Enterobacter sp. YB-46의 myo-Inositol dehydrogenase 유전자 클로닝과 특성분석
박찬영1, 김광규2, 윤기홍1*
1우송대학교바이오식품과학전공, 2㈜디와이내츄럴
Received: May 16, 2018 / Revised: May 23, 2018 / Accepted: May 25, 2018
서 론
Inositol (1,2,3,4,5,6-cyclohexanehexol)은 6개의수산화기 의에피머화에의해 9 종류의입체이성질체로존재하며이 들 중 myo-inositol (cis-1,2,3,5-trans-4,6-cyclohexanehexol;
MI)이자연에가장풍부하게존재한다. 주로콩과식물과곡
류에서 인산의 저장물질인 phytic acid (myo-inositol hexakisphosphate)는 MI을함유하고있으며이들식물로부 터 MI이산업적으로생산된다. MI을제외한다른이성질체 는자연계에서희소량존재하는데이들중 D-chiro-inositol
(DCI)은고혈당증이나다낭성난소증후군증상을개선하며
[1], scyllo-inositol (SI)은알츠하이머증상을완화시키는효 과가있는것으로알려졌다[2].
MI을 유일 탄소원으로 하여 성장할 수 있는 세균으로 Aerobacter aerogenes (Enterobacter aerogenes로 재분류) [3], Bacillus subtilis [4], Geobacillus kaustophilus [5], Lactobacillus casei BL23 [6], Rhizobium leguminosarum [7], Salmonella typhimurium [8], Sinorhizobium fredii [9]와 S. meliloti [10]가알려졌으며이들의 MI 대사에관여 하는효소계와그유전자및발현조절기작이연구되었다. MI을 비롯하여 그 이성질체인 DCI 및 SI을 산화하는 dehydrogenase는해당 inositol에의해생산이유도되는것 으로알려졌다[5, 8, 11]. B. subtilis는 MI, DCI과 SI을유일 탄소원으로이용할수있으며이노시톨의대사과정이자세 Molecular Cloning and Characterization of myo-Inositol Dehydrogenase from Enterobacter sp. YB-46
Chan Young Park1, Kwang-Kyu Kim2, and Ki-Hong Yoon1*
1Food Science & Biotechnology Major, Woosong University, Daejeon 34606, Republic of Korea
2DYNatural Co. Ltd., Daejeon 34145, Republic of Korea
A bacterial strain capable of metabolizing myo-inositol (MI) and converting to other substances was iso- lated from soil of orchard. The isolate, named YB-46, was grown on minimal medium supplemented with MI as the sole carbon source and was presumed to belonging to genus Enterobacter according to the 16S rDNA sequence. Escherichia coli transformant converting MI into unknown metabolites was selected from a metagenomic library prepared with fosmid pCC1FOS vector. Plasmid was isolated from the transformant, and the inserted gene was partially sequenced. From the nucleotide sequence, an iolG gene was identified to encode myo-inositol dehydrogenase (IolG) consisting of 336 amino residues. The IolG showed amino acid sequence similarity of about 50% with IolG of Enterobacter aerogenes and Bacillus subtilis. The His-tagged IolG (HtIolG) fused with hexahistidine at C-terminus was produced and purified from cell extract of recom- binant E. coli. The purified HtIolG showed maximal activity at 45℃ and pH 10.5 with the highest activity for MI and D-glucose, and more than 90% of maximal activity for D-chiro-inositol, D-mannitol and D-xylose. Km and Vmax values of the HtIolG for MI were 1.83 mM and 0.724 µmol/min/mg under the optimal reaction con- dition, respectively. The activity of HtIolG was increased 1.7 folds by Zn2+, but was significantly inhibited by Co2+ and SDS.
