527
Copyright © 2014 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
체내의염증반응은미생물감염
,
내독소,
조직손상과같은위 해성자극에대한방어기능으로,
이는조직의구조와기능을정 상적으로회복하기위해필수적으로일어나는반응이다.
정상 적인염증반응은시간이지남에따라염증촉진성매개체(pro- inflammatory mediators)
의생성은감소되고,
항염증성 매개 체(anti-inflammatory mediators)
는 증가됨으로써 스스로 염 증반응이제한되는조절과정을가지고있다(Lawrence et al.,
2002).
체내의염증반응에관여하는세포중하나인대식세포(macrophages)
는 이러한염증반응에중요한역할을하고있 다.
대식세포는interferon-γ (IFN-γ), interleukin (IL)-1β, IL-6, tumor necrosis factor-α (TNF-α)
와같은염증촉진성사이토카 인(cytokines),
그리고세균세포막성분인lipopolysaccharides
(LPS)
등의자극에노출됨으로써활성화된다(Xie et al., 1993).
활성화된대식세포는염증촉진성사이토카인이외에
inducible nitric oxide synthase (iNOS)
및cyclooxygenase-2 (COX-2)
와 같은효소의발현을통해nitric oxide (NO)
및prostaglandin E
2(PGE
2)
와같은다양한염증매개분자들을생성하게되고(Na- than, 1992; Zhang and Ghosh, 2000),
이들매개체들의과도한 생성은다양한만성염증성질환의발병에기여하고있는것으 로알려져있다.
따라서염증성매개체들의생성을억제하는천 연화합물의발견은보다부작용이적은항염증치료제개발을 위한방편으로관심을끌고있다.
대식세포에있어염증촉진성 사이토카인및 단백질들의발 현은
nuclear factor kappa-B (NF-κB)
에 의해전사수준에서 조절된다(Pahan et al., 2001).
자극이없는상태에서NF-κB
는inhibitor of kappa B (IκB)
와결합한상태로불활성형태로LPS로 유도된 RAW 264.7 대식세포에 대한 애기외톨개 모자반 (Myagropsis yendoi) 에틸아세테이트 분획물의 항염증 효과
김보운·김재일·김형락·변대석*
부경대학교 식품영양학과
Anti-inflammatory Effect of an Ethyl Acetate Fraction from Myagropsis yendoi on Lipopolysaccharides-stimulated RAW 264.7 Cells
Bowoon Kim, Jae-Il Kim, Hyeung-Rak Kim and Dae-Seok Byun*
Department of Food Science and Nutrition, Pukyong National University, Busan 608-737, Korea
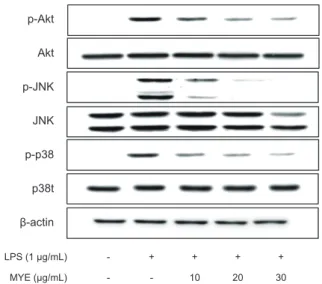
An ethanolic extract from Myagropsis yendoi was fractionated using several solvents. Among these, an ethyl acetate fraction ( Myagropsis yendoi ethyl acetate fraction: MYE) showed the highest anti-inflammatory activity based on inhibition of lipopolysaccharides (LPS)-induced nitric oxide (NO) production in RAW 264.7 cells. We thus investi- gated the molecular mechanisms underlying MYE’s inhibitory effects. Pretreatment of cells with up to 30 µg/mL of MYE significantly inhibited NO production and inducible nitric oxide synthase expression in a dose-dependent man- ner ( P <0.05). Similarly, MYE markedly reduced the production of pro-inflammatory cytokines, such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α, as well as their mRNA levels. While the nuclear translocation of nuclear factor-kappa B (NF-κB) was strongly suppressed by MYE, the activation of a nuclear factor erythroid 2-re- lated factor (Nrf2) was increased. Moreover, MYE significantly reduced the phosphorylation of JNK, p38 MAPK, and phosphatidylinositol 3-kinase/Akt in LPS-stimulated cells. These results indicate that MYE contains anti-inflam- matory compounds, and that it might be used as a dietary supplement for the prevention of inflammatory diseases.
