Original Article
戊己丸의 RAW 264.7 세포에 대한 항염증작용 연구
김일현⋅최종환⋅이세원⋅송용선
원광대학교 한의과대학 한방재활의학과교실
Anti-inflammatory Effect of Mugi-hwan Water Extract in RAW 264.7 Cells
Ilhyun Kim, O.M.D., Chonghwan Choi, M.D., Sewon Lee, O.M.D., Yungsun Song, O.M.D.
Department of Oriental Rehabilitation Medicine, College of Oriental Medicine, Won-Kwang University
RECEIVED June 25, 2013 REVISED July 7, 2013 ACCEPTED July 11, 2013
CORRESPONDING TO Yungsun Song, Department of Oriental Rehabilitation Medicine, College of Oriental Medicine, Won-Kwang University, 142-1, Deokjin-dong 2ga, Deokjin-gu, Jeonju 561-851, Korea
TEL (063) 270-1070 FAX (063) 270-1199 E-mail [email protected]
Copyright © 2013 The Society of Korean Medicine Rehabilitation
Objectives The aim of this study was to investigate anti-inflammatory activity of Mugi-hwan (MGH) Water Extract.
Methods Cells were treated with 2 ug/ml of LPS 1 hour prior to the addition of MGH.
Cell viability was measured by MTS assay. The production of NO was determined by re- acting cultured medium with Griess reagent. The expression of COX-2, iNOS and MAPKs was investigated by Western blot, RT-PCR. The content of level of cytokines (PGE
2, IL-6, in media from LPS-stimulated Raw 264.7 cells was analyed by ELISA kit.
Results MGH inhibited the production of NO, PGE
2, IL-6 as well as the expressions of iNOS, COX-2 in the murine macrophage, RAW 264.7 cells. MGH also had suppression ef- fects of LPS induced MAPKs activation.
Conclusions These results suggest that MGH has an anti-inflammatory therapeutic po- tential, which may result from inhibition of MAPK phosphorylation, thereby decreasing the expression of pro-inflammatory genes. (J Korean Med Rehab 2013;23(3):27-35)
Key words Mugi-hwan , MAPKs, Anti-inflammatory activity
서론»»»
염증반응은 상처부위에 침입한 미생물들에 대한 적대 환경을 만드는 비특이적인 방어 작용이며, 인체 내 대부 분의 기관에서 유발되는 기본적인 면역반응으로 급성과 만성으로 나누어지는데1), 급성 염증반응은 물리적 자극이 나 외부 감염 등에 의해 수 분으로부터 수 시간 이내 즉 각적으로 조직손상을 유발하게 된다. 반면 만성 염증반응 은 급성보다 오래 걸리며 지속적이고, 단핵구나 대식세 포, 림프구, 혈장세포 등의 침윤을 동반하는 특징이 있으 며, 조직파괴와 치유과정을 통해 혈관신생이나 섬유화를 유발하게 된다1,2).
대식세포는 병원균과 같은 외부 자극원의 자극에 의해
활성화되어 vascular amines, arachidonic acid metabo- lites (prostaglandins, leukotriene, lipoxin 등), inflamma- tory cytokines (TNF-α, IL-1, IL-6, IL-8, IL-12, GM-CSF), platelet activating factor, prostaglandin E2, nitric oxide, ROS 등 다양한 염증매개물질들을 분비함으로써 면역조절 을 통해 생체 내 방어기능과 항상성 유지 기능을 조절하 는 반면, 염증을 유발하여 각종 폐질환이나 만성 기관지 염, 기관지 천식, 류마티스 관절염, 다발성 경화증, 퇴행 성 뇌질환, 동맥경화 등의 만성 염증질환으로의 진행에 기여한다3-6). 따라서 대식세포의 활성이 어떻게 조절되는 가를 이해하고, 활성화된 대식세포로부터 분비되는 각종 염증매개물질들을 억제시키는 것은 여러 원인에 의해 발 생하는 염증질환의 진행을 막아줄 수 있는 유효한 접근방
cDNA Primer sequence
iNOS forward reverse 5'-CATGTTCTCTGGGAAATCGTGG-3' 5'-AACGCACTAGGTTTGCCGAGTA-3' COX-2 forward reverse 5'-CACTCAGTTTGTTGAGTCATTC-3'
5'-GATTAGTACTGTAGGGTTAATG-3' β-actin forward reverse 5-ATGAAGATCCTGACCGAGCGT-3'
5'-AACGCAGCTCAGTAACAGTCCG-3' Table I. Primer Sequences for RT-PCR
Herbal name Pharmacognostic name Weight (g) 黃蓮 Rhizoma Coptidis 15.0 吳茱萸 Fructus Evodiae 15.0 白芍藥 Radix Paeoniae Alba 15.0 Table II. The Composition of Mugi-hwan
법이 될 수 있다6,7).
