pro-apoptotic bax and inducing caspase-3- mediated cleavage of poly(ADP-ribose) polymerase (PARP).
Am. J. Hematol. 76, 199-204.
31. Yoshida, A., Pommier, Y. and Ueda, T. 2006. Endonuclease activation and chromosomal DNA fragmentation during apoptosis in leukemia cells. Int. J. Hematol. 84, 31-37.
32. Zeqiraj, E., Filippi, B. M., Deak, M., Alessi, D. R. and van Aalten, D. M. 2009. Structure of the LKB1-STRAD-MO25 complex reveals an allosteric mechanism of kinase activa- tion. Science 326, 1707-1711.