Key words: Myagropsis yendoi , Anti-inflammatory effect, Pro-inflammatory cytokines, iNOS, NF-κB
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/)which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
http://dx.doi.org/10.5657/KFAS.2014.0527 Kor J Fish Aquat Sci 47(5) 527-536, October 2014 Received 19 September 2014; Accepted 29 September 2014
*Corresponding author: Tel: +82. 51. 629. 5844 Fax: +82. 51. 629. 5842
E-mail address: [email protected]
김보운
ㆍ
김재일ㆍ
김형락ㆍ
변대석528
세포질에존재한다
(D'Acquisto et al., 1997; Makarov, 2001).
그러나
LPS
와같은자극이주어지는경우IκB
는IκB kinase (Ikk)
에의해 인산화되어proteasome
에의해분해되고,
유리된
NF-κB
는핵으로이동하여다양한염증성매개체와 같은표적유전자의 발현을 유도하게 된다
(Chen et al., 1995).
한 편NF-κB
의 활성화는extracellular signal-regulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38 kinase
를포함하 는mitogen-activated protein kinases (MAPKs)
그리고PI3K/
Akt (PKB)
와같은kinase
에의해조절되는것으로알려져있다(Marks-Konczalik et al., 1998; Jang et al., 2005).
Nuclear factor erythroid 2–related factor 2 (Nrf2)
는산화적 스트레스에대응하는세포방어기전을위한일차적인전사인 자로알려져있다(Chen and Kunsch, 2004).
정상적인상태에 서Nrf2
는Keap1
에의해비활성화상태로세포질에존재하지 만,
산화적손상을받으면Keap1
과해리되어핵내로이동하여antioxidant response element (ARE)
에결합함으로써HO-1
과 같은항산화효소의발현을조절한다(Nguyen et al., 2000).
최근의연구에의하면
, Nrf2
는패혈증을유발시킨생쥐의모델실험에서세포보호효과를나타내었으며
,
특히Nrf2 knock-out
생 쥐는LPS
로유도된염증반응에과도하게반응하는것으로보 고되고있다(Ha et al., 2011). Nrf2
활성화에의해생성된HO-1
은RAW 264.7
세포에서LPS
에의해생성된염증촉진성사이 토카인의생성을억제함으로써항염증효과를나타낸다고보고 되고있다(Lee et al., 2014).
연안에널리자생하는해조류는비타민과무기질이다량함 유되어있으며식이섬유가풍부하다
.
현재많은연구들이해조 류의항균효과,
항진균효과,
그리고항바이러스활성을가진다 는것을밝혔으며(Del et al., 2001),
고지혈증억제(Awad et al., 2003)
및항암(Lee et al., 1992)
과같은생리활성을가진다는결 과들이발표되었다.
그중갈조류는저분자생리활성물질로써fucoxanthin, fucosterol, phlorotannin
과같은다양한생리활성 물질들을가지고있는것으로보고되고있다(Kim et al., 2010b;
Kim et al., 2013a; Lee et al., 2013; Jung et al., 2014).
특히모 자반류는항염증효과를가지고있는것으로나타났고(Joung et al., 2012a; Joung et al., 2012b; Gwon et al., 2013; Kim et al., 2013b), phlorotannin, sargachromenol
및sargaquinoic acid
과 같은다양한화합물들이함유되어있는것으로보고되고있다(Kang et al., 2013; Kim et al., 2014).
다양한 해조류들 주에서 애기외톨개모자반
(Myagropsis
yendoi)
은일본과우리나라동남해연안에서식하는 모자반과
(Sargassaceae)
의개모자반속(Myagropsis)
에속하는갈조류 로(NFRDI., 2009),
저조선부근에생육하며다년생해조류로봄에성숙하고뿌리를포함한개체가섬세하고유연하다
(Dae-
kyo, 2008).
식용이가능한것으로알려져있는애기외톨개모 자반은이전연구에서뛰어난항균활성이밝혀진바있으며(Park et al., 2010),
최근에는이외의생리활성에관한연구들이활발히진행중이다
.
최근본연구실에서수행한예비실험에 서애기외톨개모자반의에탄올(95%, v/v)
추출물로부터유기 용매(hexane, ethyl acetate, butanol)
를이용하여각분획물들 을분리하였고그활성을분석한결과, EtOAc
분획물에서항 염증효과가강하게나타남을확인하였다.
따라서본연구에서 는이러한애기외톨개모자반의EtOAc
분획물(M. yendoi ethyl acetate extract, MYE)
의항염증효과및관련분자적기전을LPS
로자극한RAW 264.7 cell
을이용하여분석함으로써,
염 증성질환의발병을예방또는지연시킬수있는건강기능성소 재로서의이용가능성을검토하였다.
