152
Copyright © 2021 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815
서 론
염증반응은미생물감염, 내독소, 조직손상과같은위해성자 극에대한방어기능으로, 이는생명체의구조와기능을정상적 으로유지시키기위한방어기능이다. 염증반응은시간이지남 에따라염증촉진성매개체들의생성은감소되고, 항염증성매 개체들이증가됨으로써스스로염증반응이억제되는조절기전을가지고있다(Libby and Hansson, 2015). 대식세포
(macro-phages)는체내의염증반응에중요한역할을하는것으로알려 져있다. 대식세포는 interferon-γ (IFN-γ), interleukin (IL)-1β,
IL-6, tumor necrosis factor-α (TNF-α)와같은염증촉진성
cy-tokines, 그리고세균세포막성분인 lipopolysaccharides (LPS) 등의자극에의해활성화된다(Xie et al., 1994). 활성화된대 식세포는염증촉진성 cytokines 이외에 inducible nitric oxide
synthase (iNOS) 및 cyclooxygenase-2 (COX-2)와같은효소 의발현을통해 nitric oxide (NO) 및 prostaglandin E2 (PGE2) 와같은다양한염증매개분자들을생성하게되고
(Marks-Kon-czalik et al., 1998; Zhang and Ghosh, 2000), 이들매개체들의
지속적인생성은다양한만성염증성질환의주요한원인으로
알려져있다. 부작용이적은항염증활성을지닌천연물의발견
은염증치료제개발을위한방편으로관심을끌고있다.
대식세포의 염증촉진성 cytokines 및 단백질들의 발현은
LPS로 유도된 RAW 264.7 대식세포에 대한 대황(
Eisenia bicyclis) 헥산
분획물의 항염증 효과
김보운·최창근
1·김재일·김형락*
부경대학교 식품영양학과, 1부경대학교 생태공학과
Anti-Inflammatory Effect of Hexane Fraction from Eisenia bicyclis on
Lipopolysaccharides-Treated RAW 264.7 Cells
Bowoon Kim, Chang-Geun Choi
1, Jae-Il Kim and Hyeung-Rak Kim*
Department of Food Science and Nutrition, Pukyong National University, Busan 48513, Korea
1Department of Ecological Engineering, Pukyong National University, Busan 48513, Korea
Eisenia bicyclis
is known to have secondary metabolites exhibiting various biological activities. In a preliminary study, the n-hexane fraction obtained from the ethanolic extract ofE. bicyclis
showed higher anti-inflammatory activ-ity than the ethyl acetate and butyl alcohol fractions based on the inhibition of lipopolysaccharide (LPS)-stimulated nitric oxide (NO) production in RAW 264.7 cells. Using this fraction (E. bicyclis
hexane fraction, EHF), we inves-tigated the molecular mechanisms underlying its anti-inflammatory effect in LPS-stimulated RAW 264.7 cells. Pre-treatment of the cells with up to 50 µg/mL EHF significantly inhibited NO and prostaglandin E2 production as well as their responsible enzyme proteins and mRNAs, in a dose-dependent manner (P<0.05). Similarly, EHF markedly reduced the production of pro-inflammatory cytokines, such as interleukin (IL)-1β, IL-6 and tumor necrosis factor (TNF)-α as well as their mRNA levels. Nuclear translocation of nuclear factor-kappa B (NF-κB) was strongly sup-pressed by EHF treatment. EHF significantly reduced the phosphorylation of mitogen-activated protein kinases and phosphatidylinositol 3-kinase/Akt in LPS-stimulated cells. Moreover, EHF reduced ear edema in phorbol myristate acetate (PMA)-induced mice. These results indicate that EHF contains potent anti-inflammatory compounds, which may be used as a dietary supplement for the prevention of inflammatory diseases.Keywords: Anti-inflammatory effect,
Eisenia bicyclis
, iNOS, NF-κB, Pro-inflammatory cytokines*Corresponding author: Tel: +82. 51. 629. 5847 Fax: +82. 51. 629. 5842
E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 29 January 2021; Revised 15 February 2021; Accepted 19 February 2021
저자 직위: 김보운(대학원생), 최창근(교수), 김재일(교수), 김형락(교수)
https://doi.org/10.5657/KFAS.2021.0152
nuclear factor kappa-B (NF-κB)에의해전사수준에서조절된 다(Marks-Konczalik et al., 1998; Rahman et al., 2006). 정상 상태의세포에서 NF-κB는 inhibitor of kappa B (IκB)와결합 한상태로불활성형태로세포질에존재한다(D'Acquisto et al.,
1997; Rahman et al., 2006). LPS, cytokines과같은자극이주 어지는경우 IκB는 IκB kinase (Ikk)에의해인산화와유비퀴 톤화(ubiquitination)를통해 proteasome에의해분해되고, 유 리된 NF-κB는핵으로이동하여염증관련유전자의발현을유 도하게된다(Elliott et al., 2003). 또한 extracellular
signal-reg-ulated kinase (ERK), c-Jun N-terminal kinase (JNK), p38 ki-nase를포함하는 mitogen-activated protein kinases (MAPKs) 와 PI3K/Akt (PKB)와같은단백질인산화효소에의해 NF-κB 가활성화되어염증반응을촉진하게된다(Marks-Konczalik et
al., 1998; Kaminska, 2005).
갈조류는 다양한 종류의 생리활성물질을 함유하고 있으므
로 이들을이용하기 위하여광범위한 연구가 지속되고 있다
(Thomas and Kim, 2011). 대황(Eisenia bicyclis)은다시마과
에속하는갈조류로써우리나라와일본의해안에다량존재하
는것으로알려져있다. 대황의주요한 2차대사산물들은플로
로탄닌(phlorotannin)으로항염증(Jung et al., 2013; Paudel et
al., 2014; Yayeh et al., 2014) 항산화(Kim et al., 2011; Kwon
et al., 2013), 간보호(Choi et al., 2015a), 항균(Eom et al., 2014;
Kim et al., 2018), 항치매(Choi et al., 2015b), 항앨러지
(Sugi-ura et al., 2009), 항암활성(Thomas and Kim, 2011) 등이보 고되고있다.
