LPS
로 유도된 RAW 264.7 cell과 마우스 귀 부종 모델을 통한 쌍발이 모자반 에탄올 추출물의 항염증 효과
김민지1, 김민주1, 김꽃봉우리1, 박선희1, 최현덕2, 박소영1, 김지현2, 장미란3, 임무혁4, 안동현1*
1부경대학교식품공학과/식품연구소
2부경대학교수산과학연구소
3식품의약품안전처건강기능식품정책과
4대구대학교식품공학과
Received: October 5, 2016 / Revised: December 1, 2016 / Accepted: January 14, 2017
서 론
체내에서일어나는염증반응은외부로부터물리적, 화학 적자극으로인해손상된부위를복구시키는생물체의보호
반응이다[1]. 하지만지속적인염증반응은오히려점막손상
을촉진시켜통증, 부종, 발열등기능장애를유발하며, 만 성적인염증반응은관절염및암등의발병을유도한다[2].
염증은다양한면역세포및염증매개물질이활성화되면서 나타나며, 그중대식세포(machrophage)는염증반응에관 여하는주요세포로알려져있다. 외부로부터감염초기에는 nitric oxide (NO)와 cytokine을생산하여생체방어에중요 한역할을하며이들분비물에의해대식세포가다시활성
화되어염증반응에관여한다[3]. 대식세포를활성화시키는
대표적인물질로는그람음성균의세포외막에존재하는내 독소인 lipopolysaccharide (LPS)가있으며, 대식세포또는 단핵구를 자극하여 tumor necrosis factor-α (TNF-α), interleukin-1β (IL-1β) 및 IL-6와같은염증매개성 cytokine 들의 분비를촉진한다[4, 5]. 이들은 inducible nitric oxide Anti-Inflammatory Effect of Sargassum patens C. Agardh Ethanol Extract in LPS-induced RAW264.7 Cells and Mouse Ear Edema
Min-Ji Kim1, Min-Ju Kim1, Koth-Bong-Woo-Ri Kim1, Sun-Hee Park1, Hyeun-Deok Choi2, So-YeongPark1, Ji-Hyun Kim2, Mi-Ran Jang3, Moo-Hyeog Im4, and Dong-Hyun Ahn1*
1Department of Food Science & Technology/Institute of Food Science, Pukyong National University, Busan 48513, Republic of Korea
2Institute of Fisheries Sciences, Pukyong National University, Busan 46041, Republic of Korea
3Health Functional Food Policy Division, Ministry of Food and Drug Safety, Chungcheongbuk-do 28519, Republic of Korea
4Department of Food Science and Biotechnology, Daegu University, Gyeongsan 38453, Republic of Korea
The anti-inflammatory effect of Sargassum patens C. Agardh ethanol extract (SPEE) was examined based on the lipopolysaccharide (LPS)-induced inflammatory response in this study. SPEE treatment was not cytotoxic to macrophages compared to the control. The production of NO was suppressed by SPEE by approximately 28% at 100 μg/ml, and levels of interleukin-6, tumor necrosis factor-α, and interleukin-1β decreased in a dose-dependent manner. In addition, the expression of inducible nitric oxide synthase, cyclooxygenase-2, and nuclear factor-κB was suppressed by SPEE treatment. In vivo, croton oil-induced mouse ear edema was attenuated by SPEE and the infiltration of mast cells into the tissue decreased.
Based on these results, SPEE inhibits the release of LPS-induced pro-inflammatory cytokines and media- tors, suggesting that SPEE is a potential agent for anti-inflammatory therapies.
