IEG 환경지질연구정보센터
14
0
0
전체 글
(2) 174. Thomas Wotte. Fig. 1. (a) Simplified geological map of the Iberian Peninsula (modified after Liñán et al., 2002); marked is the area of interest. (b) Generalised geological map of the Cantabrian Zone (modified after Pérez-Estaún et al., 1988). Abbreviations: Ge, Genestosa section; BL, Los Barrios de Luna section; Ve, Vegaservera section.. 1965; Van den Bosch, 1969; van Staalduinen, 1973; Crimes et al., 1977; Van der Meer Mohr, 1983; Truyols et al., 1990; Rodrígues Fernández et al., 1991; Aramburu et al., 1992). The rocks of the Herrería Formation are conformably overlain by the Láncara Formation (described below). The transition between the limestones of the Láncara Formation and the Oville Formation is characterised by a successive increase of siliciclastic material resulting in a unit of green shales and glauconitic sandstones with multiple facies changes. This third sedimentary succession is characterised by the disappearance of the carbonatic sedimentation in the whole Iberian Massif (Fig. 2). According to Truyols et al. (1990), the 80 m to 800 m thick Oville Formation shows several quartzitic beds towards the top. It is interpreted as shallow platform deposits of intertidal and braid plain deltaic environments (Aramburu et al., 1992; Aramburu and GarcíaRamos, 1993). Cambrian small shelly fossils from Spain are mainly known from central and southern part of the Iberian Peninsula. While molluscs, sponges and some problematic fossils are mentioned from those areas (e.g., Sdzuy, 1969; Fernández-Remolar, 2001; Vidal et al., 1999), only few organisms are reported from the Cantabrian zone. This micro-. fauna includes chancellorids (Archiasterella), sponge spicules (Eiffelia), brachiopods (?Acrotretida), molluscs (Helcionella?, Leptostega irregularis, Yochelcionella?, Scenella, Pelagiella crassa), and problematic fossils (Hadimopanella oezgueli) from the Barrios facies of the upper member of the Láncara Formation (e.g., Sdzuy, 1969; Van den Boogaard, 1983; Geyer, 1986). The calcareous succession of the upper member of the Láncara Formation was microfacial processed in detail for the first time to improve the knowledge of stratigraphic ranges, taxonomy, and palaeobiology of the microfauna of the Lower–Middle Cambrian interval for this part of the European Gondwana shelf. In addition, the microfauna permits to reconstruct the character and structure of the depositional areas and environments. Therefore the Beleñoand the Barrios facies of three sections of the Somiedo-Correcilla unit were intensively sampled. The underlying dolomites and grey limestones of the lower member as well as the boundary to the overlying siliciclastic Oville Formation were mapped although not investigated in detail in the scope of this paper. About 140 samples were analysed by point counting on thin sections, like described by Wotte et al. (2004). Chemical preparation (by acetic acid and formic.
(3) Facies distribution patterns and environment reconstruction of the Láncara Formation. Fig. 2. Simplified stratigraphic column of the Cambrian in the Cantabrian zone; (1) claystone, (2) sandstone, (3) dolomitic sandstone, (4) dolostone, and (5) limestone.. acid) of fossil remains from the carbonates supported these examinations on a small scale. However, the results presented here are principally base on the detailed examinations of the thin sections. 2. GEOLOGICAL SETTING AND SEDIMENTARY DEVELOPMENT The Láncara Formation consists of two members, differing in lithology and fossil content. The lower member (110 to 220 m in thickness) is composed of yellow dolostones and grey limestones. Birdseyes, algal mats, mud cracks, and tepee-structures are characteristic for these beds. The bathymetric conditions during the deposition of the Láncara For-. 175. mation were first studied in detail by Zamarreño (1972, 1975) and Aramburu et al. (1992). Therefore the environment of the lower member of the Láncara Formation is interpreted as a ramp environment characterised by supralitoral–litoral conditions in the western part, and shallow sublitoral conditions in the eastern part. The upper member of the Láncara Formation (10 to 50 m in thickness) includes two different facies-types, the Beleño facies and the Barrios facies. The Beleño facies is characterised by granular, crystalline, grey to roan dolomitic limestones with a high content of glauconite, and disarticulated echinoderm ossicles. The high abundance of echinoderm remains is responsible for the typical sparitic character of this lithotype. The Barrios facies is composed of three different types of carbonates (Zamarreño, 1972). The first type is represented by a glauconitic limestone (similar to that of the Beleño facies). On top of this glauconitic limestone a few meters of pink carbonates with stromatactis-like structures and red nodular limestones follow. The nodular limestone represents the major part of the upper member and is characteristic for the Barrios facies. A high fossil content and a very diverse fauna are typical for these lithotypes. Between the lower and the upper member of the Láncara Formation a discontinuity is placed that shows distinct evidence of erosion. Within the Esla nappe this boundary is marked by a hard ground level (Álvaro et al., 2000). In most parts of the Cantabrian zone this horizon is eroded. Thus only some reworked clasts were found in the lowermost part of the Beleño facies. This facies probably reflects the transgressive character during the earliest Middle Cambrian, which can also be observed in other environments of this time interval (Bechstädt and Boni, 1989, 1994; Álvaro and Vennin, 1996, 1997; Álvaro et al., 1999). The transition between the Beleño and the Barrios facies is interpreted as another discontinuity that marks a major tectonic impulse and the input of siliciclastic material into the environment (Álvaro et al., 2000). In some areas this contact is sharp (Álvaro et al., 2000), but can also be gradual. Such a gradual transition is detected in all the sections investigated here. However, this discontinuity can be observed in the whole southwestern Europe and is interpreted as the tectonic breakdown of the platforms of the Mediterranean at this time (Bechstädt et al., 1988; Bechstädt and Boni, 1989; Aramburu et al., 1992; Russo and Bechstädt, 1994; Álvaro and Vennin, 1996; Álvaro et al., 2000). 3. LITHOLOGICAL CHARACTERISTICS OF THE UPPER MEMBER OF THE LÁNCARA FORMATION IN THE SOMIEDO-CORRECILLA UNIT As described above, the Beleño- and the Barrios facies were intensively sampled from three sections. Their lithological successions are documented below..