대황주정추출물을이용한예비실험에서헥산(hexane), 에틸
아세테이트(ethyl acetate) 및부탄올(butanol) 획분으로분리하
여항염증활성을분석한결과헥산획분의항염증활성이가장 높은것으로확인되었다. 본연구에서는대황의헥산분획물의 항염증효과및관련분자적기전을 LPS (lipopolysaccharide) 로자극한 RAW 264.7 세포를이용하여분석함으로써, 염증성 질환의발병을예방또는지연시킬수있는건강기능성소재로 서의이용가능성을검토하였다.
재료 및 방법
추출과 분획
독도 인근에서 채취한 대황[Eisenia bicyclis (Kjellman)
Setchell]의건조분말 1 kg을환류냉각기가부착된집기병에담 고주정[95% ethanol (EtOH), v/v] 6 L를넣어가열, 2회추 출하여(72°C, 3시간) 얻은추출액을여과하여 rotary vacuum evaporator로농축하였다. 에탄올농축물(122 g)을 H2O:EtOH (9:1, v/v)의혼합용매로녹인후동량의 n-hexane을넣어분액 깔대기에 평형화시켜상층액의 n-hexane 가용부를 분리하였
다. 분리된 n-hexane 획분(E. bicyclis hexane fraction, EHF)을
농축하여실험에사용하였다.
세포 배양 및 처리
RAW 264.7 세포 (ATCC, Rockville, MD, USA)는 10%
fe-tal bovine serum (FBS) (GIBCO, Grand Island, NY, USA)와
penicillin (100 units/mL), streptomycin sulfate (100 µg/mL)을 첨가한 Dulbecco’s modified Eagle’s medium (DMEM) (Life
Technology, Carlsbad, CA, USA)을 사용하였고, 5% CO2,
37°C 배양기에서배양하였다. Cell culture plate에 RAW 264.7
cell이 70-80% 정도채워지면 phosphate-buffered saline (PBS) 로 한번씻어낸후, 계대배양하였다. EHF는 100% dimethyl
sulfoxide (DMSO) (Sigma-Aldrich, St. Louis, MO, USA)에
녹여사용하였고, 세포처리전에배지에희석하여처리하였다.
세포 독성 시험
RAW 264.7 cell을 96-well plate에 5×105 cells/well로분주 하고 37C에서 24시간동안배양한후, EHF가 0, 10, 25, 50 µg/
mL 농도로희석된 DMEM (Life Technology, Carlsbad, CA,
USA) 배지로교체하여 1시간처리한후 LPS (Sigma-Aldrich,
St. Louis, MO, USA) (1 µg/mL)를함유한 DMEM 배지에다 시 24시간배양하였다. 이후 CellTiter96ⓇAqueous One
Solu-tion Cell ProliferaSolu-tion Assay [3-(4,5-dimethylthiazol-2-yl)- 5-(3-carboxymethoxyphenyl)-2-(4-sulfophenyl)-2H-tetrazo-lium; MTS] 키트(Promega, Madison, WI, USA)를사용하여
제조사의방법에따라세포생존율을분석하였다. MTS 용액은
FBS-free DMEM (Life Technology)에 5% (v/v)의농도로섞 어 100 µL씩처리하였다. 1시간후에 microplate reader
(Glo-max Multi Detection System, Promega, Madison, WI, USA)
를이용하여 490 nm의파장에서흡광도를측정하여세포독성 을분석하였다.
NO 및 염증성 cytokines 생성 억제 효과
EHF의항염증효과비교를위해 RAW 264.7 세포에서의 NO 생성에대한억제효과를분석하였다. EHF를 1시간동안전처 리한다음 LPS (1 µg/mL)로 24시간동안자극하고, 원심분리 (2,000 g, 4°C, 10분)하여배지를회수하였다. NO의농도는배 지(100 µL)와 Griess 시약 (0.1% naphthylethylene diaminedi-hydrochloride+1% sulfanilamide+5% phosphoric acid)을동 일한비율로반응시켜 microplate reader로 540 nm의파장에 서흡광도를측정하였다. 배지중의 IL-1β, IL-6, TNF-α의양 은 enzyme-linked immunosorbent assay (ELISA) (R&D
Sys-tems, Minneapolis, MN, USA) kit를이용하여제조사의방법
에따라측정하였다.
Reverse transcription-polymerase chain reaction
(RT-PCR)
RAW 264.7 세포(1×106 cells/well)에 EHF를 0, 10, 25, 50
로 6시간동안자극시켰다. 이후 Quiazol 시약 (Quiagen
Sci-ence, Valencia, CA, USA)을 이용하여 total RNA를 분리하 였다(Kim et al., 2009). Total RNA로부터 RT-PCR 분석에의 한 mRNA 발현양의분석은이전의보고(Joung et al., 2012)에
서사용한방법을이용하였고, 유전자발현양의상대적인비교
를위해서 β-actin 또는 glyceraldehyde-3-phosphate
dehydro-genase (GAPDH)를함께분석하였다. PCR 반응에이용된각 각의 primer는 Table 1에나타내었다. 전기영동상 band의정 량분석은 cooled CCD camera system EZ-Capture II (ATTO
& Rise Co., Tokyo, Japan)와 CS analyzer ver. 3.00 software
(ATTO and Rise Co., Tokyo, Japan)를이용하여최소 3번의반
복실험을통해얻었다.