Keywords: Sargassum patens C. Agardh, anti-inflammation, nuclear factor kappa B, mitogen-activated protein kinases, ear edema
*Corresponding author
Tel: +82-51-629-5831, Fax: +82-51-629-5824 E-mail: [email protected]
© 2017, The Korean Society for Microbiology and Biotechnology
synthase (iNOS)와같이산화제인 NO를생성시키는효소 를활성화하게되는데, NO는반응성이높은물질로과잉생 성되면체내의지방질, 단백질과같은생체분자를손상시켜 혈관투과성및부종등의염증반응을촉진한다고보고되고 있다[6]. 또다른주요염증매개인자인 COX는세포막의인 지질로부터 arachidonic acid가유리된후 prostaglandin으 로의 변화를 촉진시키는 효소이며, 이로 인해 발생된
prostaglandin은종양의세포사멸을억제하고혈관생성을유
도하여종양생성에관여한다[7]. 이러한염증성매개분자들 의발현에중요한역할을하는 nuclear transcription factor- kappa B (NF-κB)는다양한 cytokine, chemokine, growth
factor의합성을조절하는전사인자이다[8]. 일반적으로세포
질에서 inhibitor of NF-κBα (IκBα)와결합하여존재하며, 외부자극에의해비정상적으로과잉활성화된 NF-κB는세 포 핵 안으로 들어가 전사인자로서 작용하여 염증관련 cytokine 및염증을매개하는생체효소(iNOS, COX-2)를합
성한다[9]. 이와더불어 MAPKs는대식세포를포함한다양
한세포의활성을조절하고세포의증식및성장에관여하 는것으로 알려져있으며, extra cellular signal-regulated kinase (ERK), c-jun NH2-terminal kinase (JNK), p-38과
같은주요 3개의인자가존재한다[10]. 따라서이들의활성
조절은염증반응을조절하여다양한질병을치료하기위한 핵심요소로서여겨진다. 최근천연물로부터염증반응을효 과적으로억제하는물질을탐색하기위해많은연구가진행 되고있으며, 그중각광받고있는천연물의하나로해조류 를들수있다. 해조류는종류와시기에따라다양한성분의 차이를보이며육상식물과는다른구조로항균[11], 고지혈 증개선[12], 콜레스테롤침착방지[13] 등의생리활성이보고 되고있으며, 그중에서도 갈조류는 fucoidan, phycocolloids, phlorotannins 등과같은생리활성물질로부터항산화, 항응 고, 항암등의다양한생리활성을보인다고알려져있다[14].
따라서본연구에서는모자반과에속하는갈조류인쌍발이 모자반(Sargassum patens C. Agardh)을이용하여항염증 효과를알아보고자한다.
재료 및 방법
실험 재료
본실험에 사용한 쌍발이모자반(Sargassum patens C.
Agardh)은 2015년부산연화리에서채취하였으며이를담
수로깨끗이수세하고동결건조한후분말화하고진공포 장하여−20℃에서저장하며사용하였다.
에탄올 추출방법
쌍발이모자반건조분말에 10배의 95% 에탄올을가하고,
교반기(H-0820, Dongwon Science Co., Korea)를이용하여 24시간 동안 상온에서 교반하여추출하였다. 원심분리기 (UNION 32R, Hanil Co., Korea)를이용하여 1,977 ×g에서 10분간원심분리한후상층액을취하였고, 이후남은잔사를 이와동일한방법으로 2회반복추출하였다. 추출한상층액 은 37℃에서감압농축기(RE200, Yamoto Co., Japan)로농 축하였으며, 농축하여건조된시료는−20℃에서보관하며실 험에이용하였다.
실험 동물
ICR 마우스(생후 8주령, 수컷)을오리엔트바이오(Orient
Co., Korea)로부터구입하여귀부종및귀조직실험에사
용하였으며, 마우스는온도 20 ± 2℃, 습도 50 ± 10%, 12시간 명암주기가유지되는동물실에서 1주일간예비사육한후실 험에사용하였다. 본동물실험은부경대학교동물실험윤리 위원회로부터동물실험승인을받아수행하였다(2015-04).
세포배양
RAW 264.7 세포는 한국세포주은행(KCLB 40071)에서
분양받아 사용하였으며, DMEM에 10 mg/ml penicillin- streptomycin과 100 mg/ml inactivated fetal bovine serum을 첨가한배지를배양액으로 37℃, 5% CO2조건에서배양하 였다. 실험과정의모든세포는 80−90% 정도의밀도로자랐 을때계대배양하였고, 20 passages를넘기지않은세포만 사용하였다.
세포 독성 측정
RAW 264.7 cell 1 × 106 cells/ml를 well plate에 분주하 고 20시간 전배양 후, 1 μg/ml의 LPS와추출물을농도별 (0.1, 1, 10, 50, 100 μg/ml)로 첨가하여 37℃, 5% CO2
incubator (MCO-15AC, Sanyo, Japan)에서 24시간배양하 였다. 배양 후, 5 mg/ml 농도의 3-[4, 5-dimethylthiazol-2- yl]-2, 5-diphenyl tetrazolium bromide (MTT) 시약을첨가 하여 2시간재배양하고이를 4℃, 879 ×g에서 10분간원심 분리(UNION 32R, Hanil Co., Korea)하여상층액을제거하 였다. 그후, 각 well에 DMSO를첨가하고이를 microplate reader (Model 550, Bio-rad, USA)를이용하여 540 nm에 서흡광도(obtical density (O.D))를측정하였다. 세포증식능 은다음식에의해계산하였다.