(4) 176. Thomas Wotte. 3.1. Genestosa Section The locality Genestosa (Fig. 1b) is situated in the northern part of the Somiedo-Correcilla unit south of the eponymous village. Zamarreño described this section in 1972, but she documented the outcrop incompletely (22 m of the upper member of the Láncara Formation and about 79 m of the Oville Formation). At Genestosa, the transitions between the lower and the upper member of the Láncara Formation and from the latter into the siliciclastic Oville Formation are excellently exposed (Pl. 1.1). The maximum thickness of the Láncara Formation in the outcrop is about 120 m. The thickness of the lower member reaches about 88 m, but its base is not exposed. The lithotypes of this unit are composed of microcrystalline to dense, grey dolomites, which are characterised by stromatactis structures in the upper part. On top of this lithotype a strongly recrystallised, violet-brown to auburn carbonate is observed, which is characterised by high dolomitisation. A crimson bioclastic wackestone follows, sometimes associated with bioclastic floatstones. They are characteristic for the Barrios facies of the upper member of the Láncara Formation. Towards the top a continuous increase of siliciclastic material can be observed which is associated by an increase of millimetre- to centimetre-thick accumulations of insoluble material (pressure solution residuals). The top of the upper member of the Láncara Formation is represented by bioclastic wackestones and bioclastic floatstones. The transition to the overlying Oville shales is well exposed (Pl. 1.2–1.4). A zone of red to greenish claystones with grey-greenish limestone concretions and small limestone beds is located between the youngest Láncara beds and the greenish clay- and siltstones of the Oville Formation. 3.2. Los Barrios de Luna Section A section with a similar startigraphic and lithologic succession is exposed south of the village Los Barrios de Luna, at the storage lake termed Embalse de Luna in the southern part of the Somiedo-Correcilla unit (Fig. 1b). The classical Los Barrios de Luna section has been studied in different aspects since many years. Stratigraphic work was realised by Lotze (1961), van den Bosch (1969), Parga and Luque (1971), van Staalduinen (1973), and Martín Parra (1989); biostratigraphic work on trilobites was performed by Sdzuy (1961), on acritarchs by Palacios and Vidal (1992), and paleoichnological investigations were realised by Seilacher (1970) and Crimes et al. (1977). Recently, palaeosols were described from the section by Álvaro et al. (2003). The Los Barrios de Luna section is characterised by tectonical duplications. At the base of the section the siliciclastic Herrería Formation is exposed and overlain by the approximately 120 m thick succession of the lower member of the Láncara Formation. The thickness of the following. upper member is about 13 m. This part is characterised by carstification, possibly the result of its closeness to the surrounding fault systems. After the transition to the overlaying Oville Formation a re-occurrence of about 48 m thick sandstones of the Herrería Formation and about 102 m thick dolomites and dolomitic limestones of the lower member of the Láncara Formation is observed. This part in turn is overlain by about 113 m of the lower member of the Láncara Formation. The duplication is the result of a thrust fault. In the lower part of both succession shales are intercalated. The Los Barrios de Luna section consists of nearly the same lithotypes as described for the Genestosa section. The only difference is a thin unit (about 1.3 m thick) within the bioclastic wackestone of the Barrios facies that is characterised by a stromatactis-bearing bioclastic mudstone and a stromatactis-bearing bioclastic wackestone. It may correspond to the “pink limestone with stromatactis-like structures” described by Zamarreño (1972, p. 23). The stromatactis layer is characterised by a dramatic decrease in faunal content (Fig. 3) and a change in colour from crimson-brown to light-coloured nuances like red-ochre (described below). The section ends after about 15 m of carbonates of the upper member of the Láncara Formation. The boundary between the Láncara Formation and the Oville Formation is not exposed. 3.3. Vegaservera Section A section with an almost identical stratigraphic succession like at Genestosa and Los Barrios de Luna is exposed in the Vegaservera section (Fig. 1b), situated in the northern part of the Somiedo-Correcilla unit. It is exposed along the road between the villages Robles de la Valcueva in the south and Cármenes in the north. The profile is characterised by a discordant boundary between carbonates of the Carboniferous and the dolomites of the lower member of the Láncara Formation. The latter are characterised by diagenetic recrystallisation. Therefore no primary structures can be observed. As in the former sections, the base of the upper member of the Láncara Formation is characterised by bioclastic wackestones and bioclastic packstones (Beleño facies). They are overlain by a bioclastic packstone and a bioclastic wackestone, which represent the Barrios facies. The transition to the siliciclastics of the Oville Formation is not exposed. The section is formed by only 14 m of carbonates of the upper member of the Láncara Formation. The limestones of the upper member of the Láncara Formation are rich in a primarily micritic matrix, which is now diagenetically recrystallised into microsparite. Within the homogenous matrix only few abiogenous components (mainly clay minerals) are observed. They are diffusely dispersed or accumulated in residual layers caused by pressure solution. The carbonates of the Beleño facies are characterised by a comparably low amount of clay minerals so that structures.
(5) Facies distribution patterns and environment reconstruction of the Láncara Formation. 177. Pl. 1.1. Genestosa section; left: massive dolomites and dolomitic limestones of the lower member of the Láncara Formation, middle: nodular limestones of the upper member (overgrown by grass), right: siliciclastic sediments of the Oville Formation (followed by the Barrios Formation). Pl. 1.2. Typical outcrop-conditions of the Barrios facies. Well observable are the claystones of the overlying Oville Formation (upper part of the figure). Genestosa section. Folding rule (2 m) for scale. Pl. 1.3. Typical texture of the nodular limestone of the Barrios facies. Genestosa section. Camera lid for scale. Pl. 1.4. Transition zone between the upper member of the Láncara Formation and the Oville Formantion. Note the zone of red to greenish claystone with grey-greenish limestone concretions and small limestone beds located between the youngest Láncara beds and the greenish clay- and siltstones of the Oville Formation. Genestosa section. Folding rule (40 cm) for scale. Pl. 1.5. Bedding plains often show an intensive bioturbation. Genestosa section. Camera lid for scale. Pl. 1.6. Thin section of the Barrios facies. Note the intensive bioturbation and the high fossil content. Upper member of the Láncara Formation. Genestosa section (Ge 6). Scale bar: 1 mm. Pl. 1.7. Thin section of the Barrios facies. Well visible are the high fossil content and the well preserved fauna (mostly echinoderms, trilobites and brachiopods). Upper member of the Láncara Formation. Los Barrios de Luna section (BL b). Scale bar: 1 mm. Pl. 1.8. Thin section of the Beleño facies. Well visible is the high amount of echinoderms and glauconite. Lower member of the Láncara Formation. Los Barrios de Luna section (BL v/1). Scale bar: 1 mm..