세포질 및 핵 단백질 추출물의 제조
NF-κB의활성화정도를분석하기위해 EHF 및 LPS를각 각처리한 RAW 264.7 세포로부터세포질단백질과 핵단백 질을각각분리제조하였다(Kim et al., 2009). 즉, EHF로처 리된 세포(2×106 cells/dish)를 PBS로 세척하여 회수하고,
180 µL의 hypotonic buffer [10 mM Tris-HCl, 10 mM NaCl,
3 mM MgCl2, 0.02% NaN3, 0.5 mM dithiothreitol (DTT), 1 mM phenylmethanesulfonyl fluoride (PMSF), pH 7.4]를넣 고, 20 µL의 5% nonidet NP-40을첨가하여 5분동안방치하 였다. 이후원심분리(1,800 g, 4°C, 5분)한후상층액을세포질 추출물로이용하였다. 침전물은 hypotonic buffer로한번세척 하고, hypertonic buffer [20 mM
4-(2-hydroxyethyl)-1-piper-azineethanesulfonic acid, 25% glycerol, 420 mM NaCl, 1.5 mM MgCl2, 0.2 mM ethylenediaminetetraacetic acid, 0.02% NaN3, 0.5 mM DTT, 1 mM PMSF, pH 7.4]를넣고 1시간동안 얼음위에방치시킨다음원심분리(13,000 g, 4°C, 10분)하여상 층액을회수하여핵단백질추출물로이용하였다.
Western blot에 의한 단백질 분석
염증관련 단백질과신호전달 단백질의인산화정도는세포 를 EHF로전처리한후 LPS로처리하여세포단백질을시료 로, NF-κB 및 IκB의활성화및인산화정도는상기의핵및 세포질추출물을 시료로이용하였고, 단백질양은이전의보 고(Joung et al., 2017)와마찬가지로전기영동으로분리된단 백질을 nitrocellulose 막에이전시켜 Western blot으로분석하 였다. 검출된 band의정량분석은 mRNA 분석과마찬가지로cooled CCD camera system EZ-Capture II (ATTO and Rise Co., Seoul, Korea)와 CS analyzer ver. 3.00 software (ATTO
and Rise Co., Seoul, Korea)를이용하여최소 3번의반복실험 을통해얻었고, 그결과를각 blot의하단에수치로표기하였다.
면역형광분석
RAW 264.7 세포를 glass coverslips (SPL Lifesciences Co.,
Pocheon, Korea) 위에 24시간배양한뒤, EHF로 1시간전처리 하고, LPS (1 µg/mL)로 30분자극시켰다. 세포를 4.0%
para-formaldehyde가 첨가된 PBS로 실온에서 15분 동안반응시 켜고정시키고, 0.5% Triton X-100이첨가된 PBS를넣어 10 분동안반응시켰다. PBS로세척한뒤에 3% BSA/PBS를넣 고 30분 동안 blocking시킨후, NF-κB polyclonal
anti-body (Cell Signaling Technology, Danvers, MA, USA)가희 석된 3% BSA/PBS를넣어 2시간동안반응시켰다. 그다음, Table 1. Primer sequences for RT-PCR
Primers Sequences
iNOS Forward 5'-TCTTTGACGCTCGGAACTGT-3‘ Reverse 5'-CCATGATGGTCACATTCTGC-3‘ COX-2 Forward 5'-TGGGCAAAGAATGCAAACAT-3‘ Reverse 5'-CAGCAAATCCTTGCTGTTCC-3’ TNF-α Forward 5'-CAAGGGACAAGGCTGCCCCG-3'
Reverse 5'-GGTCAGAGTGGGGGCTGGGT-3‘ IL-1β Forward 5'-ATGGCAACTGTTCCTGAACTCAACT-3'
Reverse 5'-CAGGACAGGTATAGATTCTTTCCTTT-3' IL-6 Forward 5'-GTATGAACAACGATGATGCACTTCCAG-3'
Reverse 5'-GCATTGGAAATTGGGTAGGAAGG-3' β-Actin Forward 5'-CCTCATGAAGATCCTGACCG-3'
Reverse 5'-TCCACATCTGCTGGAAGGTG-3’ GAPDH Forward 5'-TGGCACAGTCAAGGCTGAGA-3' Reverse 5'-CTTCTGAGTGGCAGTGATGG-3’
RT-PCR, reverse transcription-polymerase chain reaction; iNOS, inducible nitric oxide synthase; COX-2, cyclooxygenase-2; TNF-α, tumor necrosis factor- α; IL-1β, interleukin-1β; IL-6, interleukin-6; GAPDH, glyceraldehyde-3-phosphate dehydrogenase.
Alexa FluorⓇ 488-conjugated secondary antibody (Invitrogen,
Carlsbad, CA, USA)가희석된 3% BSA/PBS를넣고 1시간 동안반응시킨뒤, 2 µg/mL의 4,6-diamidino-2-phenylindole
(DAPI) (Sigma-Aldrich, St. Louis, MO, USA)로핵을염색하 고 LSM700 laser scanning confocal microscope (Carl Zeiss,
Oberkochen, Germany)로관찰하였다.
NF-κB Promoter 활성
RAW 264.7 세포(2×105 cells/well)가 들어있는 24-well
plate의각 well에 1 µg의 pNF-κB firefly luciferase DNA와 20
ng의 pRL-TK renilla luciferase DNA를 Lipofectamine/Plus
reagent (Invitrogen, Carlsbad, CA, USA)와함께처리하여 40 시간동안 transfection시켰다. 그다음, EHF를 1시간전처리하 고, LPS (1 µg/mL)로 6시간자극시켰다. 이후 PBS로세척하 고 100 µL의 lysis buffer (0.5 mM HEPES, pH 7.8, 1% Triton
N-101, 1 mM CaCl2, and 1 mM MgCl2)로 lysate를만들고,
luciferase assay kit (Promega, Madison, WI, USA)를사용하 여 luciferase 활성을측정하였다.