재료 및 방법
추출과 분획
부산기장에서채취한
M. yendoi
의건조분말2 kg
을환류냉 각기가부착된집기병에담고95%
주정(EtOH, v/v) 4 L
를넣어 가열,
추출하고(50°C, 3
시간)
추출액을여과하여회전진공농축 기를사용하여농축하였다.
이를3
회반복하여총182 g
의주정 추출물을얻었다.
이를H
2O:EtOH (9:1, v/v)
의혼합용매로녹 인후동량의n-
헥산을넣어분액깔대기에평형화시켜상층액 의n-
헥산가용부를분리하였다.
동일한방법으로수용층에동 량의에틸아세테이트(ethyl acetate, EtOAc)
를넣어평형화한 다음EtOAc
획분을분리하여농축하여MYE
를얻었다. 세포 배양 및 처리
RAW 264.7
세포(ATCC, Rockville, MD, USA)
는10%
fetal bovine serum (FBS)
와penicillin (100 units/mL), strep- tomycin sulfate (100 µg/mL)
을첨가한Dulbecco’s modified Eagle’s medium (DMEM)
을사용하였고, 5% CO
2, 37°C
배 양기에서배양하였다. Cell culture plate
에RAW 264.7 cell
이70~80%
정도채워지면phosphate-buffered saline (PBS)
로한 번씻어낸후,
계대배양하였다. MYE
는100% dimethyl sulf- oxide (DMSO)
에녹여사용하였고,
배양세포처리전에배지에 희석하여처리하였다.
세포 독성 시험
RAW 264.7 cell
을96-well plate
에5×10
5cells/well
로분주 하고37°C
에서24
시간동안배양한후, MYE
가0, 10, 20, 30
µg/mL
농도로희석된DMEM
배지로교체하여1
시간배양 하였고,
이후에LPS (1 µg/mL)
를함유한DMEM
배지에다 시24
시간배양하였다.
이후CellTiter96
ⓇAqueous 3-(4,5-di-
methylthiazol-2-yl)-5-(3-carboxymethoxyphenyl)-2-(4-sul-
fophenyl)-2H-tetrazolium (MTS)
시험키트(Promega, Madi-
son, WI, USA)
를 사용하여 제조사의방법에따라 세포생존 율을측정하였다. MTS
용액은FBS-free DMEM
에5% (v/v)
의농도로섞어100 µL
씩처리하였다. 1
시간후에microplate
reader (Glomax Multi Detection System, Promega, Madison, WI, USA)
를이용하여490 nm
의파장에서흡광도를측정하여 세포독성을분석하였다.
NO 및 염증성 사이토카인 생성 억제 효과
MYE
의 항염증효과 비교를 위해RAW 264.7
세포에서의NO
생성에대한억제효과를분석하였다. MYE
를1
시간동안 전처리한다음LPS (1 µg/mL)
로24
시간동안자극하고,
그배 지를원심분리(2,000 g, 4°C, 10
분)
하여회수하였다. NO
의농 도는배지(100 µL)
와Griess
시약(0.1% naphthylethylene di- amine dihydrochloride + 1% sulfanilamide + 5% phosphoric acid)
을동일한비율로반응시켜microplate reader
로540 nm
의파장에서흡광도를측정하였다(Kim et al., 2009).
배지중 의IL-1β, IL-6, TNF-α
의양은enzyme-linked immunosorbent assay kit (ELISA, R&D Systems, Minneapolis, MN, USA)
를 이용하여제조사의방법에따라측정하였다.
mRNA 발현양의 분석
RAW 264.7
세포(1×10
6cells/well)
를0, 10, 20, 30 µg/mL
의농도로MYE
를1
시간동안처리한후, LPS 1 µg/mL
의농 도로6
시간동안자극시켰다.
이후Quiazol
시약(Quiagen Sci- ence, Valencia, CA, USA)
을이용하여total RNA
를분리하였 다(Kim et al., 2009). Total RNA
로부터역전사중합효소연 쇄반응(RT-PCR)
분석에의한mRNA
발현양의 분석은이전 의보고(Kim et al., 2009)
에서사용한방법을이용하였고,
유 전자발현량의상대적인비교를위해서housekeeping gene
인glyceraldehye-3-phosphate dehydrogenase (GAPDH)
를함께 분석하였다. PCR
반응에이용된각각의primer
는Table 1
에나 타내었다.