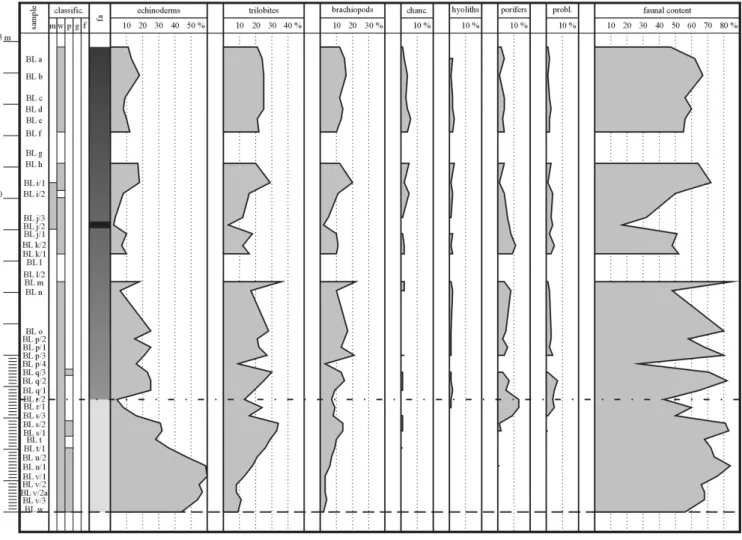
(6) 178. Thomas Wotte. of pressure solution occur less common, and limestone nodules could not be formed. Other abiogenetic components are pyrite crystals and small, subangular quartz-grains showing a diffuse dispersion or a nest-like enrichment. They are accumulated on sutures of pressure solution and residual-layers, however, their volume remains generally low (1% to 7%). The occurrence of local sparitic nests, which are observed in all of the investigated sections, is remarkable. Sometimes these areas are bound by shells or other faunal remains. These sparitic areas are caused by diagenetic coarsening of the primary cement. The most frequent lithotype of the upper member of the Láncara Formation is a moderately crystalline, reddish to crimson bioclastic wackestone with a high fossil content. Bioclastic packstones and fossil bearing mudstones are rarely observed. The top of the Barrios facies is mainly represented by bioclastic wackestones to bioclastic floatstones or only by bioclastic floatstones characterised by a lesser amount of (often fragmented) organismal remains. In the transitional zone to the Oville Formation, these lithotypes are characterised by a high amount of siliciclastic material, or they alternate with crimson to greenish claystones.. 4. BIOFACIES OF THE UPPER MEMBER OF THE LÁNCARA FORMATION In contrast to the underlying and overlying strata (dolomites and carbonates of the lower member of the Láncara Formation and the siliciclastics of the Oville Formation), the carbonates of the upper member of the Láncara Formation are characterised by a high fossil content (Pl. 1.6, 1.7). This content ranges between 48 and 87% at the base (Beleño facies) and between 17 and 89% in the Barrios facies (Figs. 3–5). The remarkable chaotic orientation of the fossils (especially within the Barrios facies) indicates an intensive bioturbation, which is well visible in thin sections and by burrows on the bedding plains (Pl. 1.5, 1.6). The vertical succession of biofacies is recorded by means of thin sections. The quantitative microfacial analysis of these thin sections indicates changes in the facies associations and allows to document the appearance and disappearance of faunal elements (Figs. 3–5; Tables 1–3). The most frequent biota of the upper member of the Láncara Formation are echinoderms, trilobites, and brachiopods. Chancellorids, sponges, molluscs (hyolithids and pelagiel-. Fig. 3. Faunal distribution patterns of the Upper Láncara Formation of the Los Barrios de Luna section; for legend see (Fig. 4)..
(7) Facies distribution patterns and environment reconstruction of the Láncara Formation. 179. Fig. 4. Faunal distribution patterns of the Upper Láncara Formation of the Vegaservera section.. lids), and palaeoscolecid sclerites are common, too. Echinoderms generally occur as disarticulated (although not abraded) ossicles. An abundance of plates, segments, and barrel-shaped elements were isolated by chemical preparation. Trilobites are generally detectable as isolated segments. However, complete organisms are also observed in thin sections and identified by their typical crook structures. The size of these fossils ranges from a few millimetres up to three centimetres. Some trilobite fragments could not be distinguished from brachiopods due to extensive breaking. Chemical preparation provides only disarticulated remains. Brachiopods occur (like trilobites) frequently, but usually as fragments. Complete valves are found primarily on bedding plains, mostly in curved-up position. The occurrence of different morphological forms is remarkable. A nearly smooth and spherical form (?Westonia) is situated only at the base of the upper member of the Láncara limestone, whereas the genuses Nisusia and Eoorthis are typical for the Barrios facies. During the chemical preparation a lot of microbrachiopods were found. They are represented by lingulids, acrotretids and paterinids. Acrotretid brachiopods. can be detected by the two genuses Vandalotreta and Acrothele. Lingulid and paterinid forms are represented by the genuses Lingulella and Micromitra, respectively. Chancellorid and sponge spicules are detectable in all of the studied sections. Many articulated sclerites were found in thin sections. Chemical preparation rendered mostly disarticulated remains of both groups. Molluscs are represented by helcionellids and hyolithomorph hyolithids. Specimens of both groups are visible in thin sections. Helcionellids are represented by the genuses of Helcionella, Latouchella, Pelagiella, and Protowenella, and were found in all of the sections during chemical preparation, whereas hyolithids were only observed in the Los Barrios de Luna section. Specimens believed to be palaeoscolecids are represented by hadimopellids. Van den Boogaard (1983) documented Hadimopanella for the first time from different sections of the Griotte limestone of the upper member of the Láncara Formation. Most samples yielded Hadimopanella oezgueli whose amounts varied from 14 to 12,000 specimens per 100 g of limestone (van den Boogaard, 1983). H. oezgueli also occurs in the studied section, but never in such mass.