Keywords: Enterobacter sp. YB-46, myo-inositol dehydrogenase, cloning, purification, characterization
*Corresponding author
Tel: +82-42-630-9742, Fax: +82-42-630-9389 E-mail: [email protected]
© 2018, The Korean Society for Microbiology and Biotechnology
히밝혀졌다[12]. B. subtilis의유전체에는 MI 대사에관여 하는여러단계의효소를암호화하는 iolABCDEFGHIJ 오 페론이존재한다[4, 13]. Fig. 1에나타낸바와같이 MI 대사 의첫단계는 iolG 유전자산물인 myo-inositol dehydrogenase (IolG)에의해 MI이 scyllo-inosose (SIS)로전환되는과정이 며이때 NAD+가조효소로작용된다[14]. 또한 IolG는 DCI 에도작용하여 1-keto-D-chiro-inositol을생성하고이는 iolI 유전자산물인 inosose isomerase에의해 SIS로전환된다. 둘째 단계에서는 iolE 유전자 산물인 2-keto-myo-inositol dehydratase가 탈수반응을 통해 SIS를 D-2,3-diketo-4- deoxy-epi-inositol (DKEI)로 전환한다. 세번째 단계에서 DKEI는 5-deoxy-D-glucuronic acid (5DG)로 가수분해되며, 이때는 iolD 유전자산물인 DKEI hydrolase가관여한다. 네 번째단계에서 iolB 유전자산물인 5DG isomerase에의해 5DG가 2-deoxy-5-keto-D-gluconic acid (DKG)로 전환된 후, 다섯번째단계에서 iolC 유전자산물인 DKG kinase에 의해 2-deoxy-5-keto-D-gluconic acid 6-phosphate (DKGP) 로인산화된다. 마지막으로, iolJ 유전자산물인 aldolase가 작용하여 DKGP를 dihydroxyacetone phosphate와 malonic semialdehyde로 분해하며, malonic semialdehyde는 iolA 유전자산물인 malonic semialdehyde dehydrogenase에의 해 acetyl-CoA와 CO2로분해된다. B. subtilis는 IolG외에도 inositol dehydrogenase 활성을갖는 IolX와 IolW를생산한 다. IolG는 MI과 DCI 뿐아니라 pinitol (3-O-methyl-DCI) 에도작용하지만 SI에는작용하지못하는데[15, 16], IolX와
IolW는 SI에특이적으로작용하여 SIS로전환시키고이과 정중에 NAD+와 NADP+를각각조효소로이용한다. 특히 IolX는 SI의이화대사에주요역할을하는것으로나타났고 반면에 IolW는 NADPH를산화하면서 SIS를 SI로환원시키
는역할을하는것으로추정되고있다[12].
식물과공생하는 S. fredii, S. meliloti와 R. leguminosarum 에서 MI의대사기능이알파를비롯한콩과식물의질소고 정이나뿌리혹의형성에영향을미치는것으로알려졌다[9, 15, 17]. S. fredii 및 S. meliloti [9, 10]의 iolG 및 iolE 유 전자와 효소의 기능이 밝혀졌고, R. leguminosarum에서 iolA 및 iolDEB 유전자기능이연구되었다[17]. S. meliloti 의이노시톨대사관련유전자(iolA 및 iolCDEB)는 MI 뿐 만아니라 DCI와 SI을유일탄소원으로한성장에도필요 하다고알려졌다. 또한플라스미드에존재하는 idhA (iolG) 유전자산물이 MI과 DCI의산화과정에관여하며 SI을 SIS 로전환시키는반응은 smc01163 (iolY) 유전자에의해암호 화된별개의 dehydrogenase (IolY)가관여하는것으로알려 졌다[11, 15]. 한편대부분의유산균과는달리 MI을탄소원 으로 이용하는 L. casei BL23은 B. subtills와 유사하게 iolTABCDG1G2EJK의 operon을 가지고 있으며 2종류의 iolG 유전자(iolG1, iolG2)가연이어배열된것이특징적이 다[6]. 내열성균인 G. kaustophilus HTA426도 MI, SI 및 DCI을유일한탄소원으로하여성장하며이들의대사에관 여하는 유전자는 iolIDEBCAJ 오페론과 3종류의 inositol dehydrogenases 유전자에해당하는 gk1897-1898-1899 오페
론으로존재한다[5]. 본연구에서는과수원토양에서 MI을
다른대사산물로전환하는미생물을분리하고 inositol 대사 에관여하는유전자를클로닝하였으며이들중 iolG 유전자 와효소의특성을조사하였다.