귀 부종 억제 효과
동물시험방법과과정은부경대학교동물윤리위원회의승인 을받아수행하였다. ICR 생쥐(수컷, 25-30 g)는샘타코바이오 코리아(Osan, Korea)에서구입하였다. 구입된생쥐들은 1주일 간동물사육실에적응시킨후그룹별로 6 마리씩할당하였다.귀 부종은 phorbol 12-myristate 13-actate (PMA)
(Sigma-Aldrich, St. Louis, MO, USA)를 0.2 μg/mL 농도로아세톤에
녹여사용하여유도하였다. 대조군은생리식염수로처리하고,
PMA 처리군, PMA+0.5 mg EHF 처리군, PMA+1.0 mg EHF 처리군및 PMA+indomethacin (Indo) 처리군으로나누었다. 왼쪽귀(reference)는 30 µL의 아세톤으로 처리하였다. EHF (0.5 mg, 1.0 mg)와 Indo (1 mg)는 30 µL의아세톤에녹여오 른쪽귀안쪽에처리하고, 1시간후에동일부위에 PMA 6 µg을 30 µL의아세톤에녹여처리하였다. PMA 처리 6시간후에생 쥐를희생시켜 6 mm 구경의금속펀쳐로부종부위를절취하여 무게를재었다. 귀부종무게는오른쪽귀의무게에서왼쪽귀무
게를뺀값으로하였다. 저해퍼센트(inhibition percentage, IP) 는 PMA 처리군에대한상대적인무게를나타낸값이다.
통계 처리
본연구의모든실험은세번이상반복하였으며, 얻어진결
과들을평균값과표준편차(mean±SD)를계산하여나타내었 다. 실험군간의유의성검증은 Student’s t-test로검증하였다. Fig. 1. Effect of EHF on cell viability, nitric oxide (NO) and prostaglandin E2 (PGE2) production in LPS-stimulated RAW 264.7 cells. (A) Cell viabilities were measured with MTS assay. Cells pretreated with various concentrations of EHF for 1 h were stimulated with LPS (1 μg/mL) for 24 h. (B) The culture media of the treated cells were used to measure the amount of nitrite to evaluate NO level, or (C) the amount of PGE2. All data are presented as mean±SD of three independent experiments. #P<0.001 compared to non-treated group. *P<0.05
and **P<0.01 compared to LPS-only group.
(A) (B)
(C)
(A) (B)
결 과
EHF에 의한 NO와 PGE
2생성 억제 효과
RAW 264.7 세포에대한 EHF의세포독성을측정하기위하 여 0-50 µg/mL 농도범위로 EHF를 24시간처리한다음세포생 존율의변화를 MTS assay로분석하였다. Fig. 1A에나타난바 와같이세포의생존율은 EHF 처리에의해뚜렷한변화가없었 다. 따라서이후의실험은 EHF를 50 µg/mL 이하의농도에서 행하였다. LPS로자극된세포에서생성되는 NO에대한 EHF 의억제효과를확인하기위하여세포를 EHF (0-50 µg/mL)로 1시간전처리한후 LPS로 24시간자극하여배지에방출된 ni-trite 생성량을측정하여 NO 생성량을분석하였다. LPS 처리에 의해증가된 NO는 EHF의전처리에의해농도의존적으로현 저하게감소하는것으로나타났고, 특히 25 µg/mL 이상의농 도처리군에서는 50% 이상의감소효과가나타났다(Fig. 1B). EHF에의한 PGE2생성억제효과또한 NO 생성억제효과와유 사한결과를나타내었다(Fig.1C).
EHF에 의한 iNOS와 COX-2 발현 억제 효과
NO는 iNOS에의해그리고 PGE2는 COX-2에의해생성되므 로, 거식세포에 EHF를전처리한다음 LPS로처리후에세포단 백질을분리하여 iNOS와 COX-2 단백질발현수준을 Western
blot으로분석하였다. LPS 처리에의해생성된 iNOS 단백질 은 EHF 처리에의해농도의존적으로감소하는경향을나타내 었다(Fig. 2). COX-2 단백질역시 EHF에의해감소하는경향 을보였다(Fig. 2). iNOS와 COX-2 단백질발현이전사수준에 서조절되는것을확인하기위하여 iNOS와 COX-2 mRNA 발 현을 RT-PCR로분석하였다. 단백질과마찬가지로두유전자의
mRNA 발현양상이유사하게나타났고, 25 µg/mL 이상의농
도처리군에서는감소경향이보다현저하였다(Fig. 2). 따라서
EHF에의한 iNOS와 COX-2 단백질의발현감소는전사수준
에서효과적으로조절되고있음을보여주었다.
LPS로 유도되는 염증성 cytokines의 생성에 대한
EHF의 억제효과
LPS로자극된 RAW 2645.7 세포에서생성되는염증촉진성
cytokines의생성에대한 EHF의효과를 ELISA 방법으로분석 하였다. LPS 자극에의해 TNF-α (Fig. 3A), IL-1β (Fig. 3B) 및
IL-6 (Fig. 3C)와같은염증촉진성 cytokines의생성량은크게
증가하는것으로나타났고, 이러한증가는다소차이는있지만
EHF 처리에의해농도-의존적으로현저하게감소하는것으로
관찰되었다(Fig. 3A, 3B, 3C). 또한처리된세포로부터 mRNA 를분리하여 RT-PCR로분석한결과 ELISA 결과와유사하게 나타났다(Fig. 3D).