(8) 180. Thomas Wotte. Fig. 5. Faunal distribution patterns of the Upper Láncara Formation of the Genestosa section. See Figure 4 for legend.. accumulations as reported by van den Boogaard (1983). The preservation of the organisms differs from sample to sample being corroded in some, but well preserved in others. The diameter of the individual sclerites varies from 100 to 200 µm. 5. BIOFACIES SUCCESSION AND PALEOECOCLOGICAL CONSIDERATIONS As described above, the preservation of the fossil remains varies considerably. Apart from a fragmentation of some brachiopod and trilobite remains, the generally good preservation of other faunal elements is remarkable (Pl. 1.6, 1.7). The fossils of the upper member of the Láncara Formation can thus be interpreted as representing a more or less autochthonous fauna with apparently little allochthonous input. Current activities, which can be held responsible for the transport are documented by the curved-up position of some brachiopods (Pl. 1.8). A clear vertical faunal differentiation is observable in the sections of Los Barrios de Luna and Vegaservera, lesser distinct in Genestosa (Figs. 3–5; Tables 1–3). In the first two. sections the base of the upper member of the Láncara Formation (developed in Beleño facies) is characterised by a high amount of echinoderms with trilobites and brachiopods of minor importance Within the Los Barrios de Luna section (Fig. 3; Table 2) the content of echinoderms amounts 44%, whereas trilobites and brachiopods are represented by 9% and 2%, respectively. The following part (about 1 m) shows an increase of echinoderms (up to 60% at BL v/1), whereas the number of trilobites and brachiopods does not change considerably. Towards the top a general increase of trilobites and brachiopods can be observed. The Los Barrios de Luna section is characterised by an increase of trilobites from 13 to 34% and of brachiopods from 3 to 14% (both values measured on sample BL v/1 and BL s/2). In contrast, the echinoderms show a decrease from 60 (BL v/1) to 31% (BL s/2). The following part of the Los Barrios de Luna section is characterised by a further decrease of echinoderms (up to 4% at sample BL r/2). An identical trend in the amount of trilobites and brachiopods should be emphasized even though the decline is not so drastic (minimum of trilobites 13% and brachiopods 7%). In contrast, the content of sponges shows an increase from 1 to 13%. After this fau-.
(9) Facies distribution patterns and environment reconstruction of the Láncara Formation. 181. Table 1-3. Values of point counting analysis. Every component (echinoderms, trilobites, brachiopods, chancellorids, hyoliths, poriferids, and problematic components) corresponding to a grid point was counted. “points” mean the number of counted points per thin-section. Point counting data of the Genestosa section points echinoderms trilobites brachiopods chancellorids hyoliths poriferids problematics faunal content Sample countedcounted % counted % counted % counted % counted % counted % counted % counted % Ge 23 363 34 9 71 20 58 16 1 0 1 0 24 7 2 1 191 53 Ge 22 358 40 11 64 18 57 16 0 0 1 0 26 7 4 1 192 54 Ge 21 363 46 13 76 21 60 17 2 1 2 1 27 7 2 1 215 59 Ge 20 287 41 14 50 17 40 14 1 0 1 0 25 9 6 2 164 57 Ge 19 sample not analysed Ge 18J 364 47 13 111 30 75 21 2 1 3 1 8 2 1 0 247 68 Ge 18K 357 45 13 104 29 74 21 2 1 3 1 9 3 6 2 243 68 Ge 18I 368 56 15 111 30 83 23 2 1 4 1 20 5 3 1 279 76 Ge 18H 328 35 11 59 18 55 17 0 0 1 0 18 5 2 1 170 52 Ge 18G 336 44 13 92 27 72 21 3 1 0 0 9 3 4 1 224 67 Ge 18F 362 61 17 105 29 92 25 5 1 1 0 4 1 4 1 272 75 Ge 18E 351 88 25 121 34 88 25 8 2 2 1 4 1 2 1 313 89 Ge 18D sample not analysed Ge 18C 321 36 11 89 28 93 29 2 1 1 0 9 3 2 1 232 72 Ge 18B 331 58 18 90 27 95 29 2 1 0 0 9 3 4 1 258 78 Ge 18A 342 59 17 87 25 88 26 1 0 2 1 4 1 4 1 245 72 Ge 17G 361 56 16 72 20 77 21 1 0 0 0 1 0 2 1 209 58 Ge 17F 366 56 15 94 26 98 27 0 0 1 0 1 0 3 1 253 69 Ge 17E 363 48 13 72 20 85 23 2 1 1 0 5 1 3 1 216 60 Ge 17D 339 56 17 78 23 77 23 2 1 2 1 1 0 1 0 217 64 Ge 17C 348 67 19 111 32 93 27 0 0 4 1 3 1 5 1 283 81 Ge 17B 365 72 20 90 25 92 25 0 0 3 1 3 1 2 1 262 72 Ge 17A 345 69 20 99 29 93 27 1 0 2 1 5 1 2 1 271 79 Ge 16H 254 49 19 60 24 57 22 2 1 4 2 5 2 3 1 180 71 Ge 16G 355 42 12 55 15 59 17 8 2 7 2 6 2 7 2 184 52 Ge 16F 365 71 19 100 27 105 29 3 1 4 1 4 1 3 1 290 79 Ge 16E 356 82 23 111 31 106 30 5 1 2 1 3 1 2 1 311 87 Ge 16D 317 61 19 64 20 60 19 21 7 9 3 1 0 10 3 226 71 Ge 16C 335 79 24 95 28 88 26 12 4 4 1 4 1 6 2 288 86 Ge 16B 348 87 25 92 26 76 22 10 3 2 1 2 1 4 1 273 78 Ge 16A sample not analysed Ge 15 343 84 24 96 28 83 24 0 0 1 0 0 0 1 0 265 77 Ge 14 346 56 16 81 23 88 25 1 0 0 0 2 1 3 1 231 67 Ge 13A 365 85 23 88 24 92 25 8 2 6 2 2 1 3 1 284 78 Ge 13 329 55 17 53 16 57 17 1 0 0 0 0 0 0 0 166 50 Ge 12 326 54 17 47 14 50 15 1 0 1 0 0 0 2 1 155 48 Ge 11 368 77 21 98 27 84 23 12 3 3 1 3 1 4 1 281 76 Ge 10K 368 90 24 92 25 100 27 10 3 1 0 6 2 9 2 308 84 Ge 10J 343 33 10 54 16 66 19 2 1 3 1 5 1 6 2 169 49 Ge 10I 318 53 17 79 25 95 30 0 0 2 1 7 2 9 3 245 77 Ge 10H 357 86 24 107 30 80 22 0 0 0 0 2 1 0 0 275 77 Ge 10G 346 88 25 99 29 104 30 1 0 3 1 3 1 6 2 304 88 Ge 10F 354 82 23 102 29 86 24 0 0 1 0 4 1 5 1 280 79 Ge 10E 352 81 23 94 27 80 23 3 1 2 1 2 1 0 0 262 74 Ge 10D 368 93 25 96 26 83 23 21 6 13 4 3 1 3 1 312 85 Ge 10C 342 80 23 106 31 71 21 17 5 5 1 3 1 2 1 284 83.