재료 및 방법
myo-Inositol 분해균의 분리
MI을대사하여다른대사산물로전환하는미생물을선발 하기위해충청북도와경상북도의과수원에서채취한토양 을생리식염수로희석한후 LB 평판배지(yeast extract, 5 g;
tryptone, 10 g; NaCl, 5 g; agar, 15 g; water, 1 L)에도말 하여 30℃에서배양하였다. 콜로니의모양이서로다른균 주를선별하여 MI (0.5%)이첨가된 LB 액체배지에접종하 고 3일간진탕배양한후배양상등액에존재하는물질을박 층크로마토그래피(Thin-layer chromatography, TLC)로분 석함으로써 MI이다른물질로전환된균주를탐색하였다. TLC는배양상등액을적정량취해 silica gel-precoated thin layer glass plate (Merck Kiesegel, 60F254, Merck Millipore Co., Germany)에점적하고 methanol, chloroform과증류수 Fig. 1. Inositol catabolic pathway in B. subtilis.
(5:5:1-v/v/v)의혼합용액으로전개한후이를 2% KMnO4용 액으로 5초간염색하고 열풍으로말려 관찰하였다. MI와 pinitol은 Sigma-Aldrich. (USA)로구입하였다.
분리균의 동정
그람염색을실시하여분리균의형태적특성을관찰하였 으며, LB 액체배지에서배양한균체를 0.85% NaCl 용액에 현탁한후제조사의지침에따라 API 20E와 API 50 CHE (Biomerieux, France) kits에접종하고 30℃에서배양하면서
1일과 2일째각각탄수화물이용능과생화학적특성을관찰
하였다. 분리균의 16S rRNA 유전자의염기서열을분석하기 위해서총유전체 DNA를주형으로하고, 5'-AGAGTTTGA TCCTGGCTCAG-3' (Escherichia coli 16S rRNA 유전자 염기서열 8−27)과 5'-GGTTACCTTGTTACGACTT-3' (E.
coli 16S rRNA 유전자염기서열 1,492−1,510)을 primers로
이용하여중합효소연쇄반응(PCR)을실시함으로써증폭된
DNA 단편을정제한후염기서열분석에사용하였다.
MI 대사 효소 유전자의 클로닝
분리균의배양균체로부터총유전체 DNA는 Biofact Co.
(Korea)의 Bacterial Genomic DNA Preparation kit를 사 용하여지침서에따라분리하였다. 유전자은행을제조하기 위해서는 Epicentre Biotechnologies (USA)의 CopyControlTM Fosmid pCC1FOSTM Library Production kit를다음과 같 이사용하였다. 분리된유전체 DNA를 pipetting하여물리 적충격을가함으로써적절한크기의단편으로(30−50 kb) 절 단한후 End-Repair Enzyme Mix (Epicentre Biotechnologies,
USA)를사용하여절단단편의평활화와 5’ 인산화를실시하
였다. 전기영동을수행하여약 40 kb 크기의 DNA 단편을 GELase로 추출한 후 Fast-Link DNA ligase를 사용하여 pCC1FOS vector와 ligation하였다. Ligated DNA를 70℃에 서 10분간 열처리하고 MaxPlax Lambda Packaging Eextracts와혼합하여최종적으로 E. coli EPI300으로형질 도입한후 chloramphenicol (12.5 μg/ml)이첨가된 LB 평판 배지에도말하였다.