전기영동상DNA
밴드의정량분석은cooled CCD
camera system EZ-Capture Ⅱ (ATTO & Rise Co., Tokyo, Ja- pan)
과CS analyzer ver. 3.00 software (ATTO)
를이용하여최 소3
번의반복실험을통해얻었다.
세포질 및 핵 단백질 추출물의 제조
NF-κB
의활성화정도를분석하기위해MYE
및LPS
를각 각처리한RAW 264.7
세포로부터세포질및핵단백질추출 물을각각분리제조하였다(Kim et al., 2009).
즉, MYE
로처 리된 세포(2×10
6cells/dish)
를PBS
로 세척하여 회수하고, 180 µL
의hypotonic buffer [10 mM Tris-HCl, 10 mM NaCl, 3 mM MgCl
2, 0.02% NaN
3, 0.5 mM dithiothreitol (DTT), 1mM phenylmethanesulfonyl fluoride (PMSF), pH 7.4]
를넣 고, 20 µL
의5% nonidet NP-40
을첨가하여5
분동안반응시 켰다.
이후원심분리(1,800 g, 4°C, 5
분)
한후상층액을세포질 추출물로이용하였다.
침전물은hypotonic buffer
로한번세척 하고, hypertonic buffer [20 mM 4-(2-hydroxyethyl)-1-piper- azineethanesulfonic acid, 25% glycerol, 420 mM NaCl, 1.5 mM MgCl
2, 0.2 mM ethylenediaminetetraacetic acid, 0.02%
NaN
3, 0.5 mM DTT, 1 mM PMSF, pH 7.4]
를넣고1
시간동안 얼음위에방치시킨다음원심분리(13,000 g, 4°C, 10
분)
하여상 층액을회수하여핵단백질추출물로이용하였다.
Western blot 분석에 의한 단백질 분석
염증관련 단백질과신호전단 단백질의인산화정도는세포 를
MYE
및LPS
로처리한이후whole cell lysate
를제조하여 시료로, NF-κB
및IκB
의활성화및인산화정도는상기의핵 및세포질추출물을시료로이용하였고,
단백질의양은이전의 보고(Kim et al., 2009)
와마찬가지로sodium dodecyl sulfate- polyacrylamide gel electrophoresis (SDS-PAGE)
분리된 단 백질을nitrocellulose membrane
에 이전시켜Western blot
으 로분석하였다.
검출된밴드의정량분석은mRNA
분석과마 찬가지로cooled CCD camera system EZ-Capture Ⅱ
와CS analyzer ver. 3.00 software
를이용하여최소3
번의반복실험 을통해얻었고,
그결과를각blot
의하단에수치로표기하였 다.
그리고, Western blot
에사용된각각의1
차항체들은다음 과같다: iNOS (sc-650), β-actin (sc-47778), phospho-Akt (sc- 4060), Akt (sc-1618), phospho-ERK (sc-7883), ERK (sc-94), phospho-JNK (sc-6254), JNK (sc-7345), NF-κB/p65 subunit (sc-8008), HO-1 (sc-7696), Nrf2 (sc-722), poly (ADP-ribose) polymerase (PARP, sc-7150)
는Santa Cruz Biotechnology (Santa Cruz, CA, USA)
에서 구입하였고, phospho-IκB-α (4814), IκB-α (9246), phospho-p38 (4511), p38 (9212)
는Cell Signaling Technology (Danvers, MD, USA)
에서각각구입하 였다. Horseradish peroxidase (HRP)
가conjugate
되어있는각 각의2
차항체들[rabbit anti-goat IgG (LF-SA5004), goat anti- mouse IgG (LF-SA5001), goat anti-rabbit IgG (LF-SA5002)]
은
AbFrontier (Seoul, Korea)
에서 구입하였고, Enhanced
Table 1. Primer sequences used in this studyPrimers Sequences
iNOS Forward 5'-TCT TTG ACG CTC GGA ACT GT-3‘
Reverse 5'-CCA TGA TGG TCA CAT TCT GC-3‘
COX-2 Forward 5'-TGG GCA AAG AAT GCA AAC AT-3‘
Reverse 5'-CAG CAA ATC CTT GCT GTT CC-3’
HO-1 Froward 5'-AAG ATT GCC CAG AAA GCC CTG GAC-3' Reverse 5'-AAC TGT CGC CAC CAG AAA GCT GAG-3‘
TNF-α Forward 5'-CAA GGG ACA AGG CTG CCC CG-3' Reverse 5'-GGT CAG AGT GGG GGC TGG GT-3‘
IL-1β Forward 5'-ATG GCA ACT GTT CCT GAA CTC AAC T-3' Reverse 5'-CAG GAC AGG TAT AGA TTC TTT CCT TT-3' IL-6 Forward 5'-GTA TGA ACA ACG ATG ATG CAC TTC CAG-3' Reverse 5'-GCA TTG GAA ATT GGG TAG GAA GG-3' β-actin Forward 5'-CCT CAT GAA GAT CCT GAC CG-3'
Reverse 5'-TCC ACA TCT GCT GGA AGG TG-3'
김보운
ㆍ
김재일ㆍ
김형락ㆍ
변대석530
chemiluminescence (ECL) detection kit
은GE Healthcare Bio-Science (Piscataway, NJ, USA)
를사용하였다. β-actin
과PARP
는각각세포질과핵의control
단백질로서분석에포함 시켰다.