Cytotoxicity (%) = sample 흡광도/control 흡광도× 100
Nitric Oxides 생성량 측정
Lee 등[10]의방법에따라 RAW 264.7 cell은 DMEM 배 지를이용하여 2.5 × 105 cells/ml로조절한후 24 well plate
에접종하고 5% CO2 incubator (MCO-15AC, Sanyo, Japan) 에서 20시간전배양하였다. 세포에 1 μg/ml의 LPS와 0.1, 1, 10, 50, 100 μg/ml의추출물을처리하여 24시간재배양 하였다. 배양액의 상층액을 얻은 후, 동량의 griess 시약 (10 mg/ml sulfanilamide + 1 mg/ml naphthylendiamine dihydrochloride, 1:1)을첨가하여실온에서 10분간반응시 키고, microplate reader (Model 550, Bio-rad, USA)를 이
용하여 540 nm에서흡광도를측정하였다. 세포배양액내
NO의농도는 sodium nitrite (NaNO2)의농도별표준곡선과 비교하여산출하였다.
염증 관련 cytokines 분비량 측정
세포배양액내의 TNF-α, IL-6 및 IL-1β cytokine의분비 량을 ELISA kit (Mouse ELISA set, BD Bioscience, USA) 를이용하여 측정하였다. 이를 위해 ELISA microplate에 capture antibody로 anti-mouse TNF-α, IL-6 및 IL-1β를 분주하여 4℃에서하룻밤동안 coating시켰다. 이를 0.5 mg/
ml Tween 20이포함된 PBST로세척하고 100 mg/ml FBS 용액으로 blocking 하였다. PBST로세척한뒤, 배양액을분 주하고실온에서 2시간반응시켰다. 다시 PBST로세척한뒤 희석한 biotinylated anti-mouse TNF-α, IL-6 detection antibody와 streptavidin-horseradish peroxidase conjugate를 첨가하여 실온에서 1시간 반응시켰다. IL-1β의 경우, biotinylated anti-mouse IL-1β detection antibody를첨가 하고 1시간 반응 후, streptavidin-horseradish peroxidase conjugate를첨가하여 30분반응시켰다. 그후, 이를다시 PBST로세척한다음, o-phenylenediamine-dihydrochloride
(OPD) 용액을첨가하여실온에서 30분동안암반응시켰다.
2 N H2SO4로 반응을 종료시킨 후, microplate reader (Model 550, Bio-rad, USA)를이용하여 490 nm에서흡광도 를측정하였다.
단백질 분리와 Western blot 분석
배양이끝난세포들을수집하여 cytosol extraction lysis buffer (50 mM HEPES (pH 7.4), 150 mM NaCl, 5 mM EDTA, 1% deoxycholate, 5 mM phenylmethylsulfonyl fluoride (PMSF), 1μg/ml aprotinin, 1% Triton X-100, and 0.1% NP-40) 및 nulclear extraction lysis buffer (10 mM HEPES, 100 mM NaCl, 1.5 mM MgCl2, 0.1 mM EDTA, 0.1 mM DTT)를각각사용하여 30분간 4℃에서용해시켰다. 분리된단백질들의농도를측정한후, 10% sodium dodecyl sulfate (SDS)- polyacrylamide gel electrophoresis (PAGE)를 이용하여 분리하고 polyvinylidene difluoride membrane (PVDF, Bio-rad)에 70 mA에서 1시간 30분동안전이시켰다. Kim 등[15]의방법에따라각각의 membrane을적정항체및
enhanced chemiluminescence (Termo Scientific, USA) 용액 을이용하여감광하였으며, Gene tool (Syngene software, Synoptics Ltd., UK)를이용하여가시화및정량하였다.
귀 부종 측정 및 조직 관찰
6주령의 ICR 마우스에 추출물을 10, 50 및 250 mg/kg·
body weight 농도로 200 μl씩경구투여하였다. 한시간후, 오 른쪽귀에 25 mg/ml croton oil을 20 μl/ear 농도로도포하였 다. 귀두께는 croton oil을처리하고 5시간후에측정하였으
며 croton oil 처리한후두께의증가를부종의형성으로간
주하였다. 조직관찰은 ICR 마우스의오른쪽귀에추출물을
100 mg/ml 농도로 20 μl씩 도포하고 15분 뒤, 50 mg/ml croton oil을 20 μl씩도포하였다. 6시간뒤, diethylether로 마취시키고, 귀조직을절제하여 100 mg/ml formaldehyde 에 72시간고정하였다. 고정후파라핀블록을만들어박편 을제조하고 hematoxylin-eosin 및 toluidine-blue 염색을하 여조직을관찰하였다.
Edema formation (% of control) =
Ear thickness of sample/Ear thickness of control × 100
통계처리
모든실험결과에대한유의차검정은 SAS software (SAS Institute, Inc., USA)에서평균값을분산분석한후, Duncan's multiple range test 법에따라 p < 0.05 수준에서검정하였다.