iolG 유전자의 과잉발현 및 효소 정제
iolG 유전자를과잉발현시키기위해 pET23a의 T7 promoter 하단에도입하여재조합플라스미드를제조하였다. IolG의
카르복시말단에 His-tag이융합된형태로발현시키기위해
2종류의 primers P8iolG-F1 (TCAACATATGACTTTAAA AGCAGGTATTGTGG; 밑줄은 NdeI 위치)과 P8iolG-nR (TGAACTCGAGCTTGTAGAACGCAGGTTTCGCT; 밑줄 은 XhoI 위치)을제조하였다. 이들 primers로증폭된유전 자단편을 NdeI과 XhoI으로절단하여동일한효소로처리
한 pET23a(+)에도입하여재조합플라스미드 pE8LiG1을제 조하였다. His-tagged IolG (HtIolG)의유전자를과잉발현 시키기위해 pE8LiG1을 E. coli BL21 (DE3)에도입하여얻 은형질전환주를 LB 액체배지에접종하고 37℃에서하룻 밤진탕배양한후, 동일배지 300 ml을함유한 baffled flask 에 1%가되도록접종하였다. 37℃에서진탕배양하면서흡 광도(OD600)가 0.6에도달했을때 IPTG를 1.0 mM이되도록 첨가하고 25℃에서 10시간추가로진탕배양하였다. 원심분 리하여균체를회수하고 8 ml의 cell lysis buffer (50 mM NaH2PO4, 300 mM NaCl, 10 mM imidazole, pH 8.0)에 현탁하여초음파파쇄한후윈심분리하여균체파쇄상등액 을회수하였다. 이를동일한완충액으로평형화시킨 Ni-NTA 컬럼에 주입한 후 washing buffer (50 mM NaH2PO4, 300 mM NaCl, 20 mM imidazole, pH 8.0)로 세척하였다. HtIolG를 300 mM imidazole을함유한완충액으로추출하 고각분획의 IolG 활성분석과 SDS-PAGE 분석을통해활 성이높고단일단백질이포함된분획을골라 10 mM sodium phosphate 완충액(pH 8.0)에서투석하여정제효소로사용 하였다.
IolG 활성 측정
IolG의활성은 0.5 mM NAD+를조효소로하고 50 mM 기
질과 100 mM 완충액을포함한반응액에적정량의효소를
첨가하여 45℃에서 30분간반응한후 340 nm에서흡광도
의증가에따른 NADH의생성량을측정함으로써결정하였
다. 효소활성에미치는반응온도의영향을조사하기위하 여 25−50℃와 pH 6.0−11.5의범위에서 IolG 활성을각각측 정하였다. 열안정성은 25−50℃범위의온도에서 IolG 효소 액을 30분동안방치한후잔존활성을측정하여결정하였 다. 금속이온과화합물이효소활성에미치는영향을조사하
기위해서는최종농도가 5 mM이되도록반응액에첨가하
고효소활성을측정하였다.
결과 및 고찰
MI 이용균의 분리와 동정
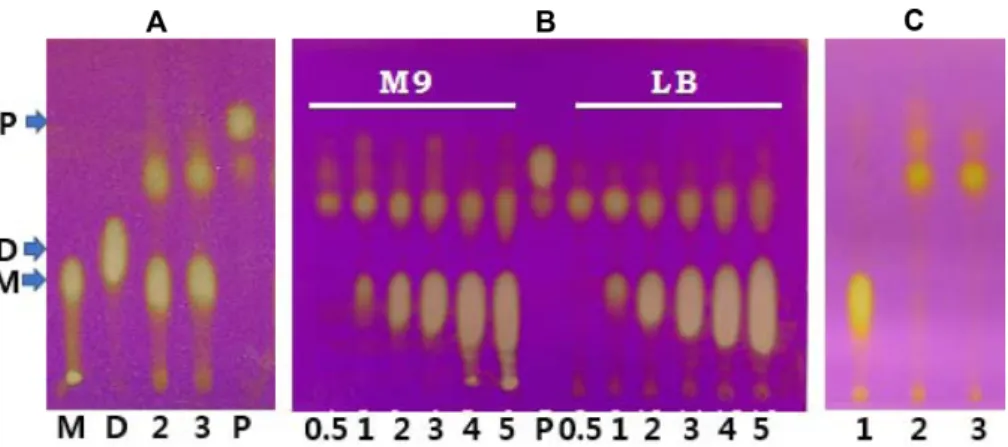
자연계에풍부하게존재하는 MI을유일탄소원으로이용 하는토양균이다수보고된바있다. MI의대사능이있는미 생물을탐색하기위해과수원토양으로부터분리된미생물 을 MI이첨가된 LB 배지에배양하여배양산물을 TLC로분 석하였다. 다수의토양세균이 MI을이용하지못하였으며,
분리균 YB-46은배지의 MI을이용하여미지의대사물질을
생산하는것으로나타났다(Fig. 2A). MI의첨가량을달리하 여배양한후 MI의이용정도를조사한결과 MI이 0.5% 첨 가된배지에서는 MI이완전히이용되어미지의물질이생산
되었으나, MI의첨가량을증가시켰을경우 MI은일정량만 소모되고배지에잔존하였으며생성된대사물질의양도크 게증가하지않았다(Fig. 2B).