면역형광분석법
RAW 264.7
세포를glass coverslips (SPL Lifesciences Co., Gyeonggi-do, Korea)
위에24
시간배양한뒤, MYE
로1
시간 전처리하고, LPS (1 µg/mL)
로30
분자극시켰다.
세포를4.0%
paraformaldehyde
가첨가된PBS
로실온에서15
분동안반응 시켜고정시키고, 0.5% Triton X-100
이첨가된PBS
를넣어10
분동안반응시켰다. PBS
로세척한뒤에3% BSA/PBS
를넣고30
분동안blocking
시킨후, anti-NF-κB polyclonal antibody
가 희석된3% BSA/PBS
를넣어2
시간동안반응시켰다.
그다음, Alexa Fluor
Ⓡ488-conjugated secondary antibody (Invitrogen, Carlsbad, CA, USA)
가희석된3% BSA/PBS
를넣고1
시간 동안반응시킨뒤, 2 µg/mL
의4,6-diamidino-2-phenylindole (DAPI)
로핵을염색하고LSM700 laser scanning confocal mi- croscope (Carl Zeiss, Oberkochen, Germany)
로관찰하였다. NF-κB Promoter/Luciferase assay
RAW 264.7
세포(2×10
5cells/well)
가 들어있는24-well plate
의각well
에1 µg
의pNF-κB firefly luciferase DNA
와20 ng
의pRL-TK renilla luciferase DNA
를lipofectamine/plus reagent (Invitrogen, Carlsbad, CA, USA)
와함께처리하여40
시간동안transfection
시켰다.
그다음, MYE
를1
시간전처리 하고, LPS (1 µg/mL)
로6
시간자극시켰다.
이후PBS
로세척 하고100 µL
의lysis buffer (0.5 mM HEPES, pH 7.8, 1% Tri- ton N-101, 1 mM CaCl
2, and 1 mM MgCl
2)
로용해물을만들 고, luciferase assay kit
를사용하여firefly luciferase activity
와renilla luciferase
활성을측정하였다. Renilla luciferase
의발현 은지속적으로일어나는반면, firefly luciferase
는NF-κB
에의 해서만발현이되므로세포수에의한오차를보정할수있다. 통계 처리
본연구의모든실험은세번이상반복하였으며
,
얻어진결과들을평균값과표준편차
(mean±SD)
를계산하여나타내었다
.
실험군간의유의성검증은Student’s t-test
로검증하였다.
결과 및 고찰
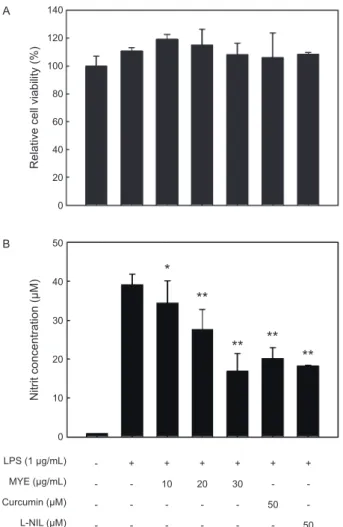
LPS로 유도되는 NO 생성 및 iNOS 발현에 대한 MYE 의 억제 효과
LPS
로자극된RAW 2645.7
세포에서생성되는NO
에대한MYE
의억제효과를알아보기위해서세포를다양한농도(0-30 µg/mL)
의MYE
로1
시간전처리하고, LPS
로24
시간자극하여상층의배지로방출되는
nitrite
생성량을측정하여NO
생성량 을분석하였다. LPS
처리에의해증가된NO
는MYE
의전처리 에의해농도의존적으로현저하게감소하는것으로나타났다(Fig. 1A, P<0.05).