戊己丸 (
Mugi-hwan
, MGH)은 宋代 「太平惠民和劑局 方」8)에 처음 기록된 처방으로 黃連, 吳茱萸, 白芍藥으로 구성되어 있으며 「東醫寶鑑」9)에 濕痢를 치료하는데, 濕 痢란 腹脹身重하고 黑豆汁이나 赤黑渾濁한 설사를 하는 것이다.戊己丸에 대한 실험적 연구는 거의 없는 실정이나, 조 성 약재인 黃連, 吳茱萸, 白芍藥에 대한 연구는 비교적 많 이 이루어졌다. 黃連 추출물은
Helicobacter pylori
의 성 장을 억제하고10), LPS에 의해 유도된 nitric oxide의 생성 을 억제하며11) 黃連에 함유된 berberine은 항염증 효과를 가지는 것으로 알려져 있다12). 吳茱萸의 주요성분으로 알려진 evodiamine은 염증에 대한 효과가 보고되어있으 며13), 白芍藥 추출물은 항혈전 작용14), 항고지혈 작용15), 항산화 작용16), 혈관확장 작용17) 등이 보고되어 있다.이에 저자는 戊己丸의 항염증 효과를 조사하기 위하여 LPS로 자극된 설치류의 대식세포주인 RAW 264.7 세포에 서 세포가 방출하는 NO와 PGE2 생성량과 iNOS, COX-2 의 발현 및 IL-6에 미치는 영향을 알아보고 그 기전을 밝 히고자 실험하여 유의한 결과를 얻었기에 보고하는 바이 다.
재료 및 방법»»»
1. 재료
1) 시약
LPS (
E. coli
lipopolysaccaride), RPMI-1640 배지는 Sigma Chemical Co. (St. Luis, MO, USA)로부터 구입하였고, 우태아 혈청(Fetal bovine serum, FBS) 및 항생제 는 Gibco BRL Co. (Grand Island, NY, USA)로부터 구입 하였다. 조직배양 plate와 직경 100 mm petri-dish는 Nunc Inc. (Rochester, NY, USA)로부터 구입하여 사용하 였다. PGE2, IL-6은 효소결합면역측정(ELISA) 키트를 R&D Systems Inc. (Minneapolis, MN, USA)로부터 구입 하였으며, COX-2, iNOS, MAPKs (ERK, JNK, p38)는 Santa Cruz Biotechnology Inc. (Santacruz, CA, USA)에 서 구입하였다.
2) 약재
戊己丸의 구성 약재인 黃連, 吳茱萸, 白芍藥은 대학한 약국(익산, 한국)에서 구입하여 사용하였다.
3) Primer 준비
RT-PCR을 위해 여러 개의 primer를 사용하였다. Primer 의 구성은 다음과 같다(Table I).