분리균 YB-46의형태적특성을관찰한결과그람음성간
균으로확인되어 API 20E와 API 50CHE kit를사용하여생 화학적특성과당이용능을조사하였다. 그결과 YB-46은β- galactosidase의활성과 citrate의이용능을보였고 indole과 유기산을 생성하였으며, arginine dihydrolase, lysine decarboxylase, ornithine decarboxylase, urease와 tryptophan deaminase 활성및 acetoin과황화수소의생성능은모두없 었다. 당이용능은 Table 1에보인바와같으며, Biomeriux 의 API web (https://apiweb.biomerieux.com/jsp/login.jsp)
에서 다른균주와 생화학적특성을비교한 결과 YB-46은
Pantoea sp. 2와유사도가 87.3%로가장높았다. 한편 PCR 로 16S rDNA 단편을증폭하고 1,510 bp 크기의염기서열을 결정한후미국 NCBI의 BLAST 검색방법으로기존에등록 된세균들의상응하는염기서열과비교한결과 Enterobacter bugandensis와 1510 bp 중 1495 bp가동일하여상동성이 가장 높았으며, Enterobacter cancerogenus 및 Klebsiella oxytoca와 1494 bp가동일하였다. 한편생화학적특성에서 가장유사도가높았던 Pantoea 속균주중에서는가장높은
상동성을보이는균주는 Pantoea agglomerans JCM 1236 이며 1451 bp 중 1427 bp가동일하여 Enterobacter 속균주 보다상동성이낮았다. 이와같이조사된결과로볼때분리
균 YB-46이 확실하게 동정된 상태라 볼 수 없으나 16S
rDNA 서열에근거하여분리균을 Enterobacter sp. YB-46으 로명명하였다.
탄소원으로 glucose, fructose와 MI이각각 1% 첨가된 M9 최소배지에분리균 YB-46을접종하여 30℃에서 1일간진탕 배양한결과 glucose와 fructose가첨가된배지에서는순조 롭게성장하였고 MI이첨가된배지에서는거의성장이일어 나지않았으나배양 2일째는성장도가 glucose와 fructose가 첨가된배지와유사한수준을보였다. 이로보아 YB-46은 MI 을유일탄소원으로이용하여성장하지만유도기가 24시간 이상으로길었으며, Salmonella enterica도최소배지에서 MI을유일탄소원으로사용하였을때 40−60시간의유도기 를갖는것으로알려졌다[8, 18].
MI 대사 효소의 유전자 클로닝
MI의대사과정에는여러효소가관여하며그유전자는미 생물에따라차이는있지만대부분이오페론으로존재한다 [6, 8, 11, 13]. 그러므로 Enterobacter sp. YB-46으로부터 Fig. 2. Thin-layer chromatogram of culture filtrates by the isolate YB-46 and E. coli carrying pMU118. In panel A, the isolate YB- 46 was grown in LB broth supplemented with 2% myo-inositol (MI) for 3 days. Lanes M, MI; D, D-chiro-inositol; P, pinitol; 2 and 3, culture filtrates on 2nd and 3th day. MI, D-chiro-inositol and pinitol were indicated by arrows. In panel B, the isolate YB-46 was grown in M9 minimal medium and LB medium, respectively, supplemented with MI of various concentrations (%) indicated in the bottom of panel.
Lane P is pinitol. In panel C, E. coli carrying pCC1FOS (lane 1) or pMU118 (lane 2) and the isolate YB-46 (lane 3) were grown in LB broth containing 0.5% myo-inositol for 3 days, respectively.
Table 1. Utilization of carbohydates of Enterobacter sp. YB-46 in API 50CHE kit.