이러한NO
생성의억제효과는MYE
에의 한세포독성으로RAW 264.7
세포가사멸되는것에의해서도 나타날수있으므로,
이러한가능성을배제하고자동일한MYE
처리조건하에서세포생존율의변화를MTS assay
로분석하였 다. Fig. 1B
에나타내었듯이세포의생존율은MYE
처리에의 해뚜렷한변화가없었다.
이러한세포독성실험결과에서MYE
처리에의한NO
생성의억제효과는세포독성에의한것이아 니라는것을확인할수있었다.
다음은
NO
를 생성하는효소인iNOS
의발현에대한MYE
의효과를알아보고자하였다.
전술과같이RAW 264.7
세포 에MYE
를0-30 µg/L
의농도로전처리한이후LPS
로자극하 였고,
이후세포단백질과total RNA
를분리하여각각iNOS
의 단백질과유전자의발현수준을분석하였다. Fig. 2
에나타내었 듯이iNOS
단백질의발현양은LPS
처리에의해현저하게높게유도되는것을알수있었고
, MYE
처리에의해농도의존적으로감소하는경향을나타내었다
. iNOS mRNA
발현양도단백질과유사한경향으로
MYE
처리에의해현저하게감소하는것으로나타났으며
,
그억제효과는전사수준에서효과적으로조 절되고있음을나타낸다(Fig. 2).
NO
는NOS
에 의해L-arginine
으로부터 생성된다. iNOS
는 세균의내독소및염증성사이토카인에의해강하게유도된다(Guha and Mackman, 2001).
병리적인조건하에서iNOS
에 의한NO
의현저한증가는다른염증성매개체들과함께과도 한염증을유발하게되고조직의손상을유발하는것으로알려 져있어염증성손상의주요매개체이다(Nathan, 1992; Pan et al., 2011).
따라서iNOS
의발현또는활성을억제함으로써NO
의생성을억제할수있는화합물은항염증물질로이용될수 있을것이다.
LPS로 유도되는 염증성 사이토카인의 생성에 대한 MYE의 억제효과
LPS
로자극된RAW 2645.7
세포에서생성되는염증촉진성 사이토카인의생성에대한MYE
의효과를ELISA
방법으로분 석하였다. LPS
자극에의해TNF-α (Fig. 3A), IL-1β (Fig. 3B)
및IL-6 (Fig. 3C)
와같은염증촉진성사이토카인의생성량은 크게증가하는것으로나타났고,
이러한증가는다소차이는있지만
MYE
처리에의해현저하게감소하는것으로관찰되었다
(P<0.05~0.01).
또한처리된세포로부터mRNA
를분리하 여RT-PCR
로분석한결과ELISA
경과와유사하게나타났다(Fig. 3D).
염증촉진성사이토카인들은체내에서 다양한면역및염증 반응을조절하는역할을한다
.
세균의LPS
에의해자극된대 식세포는TNF-α
를생성하고분비된TNF-α
및LPS
는IL-1β
애기외톨개 에틸아세테이트 추출물의 항염증 효과
531
와
IL-6
의생성을유도함으로써염증반응을지속시키게된다(Beutler and Cerami, 1989). LPS
에의해유도된TNF-α
는염 증반응의개시를촉진하며지속적인생성은만성염증을유발하 며결국에는패혈성쇼크,
염증,
세포상해성등의다양한생리 학적과정에관여하고있다(Balkwill, 2006; Dinarello, 1999).
IL-1β
는대식세포에서생성되는주요염증촉진성사이토카인으로서
,
세균감염에대한염증성응답의개시및강화에중요한사이토카인이다
(Lebovic et al., 2000). IL-6
도대식세포에서생 성되는중요한염증촉진성사이토카인으로서급성면역응답에 작용한다(Yoshimura, 2006).
본연구에서관찰된결과는MYE
가LPS-
자극에의해유도되는IL-1β, IL-6, TNF-α
의생성을억 제시키는것으로나타났고,
이는MYE
가LPS-
자극에의한염 증성응답의초기단계를억제하고있음을의미한다.