결과 및 고찰
세포 독성 측정
추출물처리후대식세포로부터염증매개물질이감소되 는것은세포독성으로인한 RAW 264.7의사멸에의한결과
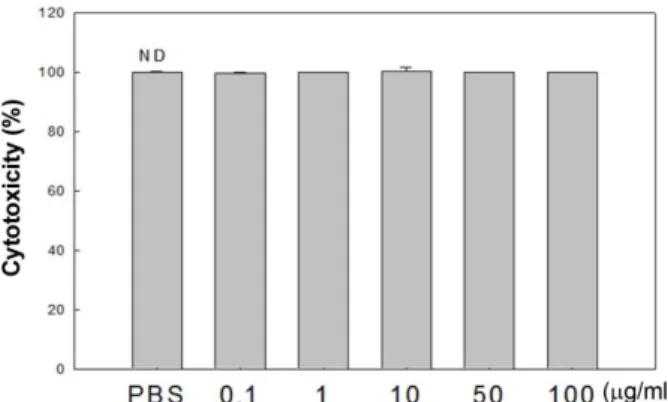
Fig. 1. Effect of SPEE on the cytotoxicity of RAW 264.7 cells.
Cytotoxicity (%) = (sample O. D./control O. D.) × 100. ND means no significant difference.
일수도있으므로이러한가능성을배제하고자 MTT assay
를수행하여 RAW 264.7 cell에미치는추출물의세포독성
을측정하였다. 추출물을 0.1, 1, 10, 50 및 100 μg/ml의농 도로첨가하여배양한결과, 모든처리농도에서유의적인 차이를보이지않음을관찰하였다(Fig. 1). 따라서쌍발이에 탄올추출물이대식세포에독성을나타내지않은것을확인 하였으며이후의모든실험에서 0.1−100 μg/ml 농도로쌍발 이모자반추출물을처리하여실험을수행하였다.
Nitric oxide 생성 억제 효과
NO는활성산소의일종으로높은반응성을가진생체생 성분자로서염증유발에중요한역할을하며, 혈관확장, 세포 독성, 조직의손상등으로염증반응을심화시키는것으로알 려져있다[16]. 따라서 LPS로활성화된 RAW 264.7 세포로 부터생성되는 NO에미치는쌍발이모자반에탄올추출물
의효과를측정하기위해추출물을각농도별로 (0.1, 1, 10,
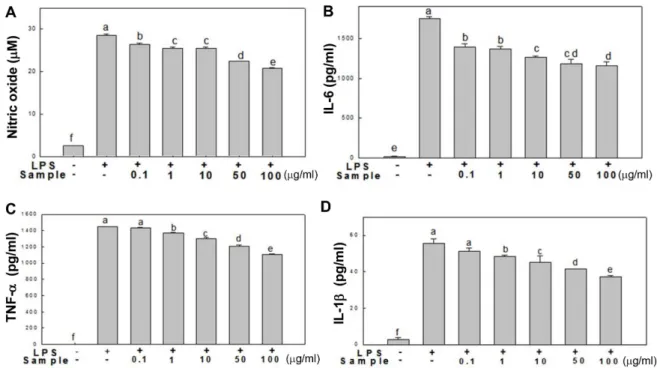
50 및 100 μg/ml) 첨가하고생성된 NO를 griess 시약을이 용하여측정하였다. 그결과(Fig. 2A), RAW 264.7 세포만배 양한대조군에서 NO의농도는 2.58 μM로매우낮게측정 되었으며, LPS를처리한군에서는 NO의농도가 28.5 μM로 현저히증가되었다. 쌍발이모자반에탄올추출물처리군에
서는 NO의분비량이 LPS 처리군보다감소함을보였으며,
특히 50 μg/ml 및 100 μg/ml 농도로추출물을처리하였을때 그분비량이각각 22.38 μM 및 20.73 μM으로약 20% 이상 감소됨을확인하였다. 이는갈조류이며모자반과에속하는 참모자반에탄올추출물이 NO 분비량감소에효과적인결 과와유사하다[17].