(10) 182. Thomas Wotte. Table 1-3. Continued. Point counting data of the Genestosa section points echinoderms trilobites brachiopods chancellorids hyoliths poriferids Sample countedcounted % counted % counted % counted % counted % counted % Ge 10B/1 368 78 21 98 27 81 22 17 5 8 2 5 1 Ge 10B 268 73 27 69 26 60 22 8 3 4 1 3 1 Ge 10A 362 89 25 101 28 101 28 5 1 3 1 6 2 Ge 9 Ge 8 301 49 16 61 20 33 11 3 1 2 1 0 0 Ge 7 321 33 10 60 19 56 17 0 0 1 0 2 1 Ge 6 341 90 26 98 29 38 11 8 2 7 2 3 1 Ge 5B 367 92 25 96 26 87 24 0 0 3 1 2 1 Point counting data of the Los Barrios de Luna section BL 347 37 11 72 21 42 12 2 1 1 0 7 2 BL a 315 41 13 77 24 46 15 6 2 6 2 12 4 BL b 356 65 18 89 25 58 16 13 4 5 1 4 1 BL c 351 30 9 86 25 43 12 10 3 7 2 14 4 BL d 359 29 8 89 25 52 14 14 4 8 2 14 4 BL e 357 34 10 74 21 45 13 20 6 11 3 9 3 BL f 321 38 12 70 22 32 10 13 4 6 2 12 4 BL g sample not analysed BL h 346 58 17 68 20 43 12 16 5 9 3 14 4 BL i/1 298 55 18 85 29 61 20 5 2 3 1 3 1 BL i/2 359 29 8 59 16 39 11 18 5 7 2 15 4 BL j/3 298 9 3 37 12 16 5 2 1 3 1 18 6 BL j/2 258 5 2 8 3 4 2 0 0 0 0 18 7 BL j/1 364 35 10 64 18 37 10 3 1 8 2 29 8 BL k/2 274 20 7 32 12 29 11 5 2 3 1 29 11 BL k/1 302 31 10 48 16 30 10 7 2 6 2 26 9 BL l sample not analysed BL l/2 sample not analysed BL m 368 65 18 131 36 82 22 8 2 5 1 16 4 BL n 324 21 6 56 17 33 10 7 2 5 2 26 8 BL o 300 76 25 83 28 51 17 1 0 4 1 16 5 BL p/2 359 55 15 74 21 55 15 1 0 3 1 10 3 BL p/1 306 76 25 66 22 40 13 0 0 2 1 18 6 BL p/3 292 60 21 80 27 62 21 5 2 3 1 13 4 BL p/4 341 53 16 29 9 10 3 0 0 0 0 0 0 BL q/3 296 69 23 88 30 38 13 2 1 2 1 8 3 BL q/2 263 66 25 66 25 40 15 3 1 3 1 19 7 BL q/1 205 51 25 38 19 18 9 2 1 5 2 10 5 BL r/2 335 14 4 43 13 23 7 0 0 4 1 44 13 BL r/1 280 23 8 66 24 26 9 0 0 3 1 35 13 BL s/3 311 50 16 51 16 24 8 2 1 0 0 28 9 BL s/2 302 94 31 103 34 41 14 4 1 0 0 2 1 BL s/1 318 101 32 106 33 44 14 1 0 1 0 6 2 BL t 320 90 28 93 29 32 10 2 1 0 0 1 0 BL t/1 296 107 36 77 26 27 9 1 0 1 0 0 0 BL u/2 293 138 47 61 21 20 7 1 0 0 0 1 0 BL u/1 292 171 59 53 18 18 6 0 0 0 0 3 1 BL v/1 307 183 60 39 13 10 3 0 0 0 0 0 0. problematics faunal content counted % counted % 3 1 290 79 2 1 219 82 5 1 310 86 0 0 0 148 49 2 1 154 48 5 1 249 73 1 0 281 77 2 6 4 7 9 8 5. 1 2 1 2 3 2 2. 163 194 238 197 215 201 176. 47 62 67 56 60 56 55. 12 4 14 9 10 8 13 9. 3 1 4 3 4 2 5 3. 220 216 181 94 45 184 131 157. 64 72 50 32 17 51 48 52. 5 7 9 11 5 11 0 3 18 10 15 14 2 1 4 0 0 0 0 0. 1 2 3 3 2 4 0 1 7 5 4 5 1 0 1 0 0 0 0 0. 312 155 240 209 207 234 92 210 215 134 143 167 157 245 263 218 213 221 245 232. 85 48 80 58 68 80 27 71 82 65 43 60 50 81 83 68 72 75 84 76.
(11) Facies distribution patterns and environment reconstruction of the Láncara Formation. Table 1-3. Continued. points echinoderms trilobites brachiopods chancellorids hyoliths poriferids Sample countedcounted % counted % counted % counted % counted % counted % Point counting data of the Los Barrios de Luna section BL v/2 236 130 55 20 8 6 3 0 0 0 0 0 0 BL v/2a 292 166 57 23 8 10 3 1 0 0 0 0 0 BL v/3 340 182 54 36 11 12 4 0 0 0 0 0 0 BL w 207 92 44 19 9 5 2 0 0 0 0 0 0 Point counting data of the Vegaservera section Ve 1 357 40 11 81 23 77 22 11 3 0 0 5 1 Ve 2 321 47 15 78 24 63 20 6 2 2 1 7 2 Ve 3 359 56 16 82 23 67 19 13 4 1 0 9 3 Ve 4 355 42 12 84 24 67 19 14 4 5 1 3 1 Ve 5 345 40 12 73 21 62 18 13 4 2 1 13 4 Ve 6 321 35 11 62 19 49 15 7 2 2 1 4 1 Ve 7 355 50 14 73 21 64 18 5 1 5 1 2 1 Ve 8 340 48 14 71 21 50 15 2 1 0 0 2 1 Ve 9 364 48 13 80 22 64 18 8 2 1 0 4 1 Ve 10 337 47 14 72 21 79 23 5 1 4 1 9 3 Ve 11 359 82 23 94 26 75 21 5 1 1 0 4 1 Ve 12 347 82 24 38 11 51 15 30 9 3 1 2 1 Ve 13 368 120 33 96 26 91 25 5 1 4 1 8 2 Ve 14 366 45 12 40 11 41 11 5 1 2 1 31 8 Ve 15 sample not analysed Ve 16 sample not analysed Ve 17 322 146 45 38 12 56 17 0 0 0 0 0 0 Ve 18 363 146 40 63 17 65 18 2 1 1 0 0 0 Ve 19 348 140 40 62 18 68 20 0 0 1 0 0 0 Ve 20 364 170 47 56 15 71 20 2 1 0 0 0 0 Ve 21 335 162 48 27 8 43 13 0 0 0 0 0 0 Ve 22 315 178 57 44 14 53 17 0 0 0 0 0 0 Ve 23 258 135 52 32 12 38 15 2 1 0 0 0 0 Ve 24 336 180 54 39 12 51 15 5 1 0 0 0 0 Ve 25 327 169 52 44 13 46 14 4 1 0 0 0 0. nal break the regular faunal succession could be observed. The content of echinoderms decreases further, whereas the content of trilobites and brachiopods fluctuates about a mean of 21% and 13%, respectively. Within the interval characterised by this assemblage stromatactis-bearing lithotypes (about 9 m of the upper member of the Láncara Formation) occur and are represented by the samples BL j/2 and BL j/3. These lithotypes are characterised by a low content of echinoderms (2 to 3%), trilobites (3 to 12%) and brachiopods (2 to 5%), whereas the content of chancellorids, hyoliths, poriferids, and problematic components is nearly unaffected. Within the Vegaservera section an almost identical faunal succession can be noticed (Fig. 4; Table 3). At the base of the upper member of the Láncara Formation echinoderms (52% at sample Ve 25) dominate, while trilobites and brachiopods are less common (13% trilobites and 14% brachi-. 183. problematics faunal content counted % counted % 0 0 0 0. 0 0 0 0. 156 200 230 116. 66 68 68 56. 2 5 3 6 11 3 4 6 2 5 3 2 4 7. 1 2 1 2 3 1 1 2 1 1 1 1 1 2. 216 208 231 221 214 162 203 179 207 221 264 208 328 171. 61 65 64 62 62 50 57 53 57 66 74 60 89 47. 0 0 0 1 0 0 0 2 0. 0 0 0 0 0 0 0 1 0. 240 277 271 300 232 275 207 277 263. 75 76 78 82 69 87 80 82 80. opods). Like at Los Barrios de Luna the following part is characterised by an increase in echinoderms (52% at sample Ve 22), whereas the content of trilobites and brachiopods is nearly constant. Towards the top an increase of trilobites and brachiopods is detected, whereas echinoderms decrease. Within the next 2 m detailed information is missing. A similar trend like at Los Barrios de Luna section can be assumed represented by a decline of echinoderms, trilobites and brachiopods. After this part a continuous decrease of echinoderms is observable (from 33% at Ve 13 to 11% at Ve 1). Approaching the top of the formation, the content of trilobites and brachiopods increases from 11 to 23% and from 11 to 22%, respectively (both values measured at sample Ve 14 and Ve 1). Section Genestosa represents an exceptional position, because neither a clear vertical faunal succession nor a con-.