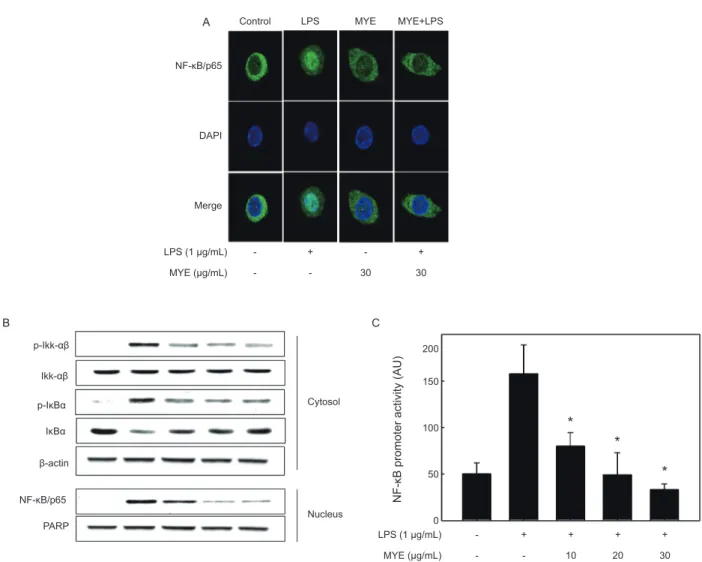
LPS로 유도되는 NF-κB의 활성화에 대한 MYE의 억 제 효과
앞에서서술하였듯이여러염증촉진성사이토카인및
iNOS
의 발현은 주요전사인자인NF-κB
에 의해 조절된다(Zhang and Ghosh, 2000).
따라서LPS
로자극한RAW 264.7
세포를 이용하여NF-κB
의활성화에대한MYE
의처리효과를분석하 였다.
먼저면역형광법으로염색을하고confocal microscopy
로분석한결과를Fig. 4A
에나타내었다.
아무런자극이가해 지지않은상태에서NF-κB/p65 subunit (
녹색)
는DAPI
로염 색된핵(
청색)
주변에대부분분포하는것이관찰되지만, LPS
로 자극한경우녹색인
NF-κB p65
의대부분은 청색인핵과함께분포하는것으로나타났고이는
NF-κB
가활성화되어핵으로이동했음을보여주는결과이다
.
이러한세포에MYE (30 µg/mL)
를전처리한경우NF-κB/p65
는다시핵주변의세포 질에대부분분포하는것으로관찰되었고,
이는MYE
에의해NF-κB
의활성화가현저하게억제되고있음을보여주고있다(Fig. 4A).
NF-κB
의 활성화에 따른 핵으로의 이동은 이를 억제하는단백질인
IκB-α
의인산화에의한분해에의한다(Chen et al.,
Relative cell viability (%) Nitrit concentration (μM)
A
B
140 120 100 80 60 40 20 0
0
*
**
** **
**
10 20 30 40 50
LPS (1 μg/mL) MYE (μg/mL) Curcumin (μM) L-NIL (μM)
- - - -
+ - - -
+ 10 - -
+ 20
- -
+ 30 - -
+ - 50
- + - - 50
A iNOS
Protein
mRNA β-Actin
iNOS β-actin B
LPS (1 μg/mL) MYE (μg/mL)
- -
+ -
+ 10
+ 20
+ 30
HO-1 Western blot
Western blot RT-PCR
β-actin
HO-1 β-actin Nrf2
PARP LPS (1 μg/mL)
MYE (μg/mL) - -
+ -
+ 10
+ 20
+ 30
p-Akt Akt p-JNK JNK p-p38 p38t β-actin
LPS (1 μg/mL) MYE (μg/mL)
- -
+ -
+ 10
+ 20
+ 30
Fig. 1. Effect of MYE on cell viability and nitric oxide (NO) pro- duction in LPS-stimulated RAW 264.7 cells.
Cells pretreated with various concentrations of MYE for 1 h were stimulated with LPS (1 μg/mL) for 24 h (A, B). (A) Effect of MYE on cell viability analyzed by MTS assay. (B) Effect of MYE on NO production in LPS-stimulated RAW 264.7 cells. The culture media of the treated cells were used to measure the amount of nitrite to evaluate NO level. Curcumin and N6- (1- iminoethyl)- L- lysine (L- NIL) were used as positive controls for the inhibitory effect. All data are presented as mean±SD of three independent experiments.
*P<0.05 and **P<0.01 indicate significant differences compared to LPS-only group.