염증 관련 cytokines 생성 억제 효과
체내의염증반응은염증매개효소및 NO 등과함께염증 성 cytokine이동반되며, 대표적인 cytokine으로는 TNF-α, IL-6, 및 IL-1β이다[18]. 종양괴사인자인 TNF-α는체내에 서대식세포나림프구등에의해생성되며종양세포에세포 독성을나타내고만성염증과관련되어있다[19]. IL-6는단 핵구를포함한다양한면역세포에서분비되며염증유발사 이토카인의하나로초기면역 반응에서중요한역할을한 다. 또한, IL-1β는 TNF-α, IL-6와 함께 대표적인 염증성 cytokine으로국소염증을발생시키고, T 세포의활성화, B세
포의성숙및 NK cell을활성화에관여하며 NO를생성하게
하는매개물질로알려져있다[20]. 따라서쌍발이모자반에
탄올추출물이염증성 cytokine의생성에미치는영향을알
아보기위하여추출물을농도별로처리하여 ELISA 방법으
로분비량을측정하였다. 그결과(Fig. 2B−D), 세가지 cytokine (IL-6, TNF-α및 IL-1β) 모두유의적으로그분비량이감소
Fig. 2. Inhibitory effect of SPEE on the production of nitric oxide (A), IL-6 (B), TNF-α (C), and IL-1β (D) in RAW 264.7 cells. RAW 264.7 cells were treated with the indicated concentrations of SPEE (0.1, 1, 10, 50, and 100 μg/ml) in the presence or absence of LPS (1μg/ml) for 24 h. Culture supernatants were then isolated and analyzed using the Griess reagent for nitric oxides and ELISA kit for cytokines. (a−f) indicates significant differences (p < 0.05).
를보였다. IL-6의경우, 추출물 0.1 μg/ml로처리였을때약 20%의분비량감소를보였으며, 100 μg/ml 처리농도에서 약 33.7%까지감소함을나타내었다. TNF-α및 IL-1β의경 우에는농도의존적감소를보이며최고처리농도인 100 μg/
ml에서각각약 23.8% 및 32.7% 감소됨을보였다. 이상의결
과를통해쌍발이모자반에탄올추출물이 RAW 264.7 세포
에서염증매개성 cytokine을효과적으로억제하여항염증
효과에관여하는것을확인하였다.
iNOS 및 COX-2 발현 억제 효과
iNOS는세포내에는존재하지않으나외부자극에의해
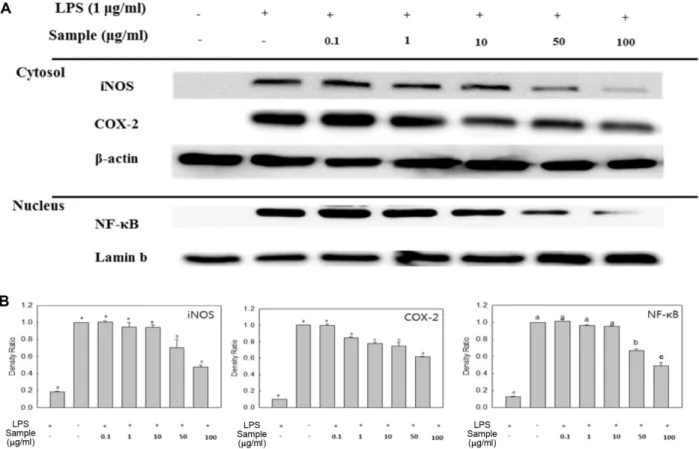
유도가되면 NO를생성하게되며과도한 NO의생성은염 증유발등병리적으로중요한작용을한다고알려져있다 [5, 21]. 또한, COX-2는혈관활성물질인 PGE2의형성에관 여하며유리된 PGE2의혈관투과성이증대되면서염증을유 발한다[22]. 따라서염증반응에서분비되는 iNOS 및 COX- 2의생성량을측정하여추출물의항염증효과를확인하였다. RAW 264.7 세포에추출물을 0.1−100 μg/ml 농도로처리하고
iNOS 및 COX-2의발현량을측정한결과(Fig. 3), LPS 단독처 리구에의해각단백질의발현량이현저히증가하였으나, 추 출물을처리하였을때그발현량이감소하는것을확인할수 가있었다. 특히 50 및 100 μg/ml 농도에서효과적인억제효 과를나타내었으며, iNOS의발현량의경우에는앞선결과에 서보인 NO의생성량억제에관여한것으로사료된다.