LPS로 유도되는 NF-κB의 활성화에 대한 EHF의 억
제 효과
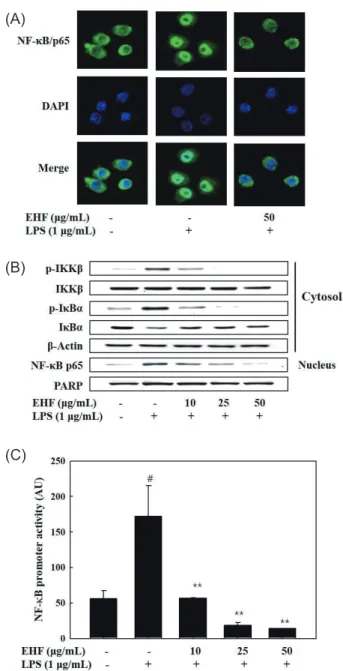
염증촉진성 cytokines, iNOS 및 COX-2의발현은전사인자 인 NF-κB에의해조절된다. LPS로자극된 RAW 264.7 세포를 이용하여 NF-κB의활성화에대한 EHF의효과를분석하였다. 먼저면역형광법으로염색을하고 confocal microscopy로분 석한결과를 Fig. 4A에나타내었다. 자극이가해지지않은상태 에서 NF-κB/p65 subunit (녹색)는주로세포질에분포하는것 이관찰되었다. LPS로처리된세포의경우녹색인 NF-κB p65 의대부분은핵에분포하는것으로나타났고, 이는 NF-κB가 활성화되어핵으로이동했음을보여주는결과이다. 그러나세 포에 EHF (50 µg/mL)를전처리한경우 NF-κB/p65는핵주변 의세포질에대부분분포하는것으로관찰되었다. 이러한결과 는 EHF에의해 NF-κB의활성화가현저하게억제되고있음을 보여주고있다(Fig. 4A). NF-κB의활성화에영향을미치는단 백질을 Western blot으로분석한결과를 Fig. 4B에나타내었다.
LPS에의해과도하게인산화된 IKK-β와 IκBα는 EHF 처리에 의해인산화정도(p-IKK-β 및 p-IκBα)가농도의존적으로감 소함을보여주었다. LPS에의해핵으로이동된 NF-κB p65 또 한 EHF 처리에의해감소됨을보여주고있다. 다음은 LPS로 자극된대식세포주에있어 NF-κB의 promoter activity에대한
EHF의효과를분석하였다(Fig. 4C). 이를위해 RAW 264.7 세 포에 NF-B promoter를가진 luciferase construct를일시적으 Fig. 2. Effect of EHF on LPS-stimulated inducible nitric oxide syn-thase (iNOS) and cyclooxygenase-2 (COX-2) protein and mRNA expressions in RAW 264.7 cells. Cells pretreated with various con-centrations of EHF for 1 h were stimulated with or without LPS (1 μg/mL) for 16 h. Whole proteins were separated with SDS-PAGE. The expression of iNOS, COX-2, and β-actin was analyzed by Western blot using corresponding antibodies. Cells were incubated with various concentrations of EHF for 1 h, and then stimulated with LPS (1 μg/mL) for 6 h. Total RNA was prepared for RT-PCR. The results presented are representatives of three independent ex-periments.
(A)
(B)
대황 헥산 분획물의 항염증 효과
157
로 transfection하고, 이세포를다양한농도의 EHF로 2시간전 처리하고이어서 LPS로 6시간동안자극하였다. Fig. 4C에나 타내었듯이 luciferase 활성은 LPS 자극에의해현저하게증가 하였으며, 이는 LPS 자극에의해활성화된 NF-κB가 NF-κB 의 promoter를가진 luciferase의발현을크게증가시켰음을의 미한다. 이에대한 EHF의억제효과는 10 µg/mL의낮은농도 뿐만아니라 50 µg/mL 농도에이르기까지유의적으로감소하 는것으로나타났다. 이들결과는대식세포에서 LPS 자극에의 해유도되는 NF-κB의활성화가 EHF에의해효과적으로억제 되고있음을보여주고있고, 이는 EHF에의한상기의염증성 cytokines 및 iNOS의발현억제는부분적으로 NF-κB 활성화 경로에의해조절되고있음을의미하고있다.LPS로 유도되는 MAPKs의 인산화에 대한 EHF의 억
제 효과
EHF에의한 NF-κB 신호경로에어떠한신호단백질들이관여 하는지를확인하기위하여 MAPKs (ERK, JNK, p38 MAPK) 와 Akt의인산화를 Western blot으로분석하였다. Fig. 5에나타 내었듯이, LPS로유도된세포에서 EHF 처리에의하여 ERK, JNK, p38 MAPK 및 Akt의인산화가농도의존적으로감소하 였다. 이러한결과는 EHF가 IKK-β의인산화는물론 NF-κB의 상위단백질의인산화를조절함으로써염증억제효과를나타낸 다는것을의미한다.부종에 대한 EHF 효과
생쥐의귀에 PMA를처리하여귀부종을유도한결과 6시간 후 PMA에의해귀부종의무게가현저히증가하였다(Fig. 6).Indo는비스테로이드성항염증제(nonsteroidal
anti-inflamma-tory drug, NSAID)로써부종의치료효능을평가하기위하여 양성대조약물로사용하였다. Fig. 6에나타난바와같이부종 은 EHF 처리에의해감소하는경향이나타났고, 특히 1 mg의 EHF 처리에의해유의적인차이가확인되었다(P<0.05). 이러 한결과에서 EHF의부종억제효과를확인할수있었다.
고 찰
NO는 NOS에 의해 L-arginine으로부터 생성되며, 세균의endotoxin 또는염증성 cytokines에 iNOS가급격하게유도되 어 NO 생성량이급증한다(Guha and Mackman, 2001). 병리적 인조건하에서 iNOS에의한 NO의현저한증가는다른염증성 Fig. 3. Effect of EHF on the production of pro-inflammatory cytokines in LPS-stimulated RAW 264.7 cells. Cells pretreated with various concentrations of EHF were stimulated with or without LPS (1 μg/mL) for 24 h. TNF-α (A), IL-1β (B), and IL-6 (C) in the culture media were measured by ELISA. (D) Cells were incubated with various concentrations of EHF for 1 h, and then stimulated with LPS (1 μg/mL) for 6 h. Total RNA was prepared for RT-PCR. The results presented are representatives of three independent experiments. Data represent mean±SD of three independent experiments. #P<0.001 compared to non-treated group. **P<0.01 compared to LPS-only group.