Fermented carbohydrates Non-fermented carbohydrates
Glycerol, D-Arabinose, L-Arabinose, Ribose, D-Xylose, Galactose, D-Glucose, D-Fructose, D- Mannose, L-Sorbose, Rhamnose, Dulcitol, Inositol, Mannitol, Sorbitol, Methyl-α-D-mannopy- ranside, Methyl-α-D-glucoside N-Acetyl-glucosamine, Amygdalin, Arbutin, Esculin, Salicin, Cellobiose, Maltose, Lactose, Melibiose, Saccharose Trehalose, D-Raffinose, β-Gentiobiose D-Turanose L-Fucose, D-Arabitol, Gluconate, 2-Keto-gluconate, 5-Keto-gluconate
Erythritol, L-Xylose, Adonitol, Methyl-β-D-xylopyranside, Inulin Melezitose, Amidon, Glycogen, Xylitol, D-Lyxose, D-Tagatose, D-Fucose, L-Arabitol
MI의대사에관련된유전자를클로닝하기위해크기가큰 DNA 단편의 삽입에적합한 pCC1FOSTM fosmid vector로 metagenomic library를제조함으로써약 700개의형질전환 주얻었다. 임의로 5개의형질전환주로부터재조합플라스 미드를분리한후여러종류의제한효소로절단하여삽입 된크기를예측한결과약 40 kb 크기의 DNA 단편이삽입 된것으로판단되었으며이로보아 YB-46의 metagenomic library는총 28 Mb 수준의유전체를함유한것으로추정 된다.
MI의대사유전자를함유한형질전환주를효과적으로탐 색하기위해 1% MI이첨가된 LB 액상배지에 10개의형질 전환주를동시에접종하고혼합배양한후배양상등액을 TLC로분석하여 1개집단의배양액에서 MI이소모되고미 지의대사산물이생성된것을확인하였다. 그러므로이집단 에포함된 10개의형질전환주를각각동일한배지에서배양 한후배양상등액을분석함으로써최종적으로 MI을대사하
여분리균 YB-46과동일하게미지의대사산물을생산하는
단일한형질전환주를선발하였다(Fig. 2C).
iolG 유전자의 염기서열과 유전자 산물
MI을대사하는형질전환주로부터플라스미드를분리하여
pMU118로명명하고삽입된유전자의염기서열을분석하기
위해 EcoRI, PstI, BamHI과 HindIII로각각절단된단편들 중클로닝된유전자로부터유래된단편을추출하고이를동
일한효소로절단된 pUC19에도입하여그염기서열을결정
하였다. 그결과 pMU118에클로닝된 DNA의부분적인염 기서열이결정되었으며밝혀진염기서열을기반으로하여
orf를조사한결과 336 아미노산잔기로구성된단백질을코 드하는 1,011 bp 크기의 iolG 유전자가관찰되었다(Fig. 3).
개시코돈 ATG로부터 5 nucleotides가떨어진지역에 purine 염기가집중적으로배열되어있었으나 RBS의전형적인염
기배열인 AGGAGG와는차이가있었다.
유전자염기서열로부터유추된 YB-46 유래 IolG (YBIolG) 의아미노산배열을미국의 NCBI database에등록된다른 효소와비교한결과 S. enterica의 inositol 2-dehydrogenase (WP_000172704)와상동성이 93%로가장높았으며 E. coli ED1a (CAR06517)와 Citrobacter werkmanii (WP_085048442) 의 IolG와도 92%의높은상동성을보였다. 이들효소는모 두 YBIolG와동일하게 336 아미노산잔기로구성되어있는 데효소의특성에대해보고된바는없다. 한편효소의특성 이 조사된 E. aerogenes (YP_007389612; 337 residues;
51.2%) [3], B. subtilis (NP_391849; 344 residues; 49.7%) [14], S. meliloti 1021 (NP_437734; 330 residues; 25.8%) [10], S. fredii USDA191 (AF274867; 329 residues; 25.2%) [9]와 L. casei BL23 (NC_010999; 350 residues; 19.6%) [19]의 IolG는 YBIolG와크기에차이가있을뿐아니라아 미노산배열의상동성이 19.6−51.2% 수준으로높지않았다 (Fig. 4). 내열성균인 G. kaustophilus HTA426의 IolG (NC_006510)는 397 아미노잔기로구성되어다른균의효 소에비해크며 YBIolG와도상동성이 17.9%로매우낮았다 [5].