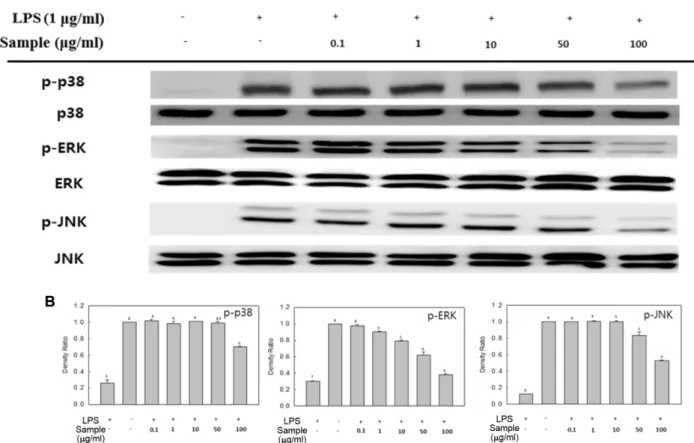
NF-κB 및 MAPKs 발현 억제 효과
NF-κB는염증반응과관련된유전자의 promoter에결합 하며활성화된 NF-κB는 COX-2 및 iNOS 발현에주로관여
하는것으로알려져있다[23]. MAPKs는대식세포활성을매
개하는또다른중요신호전달계로서핵내로이동하여다 른면역활성인자들을생성을유도하여염증반응을더욱활 성화시킨다[24]. 따라서염증반응에서 NF-κB 및 MAPKs의 발현량억제를확인하여항염증효과를확인하였다(Fig. 3,
4). LPS 단독처리구에의해각단백질의발현량이현저히증
가하였으나, 추출물을처리하였을때그발현량이감소하는 경향을나타내었다. 처리된추출물농도 10 μg/ml까지는감
Fig. 3. Effect of SPEE on LPS-induced iNOS, COX-2, and NF-κB p65 expression in RAW 246.7 cells. The levels of iNOS, COX-2 in the cytosolic protein and the p65 subunit of NF-κB in nuclear protein were determined by a western blot analysis (A). RAW 264.7 cells were treated with the indicated concentrations of SPEE (0.1, 1, 10, 50, and 100 μg/ml) and LPS (1 μg/ml) for 18 h or 30 min and the proteins were detected using specific antibodies. For quantification, the expression data were normalized to the β-actin or lamin b signal (B).
Means with different letters (a−e) above the bars are significantly different (p < 0.05).
소의폭이크지않으나 10 μg/ml 이상의농도에서발현량이 약 20% 이상감소하였으며, 특히최고처리농도인 100 μg/
ml 농도에서는발현량이현저히감소하는것을확인할수가 있었다. 이러한결과를종합해볼때, 쌍발이모자반에탄올 추출물의 NF-κB 활성억제및 MAPKs 발현량억제를통해 신호전달물질및염증관련효소들의생성을억제함으로 써항염증효과를나타낸다고생각된다.
귀 부종 억제 효과 및 조직 관찰
대표적인염증반응인부종은외부환경에의해손상된피 부부위를복구시키려는일련의생체과정으로서혈관확장
등과함께나타난다[25]. 현재항염증제로서사용되고있는
합성스테로이드제인 prednisolone 10 및 50 mg/kg과추출 물 10, 50 및 250 mg/kg 농도를 200 μl씩마우스에경구투 여한후, croton oil로염증유발하고귀두께를측정하였다. 그결과(Fig. 5), 부종을유발한 control과비교하여모든농 도에서유의적으로귀두께가감소한것을확인하였다. 특히,
추출물 250 mg/kg 농도에서 positive control인 prednisolone 처 리구와비교하였을때, prednisolone 50 mg/kg 처리보다다 소감소함을보였다. 이러한추출물의귀부종완화효과는 조직관찰결과에서도나타났으며, croton oil로부종을유발
한마우스귀조직에서 croton oil만을처리한경우에비해
추출물을 100 mg/kg 농도로처리한경우 prednisolone 처리 구와유사한정도로경피및진피두께가얇아진것을확인 하였다(Fig. 4B). 또한, toluidine-blue 염색을통해조직내 mast cell 침윤정도를확인한결과(Fig. 4C), 추출물의처리 가조직내 mast cell 침윤을현저히억제함을보였다. 외부 자극으로부터활성화된신체내 mast cell은다양한 protease 나히스타민과같은혈관확장물질을분비함으로써병원균 에대한숙주의방어기작을하는것으로알려져있다[26]. 따
라서 croton oil을피부에도포하여유도한급성염증반응에
서쌍발이모자반에탄올추출물의처리가손상된조직내
로 mast cell의침윤을감소시킴으로써부종완화에영향을
미친것으로사료된다.
Fig. 4. Effect of SPEE on MAPKs expression in LPS-induced RAW 246.7 cells. The levels of p-p38, p-ERK, and p-JNK in the cytosolic protein were determined by western blot analysis (A). RAW 264.7 cells were treated with the indicated concentrations of SPEE (0.1, 1, 10, 50, and 100 μg/ml) and LPS (1 μg/ml) for 30 min, and the proteins were detected using specific antibodies. For quantification, the expression data were normalized to the total MAPKs signal (B). Means with different letters (a−f) above the bars are significantly different (p < 0.05).