(C)
(A) (B)
김보운
ㆍ
최창근ㆍ
김재일ㆍ
김형락158
매개체들과함께과도한염증을유발하게되고조직의손상을
유발하는것으로알려져있어염증성손상의주요매개체이다
(Pan et al., 2011). PGE2는 COX-2의작용에의해 arachidonic
acid로부터생성된다(Sil and Ghosh, 2016). 체내의염증작용 이 증가함으로써생성되는 PGE2는통증, 염증, 체온상승, 혈 소판응집등의작용을한다(Park et al., 2006). 따라서 iNOS 와 COX-2의발현을억제하거나활성을억제함으로써 NO와
PGE2의생성을억제할수있는화합물은항염증물질로이용 Fig. 4. Inhibitory effect of EHF on the activation of NF-κB in
LPS-stimulated RAW 264.7 cells. A, celluar distribution of p65 subunit of NF-κB (green) protein was analyzed by immunofluorescence staining with confocal microscopy. Cells were pretreated with EHF (50 μg/mL) for 1 h, followed by LPS stimulation for 30 min. DAPI (blue) was used for nuclear staining; B, nuclear localization of NF-κB and the regulation of INF-κB-α phosphorylation was analyzed by Western blot. Cells pretreated with EHF for 1 h were stimulated with LPS for 30 min; C, effect of EHF on NF-κB promoter activity. Cells were transfected with 1 μg of NF-κB promoter-containing luciferase DNA for 40 h. Transfected cells pretreated with EHF for 1 h were stimulated with LPS for additional 6 h. The results pre-sented are representatives of three independent experiments; AU, arbitrary unit. #P<0.001 compared to non-treated group. **P<0.01
compared to LPS-only group.
(A) (B) (C) (A) (B) (C)
Fig. 6. Effect of EHF on ear edema induced by phorbol 12-my-ristate 13-actate (PMA) in mice. EHF or indomethacin (Indo) were topically administered on inner surface of the right ears of mice 1 h before application of PMA. 6 h after application of PMA, ear edema and the inhibition percentage (IP) of the ear edema by the treatment were calculated. #P<0.001 compared to non-treated
group. **P<0.05 compared to PMA-only treated group.
Fig. 5. Effect of EHF on the phosphorylations of MAPKs and Akt in LPS-stimulated RAW 264.7 cells. Cells were incubated with various concentrations of EHF for 1 h, and then stimulated with LPS (1 μg/mL) for 30 min. Whole cell lysates were prepared and analyzed by Western blot for phosphorylated proteins of Akt, JNK, p38 MAPK or β-actin using corresponding antibodies. The results presented are representatives of three independent experiments.
(B)
될수있다. 염증촉진성 cytokines들은체내에서다양한면역및염증반 응을조절하는역할을한다. 세균의 LPS에의해자극된대식 세포는 TNF-α를생성하고분비된 TNF-α는 IL-1와 IL-6의생 성을유도함으로써염증반응을증폭시키게된다(Minnich and Moldawer, 2004). LPS에의해유도된 TNF-α는염증반응의개 시를촉진하며지속적인생성은만성염증을유발하며결국에 는패혈성쇼크, 염증등의다양한생리적과정에관여하고있다
(Scheller et al., 2011). IL-1β는대식세포에서생성되는주요염 증촉진성 cytokine으로, 세균감염에대한염증응답의개시및 강화에중요한 cytokine이다(Li and Verma, 2002). IL-6도대식 세포에서생성되는중요한염증성 cytokine으로서급성면역반 응에작용한다(Bonizzi and Karin, 2004). 본연구에서관찰된 결과는 EHF가 LPS-자극에의해유도되는 IL-1β, IL-6, TNF-의생성을억제시키는것으로나타났고, 이는 EHF가 LPS-자극
에의한염증성응답의초기단계를억제하고있음을의미한다.
NF-κB의활성화를억제하는단백질인 IκBα는 IKKβ의인산 화에의하여 IκBα가인산화됨으로써유비퀴틴화되어프로테 아좀에의해분해된다(Nguyen et al., 2003). 따라서 IκB-α로 부터유리된 NF-κB는핵으로이동하여염증성유전자들의발 현에기여하게된다. 또한 NF-κB 활성화경로는세포내다양 한신호전달에관여하는 MAPKs나 Akt와같은 protein kinase 들에의해조절된다(Liu et al., 2017; Tian et al., 2017). 많은연 구들에서 p38, JNK, ERK와같은 MAPKs가설치류의대식세 포에서 LPS에인한 NF-κB의활성화에중요한역할을한다고 밝히고있다(Guha and Mackman, 2001; Joung et al., 2017). 그리고 Akt 또한 NF-κB의활성화를조절함으로써여러염증 성유전자의발현을조절하는것으로알려져있다(Joung et al.,
2015). 본연구에서 LPS 자극에의해유도되는 NF-κB의활성 화에대한 EHF의억제효과는 MAPKs 및 Akt와같은 kinase의
인산화과정과관련되어있음을나타낸다.
앞서설명한것처럼모자반류에서분리된 sargaquinoic acid
(Gwon et al., 2015; Joung et al., 2015), fucosterol (Jung et al., 2013; Gwon et al., 2017), sargachromenol (Gwon et al., 2017)
과같은화합물들의항염증활성에대해활발한연구가진행되
었다. 본실험에사용된 EHF를 HPLC로분석한결과
sargahy-droquinoic acid, sargaquinoic acid 및 sargachromenol은발견
되지않았다. 따라서이들항염증활성을나타내는유효성분을 분리동정하기위한후속적인연구가필요하다. 이상의결과에서, LPS로자극한 RAW 264.7 세포에있어 NO 와같은염증성매개체뿐만아니라염증촉진성 cytokines의생 성및발현이 EHF에의해억제된다는것을증명하였다. 이러 한 EHF의억제효과는 IκB의분해를저해함으로써 NF-κB 활 성화경로를억제시키는것으로나타났다. 또한 EHF는 NF-κB 활성화의상위신호전달경로인 MAPKs 및 Akt에도영향을미 치는것으로확인되었다. 여러만성염증성질환에과도한염증 성매개체들의발현및생성이중요한병인적역할을하고있 음을고려했을때, 본연구결과는 EHF가항염증효능을가진 기능성식품의소재로이용될수있다는가능성을제시하고있 다. 향후 EHF의항염증효과를나타내는유효화합물의분리.동 정및활성분석에대한연구가진행되어야할것으로판단된다.