YBIolG의 9−15번째아미노잔기는 dehydrogenases의 N
말단영역에서 NAD(H)의 ADP 부분에결합에관여한다고
알려진아미노산배열(GXGXXG)과동일하였다[20]. 또한 B.
Fig. 3. Nucleotide sequence and deduced amino acid sequence of iolG gene. The amino acid deduced from the open reading frame is shown with the one-letter code below the nucleotide sequence. Putative ribosome binding site (SD) is depicted below the underlines.
The numbers at the end of each line correspond to the nucleotide positions.
subtilis IolG의 233−235번째잔기인 YGY는기질인 inositol 의인식부위주변에서소수성공간을형성하는데 YBIolG 는 232−234번째 잔기에서이와 동일한 배열을 보였고 L.
casei BL23의 IolG2는 HGY로배열되어다른균의효소와 는 차이가 있었다. B. subtilis의 IolG에서 K97, D172와 H176은활성잔기이며 H176은산-염기촉매제작용을하는 것으로보고되었는데[21], YBIolG는 K97과 H176 잔기가일 치하였으나 172번 잔기는 Asp와 유사한 아미노산인 Glu (E172)로치환되어있었다. 한편 S. fredii, S. meliloti와 L.
casei BL23의 IolG 효소에서는이들활성잔기가 B. subtilis 의 IolG과동일하게배열되어있었으며 E. aerogenes의 IolG 는 172번잔기로 Asp와는특성이다른 Asn (N172)을함유 하였다.
재조합 대장균이 생산하는 IolG의 정제 및 특성
YBIolG 카르복시 말단에 His tag을융합한 His-tagged
YBIolG (HtIolG)의 유전자가 과잉 발현되도록 제조된
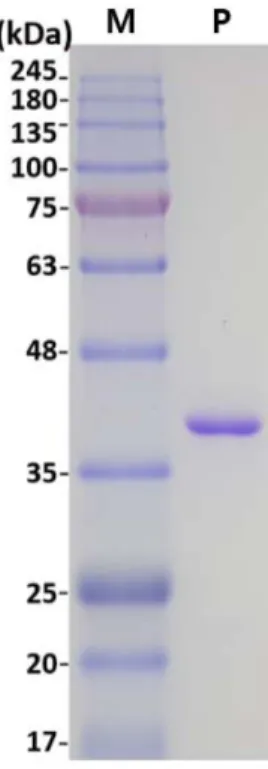
pE8LiG1을 E. coli BL21 (DE3)에도입하고그형질전환주 를 IPTG로처리하여 HtIolG의유전자를과잉발현한결과 IPTG 첨가후 37℃에서배양하였을때발현된 HtIolG이대 부분불용성단백질로존재하였으나 25℃로배양하였을때 는수용성단백질로발현된양이증가하였다. 그러므로 IPTG 첨가후 25℃에서배양된균체를파쇄하고 Ni-NTA 컬럼크 로마토그래피를 수행하여 HtIolG를 정제하였다. 정제된 HtIolG를 SDS-PAGE로분석한결과분자량이약 38−40 kDa 으로나타났으며(Fig. 5), 이는 iolG 유전자의염기서열로부 터예측된 YBIolG와카르복시말단에융합된 6개의 His 잔 기를포함한 HtIolG의분자량(38 kDa)과유사하였다. 한편 SDS-PAGE에서 39 kDa의분자량을 보이는 B. subtilis의 IolG는 분자량이 150−160 kDa인 사량체로 존재하며[16, 21], L. casei BL23의 IolG2도사량체로보고되었다[19].