Relative cell viability (%) Nitrit concentration (μM)
B
100 80 60 40 20 0
0
*
**
** **
**
10 20 30 40 50
LPS (1 μg/mL) MYE (μg/mL) Curcumin (μM) L-NIL (μM)
- - - -
+ - - -
+ 10
- -
+ 20 - -
+ 30
- -
+ - 50
- +
- - 50
A iNOS
Protein
mRNA β-Actin
iNOS β-actin B
LPS (1 μg/mL) MYE (μg/mL)
- -
+ -
+ 10
+ 20
+ 30
HO-1 Western blot
Western blot RT-PCR
β-actin HO-1 β-actin Nrf2 PARP LPS (1 μg/mL)
MYE (μg/mL) - -
+ -
+ 10
+ 20
+ 30
p-Akt Akt p-JNK JNK p-p38 p38t β-actin
LPS (1 μg/mL) MYE (μg/mL)
- -
+ -
+ 10
+ 20
+ 30
Fig. 2. Effect of MYE on LPS-induced inducible nitric oxide syn- thase (iNOS) protein and mRNA expression in RAW 264.7 cells.
(A) Western blot analysis of iNOS protein expression. Cells pre- treated with various concentrations of MYE for 1h were stimulated with or without LPS (1 μg/mL) for 16 h. (B) RT-PCR analysis of iNOS mRNA expression. Cells were incubated with various con- centrations of MYE for 1 h, and then stimulated with LPS (1 μg/
mL) for 6 h. mRNA levels of iNOS and β-actin were determined by RT-PCR analysis using respective gene-specific primers. The results presented are representatives of three independent experi- ments.
김보운
ㆍ
김재일ㆍ
김형락ㆍ
변대석532
1995).
따라서MYE
로처리된세포의세포질과핵을분획하여 이들에서NF-κB
및IκB-α
의양및인산화정도를Western blot
으로분석하였다. Fig. 4B
에나타낸것과같이, LPS
자극에의 해세포질의p-Ikk-αβ
의증가로인하여p-IκB-α
의양은증가하는반면
IκB-α
양은유의적감소하는것으로나타남으로써인산화증가에의한
IκB-α
의분해가일어났음을나타내고있다. MYE
처리에의해p-IκB-α
의경우그양이현저하게감소하 는것으로나타남으로써IκB-α
의인산화가억제되었고,
이러한결과는핵에서
NF-κB
의수준이농도의존적으로감소하는결과를초래하였다
.
이상의결과는Fig. 4A
의면역형광염색결 과와도일치하는것이다.
다음은
LPS
로자극된대식세포주에있어NF-κB
의promot- er
활성에대한MYE
의효과를분석하였다.
이를위해RAW 264.7
세포에NF-κB promoter
를가진luciferase construct
를 일시적으로transfection
하고,
이세포를다양한농도의MYE
로2
시간전처리하고이어서LPS
로6
시간동안자극하였다. Fig.
4C
에나타내었듯이luciferase
활성은LPS
자극에의해현저하 게증가하였으며,
이는LPS
자극에의해활성화된NF-κB
가NF-κB
의promoter
를가진luciferase
의발현을크게증가시켰 음을의미한다.
이에대한MYE
의억제효과는10 µg/mL
의낮은농도뿐만아니라
30 µg/mL
농도에이르기까지유의적으로감소하는것으로나타났다
.
이들결과는대식세포주에서LPS
자극에의해유도되는NF-κB
의활성화가MYE
에의해효과 적으로억제되고있음을보여주고있고,
이는MYE
에의한상 기의염증성사이토카인및iNOS
의발현억제는부분적으로NF-κB
활성화경로에의해조절되고있음을의미하는것이다.
MYE에 의한 Nrf2 활성화 효과
최근의많은연구를통하여
HO-1
는뛰어난항염증효과가있는것으로보고되고있다
.
이러한사실을입증하기위하여LPS
로처리된대식세포주에MYE
를처리하여HO-1
와HO-1
의발 현을조절하는전사인자인Nrf2
의발현을분석하였다. Fig. 5
Fig. 3. Effect of MYE on the production of pro-inflammatory cytokines in LPS-stimulated RAW 264.7 cells.
Cells pretreated with various concentrations of MYE were stimulated with or without LPS (1 μg/mL) for 24 h. TNF-α, IL-1β, and IL-6 protein levels in the cell culture media were measured by ELISA (A, B and C, respectively) or their mRNA levels in the cells by RT-PCR analysis (D). Data represent mean±SD of three independent experiments. *P<0.05 and **P<0.01 indicate significant differences compared to LPS-only group.