(12) 184. Thomas Wotte. tinuous decrease of echinoderms nor an increase of trilobites and brachiopods was detected (Fig. 5; Table 1). Across the entire section the content of these fossil groups fluctuates about 19% (echinoderms), 25% (trilobites), and 22% for brachiopods. However, the top of the Genestosa section is dominated by trilobites (20%) and brachiopods (16%). Echinoderms are relatively rare (9%). In all the investigated sections the occurrence of chancellorids, sponges, and molluscs is generally low. However, specimens of these groups have been detected in nearly all samples. The detected fossil groups are divided into primary producers and consumers. The fauna at the base of the upper member of the Láncara Formation (Beleño facies) is dominated by sessile suspension feeders (echinoderms) acting as primary colonisers of the soft substrate. The amount of trilobites (deposit feeders) and filter feeders (brachiopods) is clearly lower. Towards the top of the Láncara Formation a gradual decrease of the sessile suspension feeders is observed, whereas filter feeders (brachiopods) and mobile benthic organisms (trilobites) increase in number. The successive change of the faunal content is accompanied by a continuous increase of siliciclastic material, which resulted in the formation of thick bands of insoluble material that were created by pressure solution processes, and in the colour change from roan to red and violet-brown. Taking into account all these considerations the faunal succession can be subdivided into several faunal assemblages (Figs. 3–5). The echinoderm-rich facies characterises the lowermost meters in the sections of Los Barrios de Luna and Vegaservera and is termed the Fauanl Assemblage A (Figs. 4 and 5) dominated by sessile suspension feeders. The decline of the echinodermal content (sessile suspension feeders) as well as the increase of the content of trilobites (mobile deposit feeders) and brachiopods (sessile filter feeders) marks the beginning of Fauanl Assemblage B (Figs. 4 and 5). This faunal association continues upward into the Fauanl Assemblage C (Figs. 3–5). Here the mobile benthos represented by trilobites dominates. Within this interval stromatactis-bearing lithotypes occur and are characterised by a distinct decrease in the faunal elements (Fauanl Assemblage D). Echinoderms, trilobites, and brachiopods decrease in number, whereas other faunal elements (chancellorids, hyolithids, sponge remains) do not show a marked change from Fauanl Assemblage C into Fauanl Assemblage D (Fig. 3). The Beleño facies (based on its definition) is equivalent to the Fauanl Assemblage A. Therefore the facies assemblages B, C, and D are components of the Barrios facies. 6. RECONSTRUCTION OF THE ENVIRONMENT AND DISCUSSION As described above the environment of the lower member of the Láncara Formation of the Somiedo-Correcilla. unit is interpreted as an eastward-tilting carbonate-dominated homoclinal ramp. The boundary between the dolomites and dolomitic limestone of the lower member of the Láncara Formation and the Faunal Assemblage A (Beleño facies) of the upper member is marked by a discontinuity termed “D2” by Álvaro et al. (2000). Towards the top a successive decrease of the echinodermal content, an increase in the content of trilobites and brachiopods, and also in siliciclastic content is detected. The continuous lithological and faunal succession, but overall low faunal and facial gradients of the upper member of the Láncara Formation are characteristic for a ramp environment. Therefore the development from the lower member to the upper member of the Láncara Formation is interpreted as a turnover from a homoclinal carbonate ramp to a mixed carbonatic-siliciclastic ramp (Beleño and Barrios facies, respectively Faunal Assemblages A, B, C, and D). The abrupt onset of the echinoderm-rich facies at the top of the dolostones points to an abrupt subsidence of the environment accompanied with a higher siliciclastic input (represented by slight residual layers caused by pressure solution). The high abundance of echinoderm ossicles indicates the existence of an echinodermal swell in the western part of the ramp. This swell supplied echinoderm debris into the distal environment and protected the distal environments from the siliciclastic influx as well. Therefore siliciclastic material reached the distal environment only in suspension. The short-termed abundance of echinoderms is eventually caused by the optimal adaptation to the deeper environmental conditions. The almost identical thickness of the Beleño and Barrios facies as well as the even biofacies distribution patterns within the studied sections point to a more or less constant rise in sea-level that caused a landward shift of facies belts, resulting in the same thickness of the Barrios facies. The long-term existence, the continuous increase of mobile deposit feeders as well as the decrease of sessil suspension feeders indicate a continuous and increasing siliciclastic input. The high siliciclastic content at the top of the Barrios facies as well as the clay- and siltstone layers at the transition to the Oville Formation point to the continuous drowning of the ramp. Álvaro et al. (2000) assumed a discontinuity (D3 after Álvaro et al., 2000) for the boundary between the Beleño and the Barrios facies that reflects a tectonic pulse accompanied with an input of siliciclastic material. During the examination all localities (about 20 in the whole Cantabrian zone) show a successive faunal and lithological change from the Beleño to the Barrios facies. Possibly this transition points to a widespread tectonic event, but the investigations indicate rather a continuous change in facies than a real discontinuity. Evidence for tectonically induced highs and lows, as postulated by Aramburu et al. (1992), Álvaro and Vennin (1996), and Álvaro et al. (2000), were also not observed. Assuming.