2. 방법
1) 戊己丸 물 추출
戊己丸 물 추출물은 3차 증류수(100 g/L)로 2시간 환류 추출하여 준비하였다. 그 추출물은 0.45μm 필터로 걸렀 고, 감압하에 동결 건조(lyophilized)하였으며, 4oC에서 보관하였다. 동결건조된 추출물은 실험하기 위하여 인산 완충 식염수(phosphate-buffered saline. PBS)에 용해하 였다. 본 실험에 사용한 戊己丸은 「東醫寶鑑」9)처방을 기 준으로 하였으며, 黃蓮, 吳茱萸, 白芍藥 각 1:1:1의 비율 로 구성되어있으며(Table II) 총 45 g에서 D.W 500 ml씩 2회 4.94 g을 추출하여 수득률 10.97%이다.
2) 세포 배양
설치류 대식세포주(murine macrophage cell line, RAW 264.7)는 한국 세포주 은행에서 구입하여 사용하였
다. 대식세포주는 항생제 및 항균제(penicillin G 100 U/ml, streptomycin 100 U/ml)를 첨가하고 10% 열처리 우태아 혈청(heat inactivated FBS)을 첨가한 완전한 RPMI 1640 배지에서 5% CO2의 습한 대기, 37oC의 온도 조건에서 배양하였다.
3) NO 생성량 측정
DMSO로 녹인 PA를 여러 농도(1~100μM)로 처리하고 여기에 LPS (200 ng/ml)를 각각 주입한 다음 상기 대식 세포주 RAW 264.7을 24시간 배양하였다. 세포배양액의 최종 DMSO 농도는 0.1% 이하였다. 배양한 대식세포주의 상층액을 수집하여 Griess reagent (1% sulfanilamide, 0.1% N-(1-naphthyl)-ethylene diamine dihydrochloride in 2.5% phosphoric acid solution)와 동량으로 주입한 후 10분간 실온에서 방치하였다. 아질산의 농도는 ELISA 리더기를 이용하여 570 nm에서 흡광도를 측정하여 결정 하였다. 세포가 없는 배양액은 아질산이 0~30μM로 나타 나, 이 값을 표준으로 하여 여러 실험군의 아질산 값의 흡광도를 측정하였다.
4) MTS 분석
RAW 264.7 세포(5×104 cells/well)를 96-well culture plate에 100μl의 RPMI 1640 배지와 함께 하룻밤 배양한 다음, MGH 추출물(50, 100, 250, 500μg/ml)을 각각 처 리하여 24시간 배양하였다. 각 well에 5 mg/ml 농도의 MTS 용액을 50μl씩 넣은 후 2시간 동안 배양하면서 환 원반응을 유도하여 발색정도를 microplate reader를 이용 하여 550 nm에서 흡광도를 측정하였으며, 세포독성(cell toxicity)은 세포만 배양한 비처리군의 생존도 100%를 기 준으로 약물처리군의 상대적인 세포 생존도를 계산하였 다.
5) ELISA
RAW 264.7 세포를 RPMI 1640 배지를 이용하여 1.5×105 cells/ml로 조절한 후 24 well plate에 접종하고, 5% CO2
항온기에서 18시간 전 배양하였다. 이후 배지를 제거하고 MGH를 각각 100, 250μg/ml 농도로 1시간 동안 처리한 후, 그람-음성 박테리아 내독소인 LPS (200 ng/ml)로 RAW 264.7 대식세포주를 자극한 후 세포 부유액을 원심 분리하여 세포들을 침전시켜 상층액을 수집하고, 상층액
내 IL-6 생성량을 ELISA kit (R&D Systems Inc., Minnea- polis, MN, USA)를 이용하여 사용자 매뉴얼에 기재된 방 법대로 정량해 분석하였다.