요 약
본연구에서는쌍발이모자반의항염증효과를알아보기 위해 LPS에의해염증반응이유도된 RAW 264.7 세포에대 한쌍발이모자반에탄올추출물의항염증효과를살펴보았 다. 세포내염증매개성 cytokine (IL-6, TNF-α및 IL-1β) 분 비량의경우농도의존적인감소효과를보였다. 또한추출 물이 iNOS, COX-2, NF-κB 및 MAPKs 발현억제에미치는
효과를알아본결과, LPS 단독처리구에의해각단백질의
발현량이현저히증가하였으나, 50 μg/ml 이상의농도로추 출물을처리하였을때그발현량이효과적으로감소하는것 을확인할수가있었다. 귀부종억제효과및조직관찰을 수행한 결과, 추출물 250 mg/kg 농도에서 prednisolone
50 mg/kg 처리보다귀부종이다소감소함을보였으며, 조
직관찰결과쌍발이모자반에탄올추출물을처리함으로써 귀조직의경피및진피두께가얇아지고, 조직내 mast cell 침윤을현저히억제함을보였다. 쌍발이모자반에탄올이보
이는항염증효과는해조류에탄올추출물유래 polyphenol
계열의화합물의영향이크다고생각되며현재까지쌍발이 모자반내의항염증효능물질에관한연구는보고되지않 고있다. 따라서본논문의결과를바탕으로향후유효성분 에관한분리연구가진행된다면쌍발이모자반에탄올추 출물의천연염증치료소재로이용될가치가충분할것으 로사료된다.
Acknowledgments
This research was supported by Basic Science Research Program through the National Research Foundation of Korea (NRF) funded by the Ministry of Education (2012R1A6A1028677) and this work was supported by the National Research Foundation of Korea (NRF) and Center for Women In Science, Engineering and Technology (WISET) grant funded by the Ministry of Science, ICT & Future Planning of Korea (MSIP) under the team research program for female engineer- ing students (WISET-2016-077).
Fig. 5. SPEE-mediated inhibition of croton oil-induced mouse ear edema (A). Photomicrographs of transverse sections of mice ears sensitized with topical application of 50 mg/ml croton oil in acetone (a−c) or acetone alone (d, non-inflamed), stained with hematoxylin–
eosin (B) or toluidine-blue (C). Photomicrographs recorded under light microscopy (magnification: 200×). Treatments: vehicle 20 mg/
ml Tween 80 (a), prednisolone 0.08 mg/ear (b), SPEE 20 μl/ear (c), and acetone (d). The numbers 1 and 2 indicate dermis and epidermis, respectively. (a−f) indicates significantly different results (p < 0.05).
References
1. Lee ST, Jeong YR, Ha MH, Kim SH, Byun MW, Jo SK. 2000. Induc- tion of nitric oxide and TNF-α by herbal plant extract in mouse macrophage. J. Korean Soc. Food Sci. Nutr. 29: 342-348.
2. Ljung T, Lundberg S, Varsanyi M, Ohansson C, Schmidt PT, Herulf M, et al. 2006. Rectal nitric oxide as biomaker in the treatment of inflammatory bowel disease: responders versus non-responders. World Gastroenterol. 12: 3386-3392.
3. Kang BK, Kim KBWR, Kim MJ, Bark SW, Pak WM, Kim BR, et al.
2014. Anti-inflammatory activity of an ethanol extract of Lami- naria japonica root on lipopolysaccharide-induced inflamma- tory responses in RAW 264.7 cells. Korean J. Food Sci. Technol.
46: 729-733.
4. Kim DH, Park SJ, Jung JY, Kim SC, Byun SH. 2009. Anti-inflam- matory effects of the aqueous extract of Hwangnyenhaedok- tang in LPS-activated macrophage cells. Korean J. Herbol. 24:
39-47.
5. Lim HR, Shin SW. 2010. Effects of the essential oil components from Ligusticum chuanxiong on proinflammatory mediators of RAW264.7 macrophage cells. Korean Sci. Pharm. 16: 259-264.
6. Lee TH, Kwak HB, Kim HH, Lee ZH, Chung DK, Baek NI, et al.
2007. Methanol extracts of Stewartia koreana inhibit cyclooxy- genase-2 (COX-2) and inducible nitric oxide synthase (iNOS) gene expression by blocking NF-kappaB trans activation in LPS-activated RAW 264.7 cells. Mol. Cells. 23: 398-404.
7. Bishop-Bailey D, Calatayud S, Warner TD, Hla T, Mitchell JA.
2002. Prostaglandins and the regulation of tumor growth. J.
Environ. Pathol. Tox. Oncol. 21: 93-101.