(13) Facies distribution patterns and environment reconstruction of the Láncara Formation. that tectonic activity yielded to a separation of the Láncara environment, siliciclastic dominated lithotypes (sediments of the grabens) should coexist with carbonates produced on the highs. Similar conditions were described for the “Amonitico rosso” by Cocozza and Gandin (1990). However, it was impossible to confirm this presumption despite an intensive sampling and a detailed microfacies analysis. Nevertheless, the environment of the Láncara Formation is characterised by a low morphology. Therefore some areas were influenced by higher or lower siliciclastic influx and also by some variations in the supply of echinodermal debris. The latter can be noticed within the Genestosa section. Here the upper member of the Láncara Formation is exclusively presented by the Barrios facies, whereas the Beleño facies disappears. This observation points to a position of this locality on a morphological high. In contrast to the Los Barrios de Luna and Vegaservera section this position was nearly unaffected by the supply of echinodermal debris. 7. CONCLUSION The environment of the Lower–Middle Cambrian Láncara Formation in the Somiedo-Correcilla unit is interpreted as a turnover from an eastward-tilting homoclinal carbonate ramp (lower member) to a mixed carbonatic-siliciclastic ramp (upper member) displaying low topographic variations. The abrupt onset of the Beleño facies (Faunal Assemblage A) points to a sudden subsidence of the environment. A continuous and increasing siliciclastic influx, and an accompanied change in the faunal content during the deposition of the Beleño and Barrios facies indicates a successive drowning of the ramp This development is mirrored by the Faunal Assemblages B-D. ACKNOWLEDGEMENTS: The author is grateful to Dr. O. Elicki (Freiberg, Germany) for the participation during field work and for many helpful discussions. Warmest thanks are also due to Prof. G. Geyer (Würzburg, Germany) and Prof. J. J. Álvaro (Lille, France) for constructive comments and criticism of the manuscript. Special thanks for linguistic help to K. Kuhnt (Paris, France). This study is element of the project “The microfauna of the Early to Middle Cambrian Láncara Formation (Cantabrian Mountains, NW-Spain): micropalaeontology, palaeobiogeography, and process dynamics at the Early–Middle Cambrian transition of western Gondwana” supported by the German Research Foundation (EL 144/12).. REFERENCES Álvaro, J.J. and Vennin, E., 1996, Tectonic control on Cambrian sedimentation in South-Western Europe. Ecologae Geologicae Helvetiae, 89, 935–948. Álvaro, J.J. and Vennin, E., 1997, Episodic development of Cambrian eocrinoid-sponge meadows in the Iberian Chains (NE Spain). Facies, 37, 49–64. Álvaro, J.J., Vizcaïno, D. and Vennin, E., 1999, Trilobite diversity patterns in the Middle Cambrian of southwestern Europe: a com-. 185. parative study. Palaeogeography, Palaeoclimatology, Palaeoecology, 151, 241–254. Álvaro, J.J., Vennin, E., Moreno-Eiris, E., Perejón, A. and Bechstädt, T., 2000, Sedimentary patterns across the Lower-Middle Cambrian transition in the Esla nappe (Cantabrian Mountains, northern Spain). Sedimentary Geology, 137, 43–61. Álvaro, J.J., Van Vliet-Lanoë, B., Vennin, E. and Blanc-Valleron, M.M., 2003, Lower Cambrian paleosols from the Cantabrian Mountains (northern Spain): a comparison with Neogene-Quarternary estuarine analogues. Sedimentary Geology, 163, 67–84. Aramburu, C. and García-Ramos, J.C., 1993, La sedimentación cambro-ordovícica en la Zona Cantábrica (NO de España). Trabajos de Geología, 19, 45–73. Aramburu, C., Truyols, J., Arbizu, M., Méndez-Bedia, I., Zamarreño, I., García-Ramos, J.C., Suarez de Centi, C. and Valenzuela, 1992, El Paleozoico Inferior de la Zona Cantábrica. In: Gutiérrez Marco, J.C., Saavedra, J. and Rábano, I. (eds.), Paleozoico Inferior de Ibero-América. Madrid, p. 397–421. Bechstädt, T. and Boni, M., 1989, Tectonic control on the formation of a carbonate platform: the Cambrian of southwestern Sardinia. The Society of Economic Paleontologists and Mineralogists, Special Publication, 44, 107–122. Bechstädt, T. and Boni, M., eds., 1994, Sedimentological, stratigraphical and ore deposits field guide of the autochthonous Cambro-Ordovician of southwestern Sardinia: Memorie descrittive della carta geologica d’Italia, vol. XLVIII. Servizio Geologico Nazionale, 434 p. Bechstädt, T., Schledding, T. and Selg, M., 1988, Rise and fall of an isolated, unstable carbonate platform: the Cambrian of Southwestern Sardinia. Geologische Rundschau, 77, 389–416. Cocozza, T. and Gandin, A., 1990, Carbonate deposition during early rifting: the Cambrian of Sardinia and the Triassic-Jurassic of Tuscany, Italy. Special Publication of the International Association of Sedimentologists, 9, 9–37. Crimes, T.P., Legg, I., Marcos, A. and Arboleya, M.L., 1977, Late Precambrian–low Lower Cambrian trace fossils from Spain. In: Crimes, T.P. and Harper, J.E. (eds.), Trace Fossils. Geological Journal, Special Publication, 9, p. 91–138. Fernández Remolar, D.C., 2001, Chancelloridae del Ovetiense Inferior de la Sierra de Córdoba, España. Revista Española de Paleontología, 16, 39–62. Geyer, G., 1986, Mittelkambrische Mollusken aus Marokko und Spanien. Senckenbergiana lethaea, 67, 55–118. Julivert, M., 1971, Décollement tectonics in the Hercynian Cordillera of NW Spain. American Journal of Science, 270, 1–29. Lotze, F., 1945, Zur Gliederung der Varisciden der Iberischen Meseta. Geotektonische Forschungen, 6, 63–81. Liñán, E., Gozalo, R., Palacios, T., Gámez Vintaned, J.A., Ugidos, J.M. and Mayoral, E., 2002, Cambrian. In: Gibbson, W. and Moreno, T. (eds.), The Geology of Spain. Geological Society, London, p. 17–31. Lotze, F., 1961, Das Kambrium Spaniens. Teil I: Stratigraphie. Akademie der Wissenschaften und der Literatur, Mainz, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse, 1–216. Martín Parra, L.M., 1989, Mapa Geológico de España, 2a serie, no. 128 (Riello) (1:50,000). ITGE Press., Madrid, p. 1–119. Palacios, T. and Vidal, G., 1992, Lower Cambrian acritarchs from Northern Spain: the Precambrian-Cambrian boundary and biostratigraphic implications. Geological Magazine, 129, 421–436. Parga, J.R. and Luque, C., 1971, Las series del Cámbrico y Eocámbrico en la Cordillera Cantábrica. Boletin Geologico y Minero,.