6) PGE2 측정
RAW 264.7 세포를 RPMI 1640 배지를 이용하여 1.5×105 cells/ml로 조절한 후 24 well plate에 접종하고, 5% CO2
항온기에서 18시간 전 배양하였다. 이후 배지를 제거하고 MGH를 각각 100, 250μg/ml 농도로 1시간 동안 처리한 후, 그람-음성 박테리아 내독소인 LPS (200 ng/ml)로 RAW 264.7 대식세포주를 자극한 후 세포부유액을 원심 분리하여 세포들을 침전시켜 상층액을 수집하고, 상층액 내 PGE2 생성량을 EIA kit (R&D Systems Inc., Minnea- polis, MN, USA)를 이용하여 사용자 매뉴얼에 기재된 방 법대로 정량해 분석하였다.
7) RNA 분리 및 RT-PCR
MGH에 의한 COX-2와 iNOS 발현과의 상관성을 알아 보기 위하여 RT-PCR로 mRNA 발현을 조사하였다. RNA 및 RT-PCR RNA 세포의 준비는 6-well culture plate에 3×105 세포로 분주한 다음, 하루 밤 동안 안정화시켰다.
이 세포에 MGH 물 추출물(100, 250μg/ml)을 처리한 후, LPS로 자극하고 24시간 후에 세포를 모아 PBS로 세 척하여 이지 블루(Easy Blue, 인트론사) 1 ml를 가하여 실온에서 교반하였다. chloroform 200μl를 넣고 다시 교 반하여 13,000 rpm, 4oC에서 10분간 원심분리 한 다음, 상등액 400μl에 isopropanol을 가하여 다시 원심분리하 여 RNA pellet을 얻었다. 여기서 얻어진 RNA에 MuLV역 전사효소(reverse transcriptase), 1 mM dNTP 0.5μg을 넣어 cDNA를 만들었다. 여기에 COX-2, iNOS와 β-actin primer를 넣고 유전자 증폭기(thermal cycler)를 이용하여 증폭시켰다. 이때 iNOS는 94oC에서 2분, 55oC에서 30초, 72oC에서 30초 동안 반응시켰으며, COX-2와 β-actin은 94oC에서 2분, 58oC에서 30초, 72oC에서 30초 동안 반응 시켰다. 만들어진 RNA를 2% agarose gel에 전기 영동시 켜 UV 검출기로 확인하였다.
8) Western blot analysis
배양이 끝난 세포를 수집하여 2~3회 PBS (phosphate buffered saline)로 세척 한 후 1 ml의 lysis buffer을 첨가
Fig. 1. Effect of MGH on cell viability in RAW 264.7 cells.
The cell viability assessed using an MTS assay following in- cubation with different doses (50, 100, 250 and 500μg/ml) of MGH for 24 hours. Values are the mean±S.E. of duplicate de- terminations from three separate experiments.
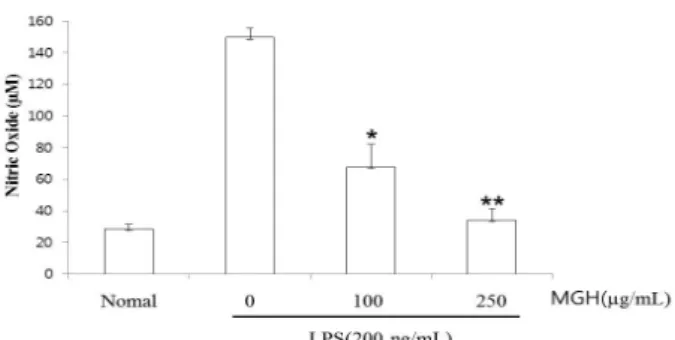
Fig. 2. Effect of MGH on LPS-induced NO production in RAW 264.7 cells. RAW 264.7 cells were pretreated with the in- dicated concentration of MGH for 30 minutes before being in- cubated with LPS (200 ng/ml) for 24 hours. The culture super- natant was subsequently isolated and analyzed for LPS treated group. Statistical significance: *p<0.05, **p<0.005, Significant differences between treated groups were determined using the Dunnett’s t-test. Values are the mean±S.E. of duplicate deter- minations from three separate experiments.