8. Ghosh S, Hayden HS. 2008. New regulators of NF-κB in inflam- mation. Nat. Rev. Immunol. 8: 837-848.
9. Majdalawieh A, Ro HS. 2010. Regulation of IκBα function and NF-κB signaling: AEBP1 is a novel proinflammatory mediator in macrophages. Mediators Inflamm. 2010: 823-821.
10. McDaniel ML, Kwon G, Hill JR, Marshall CA, Corbett JA. 1996.
Cytokines and nitric oxides in islet inflammation and diabetes.
Proc. Soc. Exp. Biol. Med. 211: 24-32.
11. Nagayama K, Iwamura Y, Shibata T, Hirayama I, Nakamura T.
2002. Bactericidal activity of phlorotannins from the brown alga Ecklonia Kurome. J. Antimicrob. Chemother. 50: 889-893.
12. See Jh, Kim ND, Choi JS, Kim YJ, Moon YH, Lim SY, et al. 1998.
Inhibitory effects of the methanolic extract of an edible brown alga, Ecklonia stolonifera and its component, phloroglucino on aflatoxin B1 mutagenicity in vitro (Ames test) and on benzo(a) pyrene or N-methyl N-nitrosourea clastogenicity in vivo (mouse micronucleus test). Nat. Prod. Sci. 4: 105-114.
13. Kim HS, Kim GJ. 1998. Effects of the feeding Hizikia fusiforme (Harvey) O kamura on lipid composition of serum in dietary hyperlipidemic rats. J. Korean Soc. Food Sci. Nutr. 27: 718-723.
14. Halliwell B, Gutteridge JMC. 1999. Antioxidant defenses. In Free radicals in Biology and Medicine. 3rd ed. Oxford Science Publications, Oxford, UK. pp. 105-159.
15. Kim MJ, Bae NY, Kim KBWR, Park JH, Park SH, Choi JS, et al. 2016.
Anti-inflammatory effect of grateloupia imbricata holmes eth- anol extract on LPS-induced RAW 264.7 cells. J. Korean Soc.
Food Sci. Nutr. 5: 181-187.
16. Nathan C. 1992. Nitric oxide as a secretory product of mamma- lian cells. FASEB J. 6: 3051-3064.
17. Jeong DH, Kim KBWR, Kim MJ, Kang BK, Bark SW, Pak WM, et al.
2014. Anti-inflammatory effect of ethanol extract from Sargas- sum fulvellum on lipopolysaccharide induced inflammatory responses in RAW 264.7 cells and mice ears. J. Korean Soc. Food Sci. Nutr. 43: 1158-1165.
18. Park SG, Jegal KH, Jung JY, Back YD, Byun SH, Kim YW, et al.
2014. Leonuri fructus ameliorates acute inflammation via the inhibition of NF-κB-mediated nitric oxide and pro-inflamma- tory cytokine production. Korean J. Orient. Physiol. Pathol. 28:
178-185.
19. Moon JH, Go H, Shin SM, Kim KT. 2013. Anti-inflammatory effect of extracts from Ligustrum obtusifolium S. Fruits in RAW 264.7 macrophages. J. Soc. Korean Med. Diagn. 17: 263-273.
20. Masters SL, Simon A, Aksentijevich I, Kastner DL. 2009. Horror auto-inflammaticus: the molecular pathophysiology of autoin- flammatory disease. Ann. Rev. Immunol. 27: 621-668.
21. Yun HY, Dawson VL, Dawson TM. 1996. Neurobiology of nitric oxide. Crit. Rev. Neurobiol. 10: 291-316.
22. Duerksen-Hughes PJ, Day D, Laster SM, Zacharidase NA, Aquino L, Gooding LR. 1992. Both tumor necrosis factor and nitric oxide participate in lysis of simian virus 40-transformed cells by activated macrophages. J. Immuno. 149: 2-14.
23. Celec P. 2004. Nuclear factor kappa B-molecular biomedicine:
the next generation. Biomed. Pharmacother. 58: 365-371.
24. Eum WS, Lee KJ, Kim DW, Lim SS, Kang IJ, Park J, et al. 2013.
Anti-inflammatory effects of extracts from Caesalpinia sappan L. on skin inflammation. J. Korean Soc. Food Sci. Nutr. 42: 1513- 1517.
25. Hahn DH, Sur BJ, Han DO, Park JH, Jung ET, Lee HJ, et al. 2008.
Anti-inflammatory activity of dandelion in mice. Korean J. Ori.
Med. Physiol. Phthol. 22: 810-814.
26. Tete S, Tripodi D, Rosati M, Conti F, Maccauro G, Saggini A, et al.
2008. The role of mast cells in innate and adaptive immunity.
Life Sci. J. 18: 891-896.