사 사
이논문은부경대학교자율창의학술연구비(2019년)에의하 여연구되었음.References
Bonizzi G and Karin M. 2004. The two NF-kappaB activa-tion pathways and their role in innate and adaptive immu-nity. Trends Immunol 25, 280-288. https://doi.org/10.1016/j. it.2004.03.008.
Choi JS, Han YR, Byeon JS, Choung SY, Sohn HS and Jung HA. 2015a. Protective effect of fucosterol isolated from the edible brown algae, Ecklonia stolonifera and Eisenia
bicyclis, on tert-butyl hydroperoxide- and tacrine-induced
HepG2 cell injury. J Pharm Pharmacol 67, 1170-1178. https://doi.org/10.1111/jphp.12404.
Choi JS, Haulader S, Karki S, Jung HJ, Kim HR and Jung HA. 2015b. Acetyl- and butyryl-cholinesterase inhibitory activi-ties of the edible brown alga Eisenia bicyclis. Arch Pharm Res 38, 1477-1487. https://doi.org/10.1007/s12272-014-0515-1.
D'Acquisto F, Iuvone T, Rombola L, Sautebin L, Di Rosa M and Carnuccio R. 1997. Involvement of NF-kappaB in the regulation of cyclooxygenase-2 protein expression in LPS-stimulated J774 macrophages. FEBS Lett 418, 175-178. https://doi.org/10.1016/s0014-5793(97)01377-x.
Elliott PJ, Zollner TM and Boehncke WH. 2003. Proteasome inhibition: a new anti-inflammatory strategy. J Mol Med 81, 235-245. http://dx.doi.org/10.1007/s00109-003-0422-2. Eom SH, Lee DS, Jung YJ, Park JH, Choi JI, Yim MJ, Jeon
JM, Kim HW, Son KT, Je JY, Lee MS and Kim YM. 2014. The mechanism of antibacterial activity of phlorofucofu-roeckol-A against methicillin-resistant Staphylococcus
au-reus. Appl Microbiol Biotechnol 98, 9795-9804. https://doi.
org/10.1007/s00253-014-6041-8.
Guha M and Mackman N. 2001. LPS induction of gene expres-sion in human monocytes. Cell Signal 13, 85-94. https://doi. org/10.1016/s0898-6568(00)00149-2.
Gwon WG, Joung EJ, Kwon MS, Lim SJ, Utsuki T and Kim HR. 2017. Sargachromenol protects against vascular in-flammation by preventing TNF-alpha-induced monocyte adhesion to primary endothelial cells via inhibition of NF-kappaB activation. Int Immunopharmacol 42, 81-89. https:// doi.org/10.1016/j.intimp.2016.11.014.
Gwon WG, Lee B, Joung EJ, Choi MW, Yoon N, Shin T, Oh CW and Kim HR. 2015. Sargaquinoic acid inhibits TNF-alpha-induced NF-kappaB signaling, thereby contributing to decreased monocyte adhesion to human umbilical vein endothelial Cells (HUVECs). J Agric Food Chem 63, 9053-9061. https://doi.org/10.1021/acs.jafc.5b04050.
Joung EJ, Gwon WG, Shin T, Jung BM, Choi J and Kim HR. 2017. Anti-inflammatory action of the ethanolic extract from Sargassum serratifolium on lipopolysaccharide-stim-ulated mouse peritoneal macrophages and identification of active components. J Appl Phycol 29, 563-573. https://doi. org/10.1007/s10811-016-0954-9.
Joung EJ, Lee B, Gwon WG, Shin T, Jung BM, Yoon NY, Choi JS, Oh CW and Kim HR. 2015. Sargaquinoic acid attenu-ates inflammatory responses by regulating NF-kappaB and Nrf2 pathways in lipopolysaccharide-stimulated RAW 264.7 cells. Int Immunopharmacol 29, 693-700. https://doi. org/10.1016/j.intimp.2015.09.007.
Joung EJ, Lee MS, Choi JW, Kim JS, Shin T, Jung BM Kim JI and Kim HR. 2012. Anti-inflammatory effects of phlo-rofucofuroeckol B-rich ethyl acetate fraction obtained from
Myagropsis myagroides on lipopolysaccharide-stimulated
RAW 264.7 cells and mouse edema. Int Immunopharmacol 14, 471-480. https://doi.org/10.1016/j.intimp.2012.08.021. Jung HA, Jin SE, Ahn BR, Lee CM and Choi JS. 2013.
Anti-inflammatory activity of edible brown alga Eisenia bicyclis and its constituents fucosterol and phlorotannins in LPS-stimulated RAW264.7 macrophages. Food Chem Toxicol 59, 199-206. https://doi.org/10.1016/j.fct.2013.05.061. Kaminska B. 2005. MAPK signaling pathways as molecular
tar-gets for anti-inflammatory therapy - from molecular mecha-nisms to therapeutic benefits. Biochim Biophys Acta 1754, 253-262. https://doi.org/10.1016/j.bbapap.2005.08.017. Kim AR, Lee MS, Shin TS, Hua H, Jang BC, Choi JS, Byun
DS, Utsuki T, Ingram D and Kim HR. 2011. Phlorofucofu-roeckol A inhibits the LPS-stimulated iNOS and COX-2 ex-pressions in macrophages via inhibition of NF-kappaB, Akt, and p38 MAPK. Toxicol In Vitro 25, 1789-1795. https://doi. org/10.1016/j.tiv.2011.09.012.