정제된 HtIolG를사용하여반응온도와 pH에따른효소 활성을조사한결과최적반응온도는 45℃이며 pH 10.5에 서최대활성을보였고 pH 8.0에서약 25%, pH 11에서약 Fig. 4. Comparison of the Enterobacter sp. YB-46 IolG with others. The amino acid sequences of six IolGs from Enterobacter sp. YB- 46 (YB), S. fredii USDA191 (Sf), L. casei BL23 (Lc), S. meliloti 1021 (Sm), B. subtilis 168 (Bs) and E. aerogenes EA1509E (Ea) are given in the one-letter code and have been aligned by introducing gaps (hypens) to maximize similarities by Clustal W method. Putative amino acid sequence corresponding to NADH-binding motif is underlined. The catalytic triad residues are marked by closed circles and tyrosine- glycine-tyrosine motif for the hydrophobic pocket are marked by asterisks on the amino acids, respectively. Numbers at the beginning of each line correspond to the amino acid position in the protein.
70% 이상의활성을나타냈다(Fig. 6). Acetomonas oxydans 와 Gluconobacter oxydans ATCC 621H의 IolG는각각 pH 6.2와 pH 8.75에서최대활성을보였고 G. oxydans의 IolG 는 pH 5 이하와 10.5 이상에서는활성이거의없었으며[22, 23], B. subtilis의 IolG는 pH 9.0에서최대활성을보이고 pH 10.5에서는최대활성의 30% 수준을보였다[16]. 이로보아
YB-46의 HtIolG는다른균의효소에비해알칼리조건에서 활성이높은것을알수있다. 열안정성을조사하기위해 25− 50℃범위에서 30분간방치한후에 HtIolG의잔존활성을조 사하였다. 30℃이하에서는대부분의활성을유지하였으며 40℃이상에서실활되기시작하였고 50℃에서는 10%의매 우낮은잔존활성을보였다(Fig. 6B). 한편고온성균인 G.
kaustophilus의 GK-1897과 GK-1899는 NAD+를조효소로
사용하여 MI을산화하는 IolG로확인되었으며최적반응온
도가각각 55℃와 75℃이고 30분간열처리하였을때 55℃
이하에서전혀실활되지않아 HtIolG에비해열안정성이높
았고특히 GK-1899는 75℃에서도 60%의높은잔존활성을 보였다[5].
기질특이성을조사하기위해서는 NAD+의농도를 0.5 mM
로하고여러종류의당을동일한농도(50 mM)로사용하여
반응을 실시한 결과 HtIolG는 MI과 D-glucose에 대해 0.629 μmol/min/mg으로동일하게최대활성으로보였으며 DCI, D-mannitol 및 D-xylose에도 90% 이상의활성을나타 냈다. 그러나 D-glucose의에피머인 D-galactose (57.5%)와 D-mannose (38.7%)에대해서는활성이낮았으며 2-deoxy- glucose에대한 활성도는 47.2%로나타났다. B. subtilis의 IolG는 MI에대한활성이가장높았으며, D-glucose (25%), D-xylose (12.5%), D-ribose와 D-fructose (2%)에대한활성 은 MI에비해낮았고 D-galactose와 2-deoxy-glucose에대한
활성을없는것으로보고되어 HtIolG와큰차이가있었다
[16]. 한편 B. subtilis의 glucose dehydrogenase는 D- glucose에비해 2-deoxyglucose에대한활성이 108% 수준으 로가장높았으나 MI에대한활성은없는것으로알려졌다 [24]. 이로보아 YBIolG는 MI 뿐아니라 D-glucose에도활성 이높은 myo-inositol dehydrogenase로판단된다. IolG의최 Fig. 5. SDS-PAGE of the purified HtIolG. Lane P, the purified
enzyme; Lane M, the molecular weight markers shown in kilo- daltons to the left side of the gel.
Fig. 6. pH and temperature profiles for activity and stability of the purified HtIolG. In panel A, the pH profiles were obtained by measuring the IolG activities at various pH’s with a constant temperature of 45℃. Buffers (100 mM) used were as follows: sodium phos- phate (-●-, pH 6−8), Tris (-□-, pH 8−9), glycine-NaOH (-■-, pH 9−11), and NaOH-sodium phosphate (-△-. pH 11−11.5). In panel B, tem- perature profiles (-●-) were obtained by measuring the IolG activities at different temperatures with a fixed pH 8.0.
Thermostability (-□-) was determined by measuring the residual activities after pre-incubation for 30 min at different temperatures.
Each curve represents the average of three independent experiments within standard errors of 2% between of them.