(14) 186. Thomas Wotte. 82, 310–320. Pérez-Estaún, A., Bastida, F., Alonso, J.L., Marquínez, J., Aller, J., Alvarez-Marrón, J., Marcos, A. and Pulgar, J.A., 1988, A thinskinned tectonics model for an arcuate fold and thrust belt: the Cantabrian Zone (Variscan Ibero-Armorican Arc). Tectonics, 7, 517–537. Rodrígues Fernández, L.R., Barba, P., Bardají, T., Silva, P.G., Suárez Rodríguez, P.I., Heredia, N., Gallastegui, G., Paniagua, A. and Galán, L., 1991, Mapa Geológico de España, 2a serie, no. 102 (Los Barrios de Luna) (1:50,000). ITGE Press., Madrid, p. 1–130. Rupke, J., 1965, The Esla nappe, Cantabrian Mountains (Spain). Leidse Geologische Mededelingen, 32, 1–74. Russo, A. and Bechstädt, T., 1994, Evolución sedimentológica y paleogeográfica de la Formación Vegadeo (Cámbrico inferior– medio) en la zona entre Visuña y Piedrafita do Caurel (Lugo, NO de España). Revista de la Sociedad Geológica de España, 7, 299– 310. Sdzuy, K., 1961, Das Kambrium Spaniens. Teil III: Trilobiten. Akademie der Wissenschaften und der Literatur, Abhandlungen der mathematisch-naturwissenschaftlichen Klasse, p. 217–408. Sdzuy, K., 1969, Unter- und mittelkambrische Porifera (Chancelloriida und Hexactinellida). Paläontologische Zeitschrift, 43, 115–147. Sdzuy, K., 1974, Mittelkambrische Graptolithen aus NW-Spanien. Paläontologische Zeitschrift, 48, 110–139. Seilacher, A., 1970, Cruziana stratigraphy of ‘non fossiliferous’ Palaeozoic sandstones. In: Crimes, T.P. and Harper, J.E. (eds.), Trace Fossils. Geological Journal, Special Publication, 3, p. 444–476. Truyols, J., Arbizu, M.A., García Alcalde, J.L., García López, S., Méndez Bedia, I., Soto, F. and Truyols Massoni, M., 1990, Cantabrian and Palentian Zones; Stratigraphy: The Asturian-Leonese Domain (Cantabrian Zone). In: Dallmeyer, R.D. and Martínez García, E. (eds.), Pre-Mesozoic Geology of Iberia. Springer Ver-. lag, Berlin, p. 10–19. Van den Bosch, W.J., 1969, Geology of the Luna-Sill region, Cantabrian Mountains (NW Spain). Leidse Geologische Mededelingen, 44, 137–225. Van den Boogaard, M., 1983, The occurrence of Hadimopanella oezgueli Gedik in the Láncara Formation in NW Spain. Palaeontology, 86, 331–341. Van der Meer Mohr, C.G., 1983, The Lower Palaeozoic of the Leonides and Pisuerga-Carrión Province in the Cantabrian zone of northwest Spain. ITC Journal, 3, 205–213. Van Staalduinen, C.J., 1973, Geology of the area between the Luna and Torío rivers, southern Cantabrian Mountains, NW Spain. Leidse Geologische Mededelingen, 49, 167–205. Vidal, G., Palacios, T., Moczydowska, M. and Gubanov, A.P., 1999, Age constraints from small shelly fossils on the early Cambrian terminal Cadomian Phase in Iberia. Geologiska Föreningens Forhandlingar, 121, 137–143. Wotte, Th., Elicki, O., Perejón, A. and Moreno-Eiris, E., 2004, Facies distribution patterns and environment interpretation of the Upper Láncara Formation in the Esla Nappe (Cantabrian zone, NW Spain) by quantitative microfacies analysis. Freiberger Forschungshefte, C 502, Paläontologie, Stratigraphie, Fazies, 12, 101–118. Zamarreño, I., 1972, Las litofacies carbonatadas del Cámbrico de la Zona Cantábrica (NW de España) y su distribución paleogeográfica. Trabajos de Geología, 5, 1–118. Zamarreño, I., 1975, Peritidal origin of Cambrian carbonates in Northwest Spain. In: Ginsburg, R.N. (ed.), Tidal Deposits: a Case Book of Recent Examples and Fossil Counterparts. Springer Verlag, Berlin, p. 323–332. Manuscript received September 21, 2004 Manuscript accepted April 18, 2005.
(15)
수치

관련 문서
Modern Physics for Scientists and Engineers International Edition,
“The Chinese Values and The Search for Culture- Free Dimensions of Culture: The Chinese Culture Connection,” Journal of Cross-Cultural Psychology, 18(2), June,
If both these adjustments are considered, the resulting approach is called a bootstrap-BC a -method (bias- corrected-accelerated). A description of this approach
③ A student who attended Korean course at KNU Korean Language Program and holds TOPIK Level 3 or a student who completed Korean course Level 4 at the KNU Korean Language
· 50% exemption from tuition fee Ⅱ for the student with a TOPIK score of level 3 or higher or completion of level 4 or higher class of the Korean language program at the
Department of Internal Medicine, Novosibirsk State University, Russia 1 , Department of Cardiology, Surgut State University, Russia 2 , Department Fundamental
웹 표준을 지원하는 플랫폼에서 큰 수정없이 실행 가능함 패키징을 통해 다양한 기기를 위한 앱을 작성할 수 있음 네이티브 앱과
_____ culture appears to be attractive (도시의) to the