Kim AR, Shin TS, Lee MS, Park JY, Park KE, Yoon NY, Kim JS, Choi JS, Jang BC, Byun DS, Park NK and Kim HR. 2009. Isolation and identification of phlorotannins from
Ecklonia stolonifera with antioxidant and anti-inflammatory
properties. J Agric Food Chem 57, 3483-3489. https://doi. org/10.1021/jf900820x.
Kim HJ, Dasagrandhi C, Kim SH, Kim BG, Eom SH and Kim YM. 2018. In vitro antibacterial activity of phlorotannins from edible brown algae, Eisenia bicyclis against strepto-mycin-resistant Listeria monocytogenes. Indian J Microbiol 58, 105-108. http://dx.doi.org/10.1007/s12088-017-0693-x. Li Q and Verma IM. 2002. NF-kappaB regulation in the
im-mune system. Nat Rev Immunol 2, 725-734. https://doi.
org/10.1038/nri910.
Libby P and Hansson GK. 2015. Inflammation and immunity in diseases of the arterial tree: players and layers. Circ Res 116, 307-311. https://doi.org/10.1161/circresaha.116.301313. Liu Q, Hu Y, Cao Y, Song G, Liu Z and Liu X. 2017.
Chico-ric acid ameliorates lipopolysaccharide-induced oxidative stress via promoting the Keap1/Nrf2 transcriptional signal-ing pathway in BV2 microglial cells and mouse brain. J Agric Food Chem 65, 338-347. https://doi.org/10.1021/acs. jafc.6b04873.
Marks-Konczalik J, Chu SC and Moss J. 1998. Cytokine-mediated transcriptional induction of the human inducible nitric oxide synthase gene requires both activator protein 1 and nuclear factor kappaB-binding sites. J Biol Chem 273, 22201-22208. https://doi.org/10.1074/jbc.273.35.22201. Minnich DJ and Moldawer LL. 2004. Anti-cytokine and
anti-inflammatory therapies for the treatment of severe sepsis: progress and pitfalls. Proc Nutr Soc 63, 437-341. https://doi. org/10.1079/pns2004378.
Nguyen T, Sherratt PJ, Huang HC, Yang CS and Pickett CB. 2003. Increased protein stability as a mechanism that en-hances Nrf2-mediated transcriptional activation of the an-tioxidant response element. Degradation of Nrf2 by the 26 S proteasome. J Biol Chem 278, 4536-4541. https://doi. org/10.1074/jbc.m207293200.
Pan MH, Hong HM, Lin CL, Jhang AZ, Tsai JH, Badmaev V, Nagabhushanam K, Ho CT and Chen W J. 2011. Se-meth-ylselenocysteine inhibits lipopolysaccharide-induced NF-kappaB activation and iNOS induction in RAW 264.7 mu-rine macrophages. Mol Nutri Food Res 55, 723-732. https:// doi.org/10.1002/mnfr.201000481.
Park JY, Pillinger MH and Abramson SB. 2006. Prostaglan-din E2 synthesis and secretion: the role of PGE2 synthases.
Clin Immunol 119, 229-240. https://doi.org/10.1016/j. clim.2006.01.016.
Paudel U, Lee YH, Kwon TH, Park NH, Yun BS, Hwang PH and Yi HK. 2014. Eckols reduce dental pulp inflamma-tion through the ERK1/2 pathway independent of COX-2 inhibition. Oral Dis 20, 827-832. https://doi.org/10.1111/ odi.12266.
Rahman I, Biswas SK and Kirkham PA. 2006. Regulation of in-flammation and redox signaling by dietary polyphenols. Bio-chem Pharmacol 72, 1439-1452. https://doi.org/10.1016/j. bcp.2006.07.004.
Scheller J, Chalaris A, Schmidt-Arras D and Rose-John S. 2011. The pro- and anti-inflammatory properties of the cytokine interleukin-6. Biochim Biophys Acta 1813, 878-888. https:// doi.org/10.1016/j.bbamcr.2011.01.034.
Sil S and Ghosh T. 2016. Role of cox-2 mediated neuroinflam-mation on the neurodegeneration and cognitive impairments in colchicine induced rat model of Alzheimer's Disease. J Neuroimmunol 291, 115-124. https://doi.org/10.1016/j.
jneuroim.2015.12.003.
Sugiura Y, Matsuda K, Okamoto T, Yamada Y, Imai K, Ito T, Kakinuma M and Amano H. 2009. The inhibitory effects of components from a brown alga, Eisenia arborea, on de-granulation of mast cells and eicosanoid synthesis. J Funct Foods 1, 387-393. https://doi.org/10.1016/j.jff.2009.08.002. Thomas NV and Kim SK. 2011. Potential pharmacological ap-plications of polyphenolic derivatives from marine brown algae. Environ Toxicol Pharmacol 32, 325-335. https://doi. org/10.1016/j.etap.2011.09.004.
Tian Y, Li Z, Shen B, Zhang Q and Feng H. 2017. Protective effects of morin on lipopolysaccharide/d-galactosamine-induced acute liver injury by inhibiting TLR4/NF-kappaB and activating Nrf2/HO-1 signaling pathways. Int Immu-nopharmacol 45, 148-155. https://doi.org/10.1016/j.in-timp.2017.02.010.
Xie QW, Kashiwabara Y and Nathan C. 1994. Role of transcrip-tion factor NF-kappa B/Rel in inductranscrip-tion of nitric oxide syn-thase. J Biol Chem 269, 4705-4708.
Yayeh T, Im EJ, Kwon TH, Roh SS, Kim S, Kim JH, Hong SB, Cho JY, Park NH and Rhee MH. 2014. Hemeoxygenase 1 partly mediates the anti-inflammatory effect of dieckol in lipopolysaccharide stimulated murine macrophages. Int Immunopharmacol 22, 51-58. https://doi.org/10.1016/j.in-timp.2014.06.009.
Zhang G and Ghosh S. 2000. Molecular mechanisms of NF-kappaB activation induced by bacterial lipopolysaccharide through Toll-like receptors. J Endotoxin Res 6, 453-457. https://doi.org/10.1179/096805100101532414.