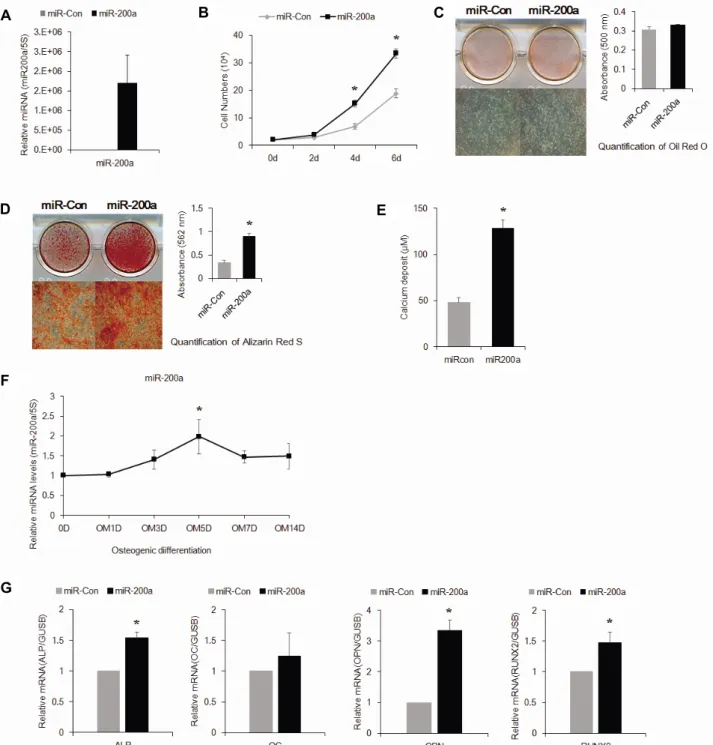
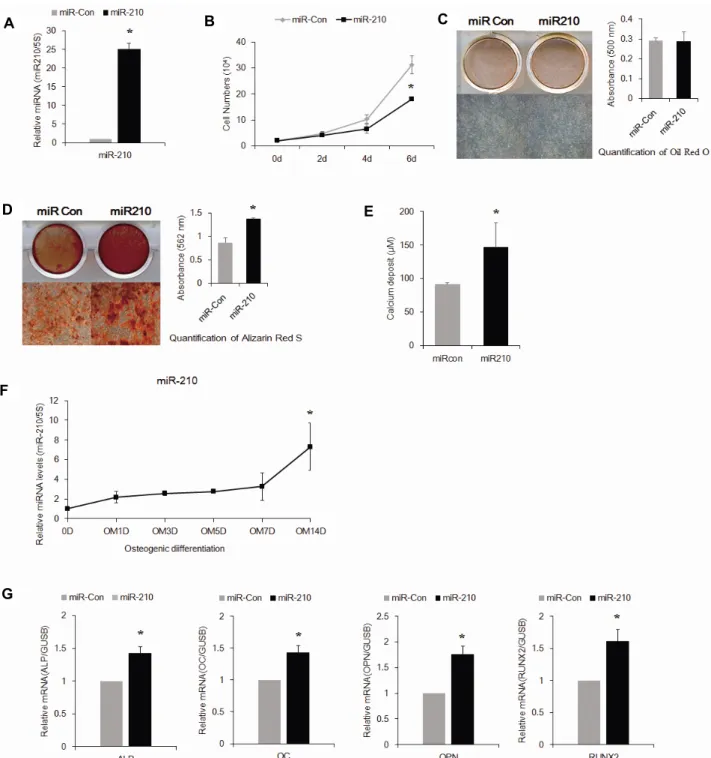
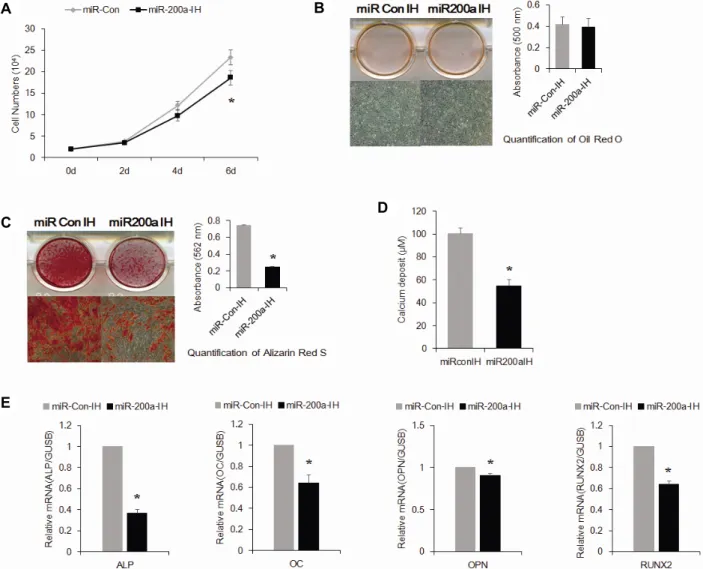
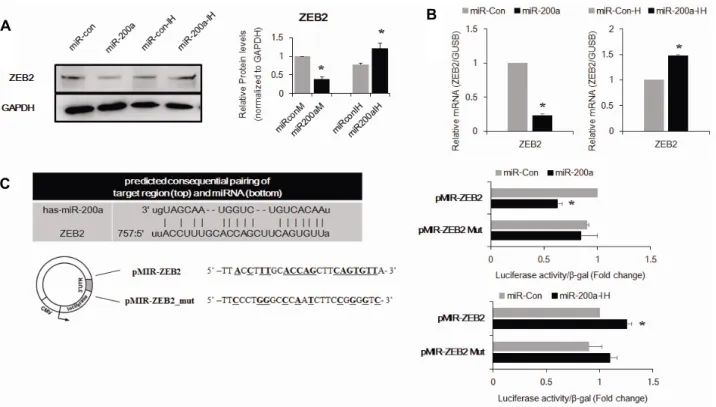
MicroRNA-200a/210 Controls Proliferation and Osteogenic Differentiation of Human Adipose Tissue Stromal Cells
Young Suk Kim
1, Hee Jeong Park
1, Keun Koo Shin
1, Sun Young Lee
1, Yong Chan Bae
2and Jin Sup Jung
1*
1
Department of Physiology, School of Medicine, Pusan National University, Yangsan 626-870, Korea
2