하여 30분간 lysis 시킨 후 12,000 rpm에서 20분간 원심 분리하여 세포막 성분 등을 제거하였다. 단백질 농도는 BSA (bovine serum albumin)를 표준화하여 Bio-Rad Protein Assay Kit를 사용하여 정량하였다. 20~30μg의 lysate를 8~12% mini gel SDS-PAGE로 변성 분리하여, 이 를 PVDF (polyvinylidene difluoride) membrane (BIO- RAD, Richmond, CA, USA)에 200 mA로 2시간 동안 transfer하였다. 그리고 membrane의 blocking은 5%
skim milk가 함유된 TBST (0.1% Tween20+TBS) 용액 에서 상온에서 2시간 동안 실시하였다. MAPKS의 발현 양을 검토하기 위한 항체로는 JNK, ERK anti-mouse (1:1,000) (Calbiochem, La Jolla, CA, USA)를 TBST 용액 에서 희석하여 상온에서 2시간 반응시킨 후 TBST로 3회 세정하였다. 2차 항체로는 HRP (horse radish perox- idase)가 결합된 anti-mouse IgG (Amersham Pharmacia Biotech, Little Chalfont, UK)를 1:2,000으로 희석하여 상 온에서 1시간 반응시킨 후, TBST로 3회 세정하여 ECL 기 질(Amersham Biosciences, Piscataway, NJ, USA)과 1~3 분 간 반응 후 X-ray 필름에 감광하였다.
9) 통계분석
모든 실험은 3회 이상 반복으로 이루어졌으며, 통계분 석은 SPSS 11.5를 사용하였다. 실험결과는 각 항목에 따 라 평균치±표준편차(SEM)를 구하여 신뢰수준 *p<0.05,
**p<0.005에서 통계적 유의성을 검증하였다.
결 과»»»
1. 세포생존율에 대한 MGH의 영향
RAW 264.7 세포에서 MGH의 세포독성을 MTS assay 방법으로 조사하였다. 그 결과 MGH를 50, 100, 250, 500 μg/ml의 농도로 처리하였을 때 50~250μg/ml의 농도에 서 세포의 생존율에 영향을 미치지 않았다(Fig. 1). 따라 서 이후 실험은 MGH의 물 추출물이 세포독성이 나타나 지 않는 농도범위에서 수행하였다.
2. Nitric oxide (NO) 생성에 대한 MGH의 억제 효과
NO는 활성산소 중 하나이며, 최근 염증 유발에 중요한 역할을 하는 것으로 알려져 있다. 따라서 NO 생성에 대 한 MGH의 효과를 알아보기 위해 Griess assay 방법을 이 용하여 세포 배양액 중에 존재하는 NO2−의 형태로 측정 하였다. MGH 물 추출물은 100μg/ml, 250μg/ml의 처 리농도에서 각각 농도 의존적이고 유의적으로(각각 p<
0.05, p<0.005) NO 생성이 억제되는 것으로 나타났다 (Fig. 2).
Fig. 3. Effect of MGH on LPS-induced iNOS mRNA expression in RAW 264.7 cells. iNOS mRNAs were assessed by RT-PCR in RAW 264.7 cells. Cells were pretreated with the indicated con- centrations of MGH for 30 minutes before being incubated with LPS (200 ng/ml) for 24 hours. The β-actin mRNA was assayed in parallel to confirm equivalency of the cDNA pre- paration. The experiment was repeated three times, and sim- ilar results were obtained.
Fig. 5. Effect of MGH on LPS-induced COX-2 mRNA ex- pression in RAW 264.7 cells. COX-2 mRNAs were assessed by RT-PCR in RAW 264.7 cells. Cells were pretreated with the in- dicated concentrations of MGH for 30 minutes before being in- cubated with LPS (200 ng/ml) for 24 hours. The β-actin mRNA was assayed in parallel to confirm equivalency of the cDNA preparation. The experiment was repeated three times, and similar results were obtained.
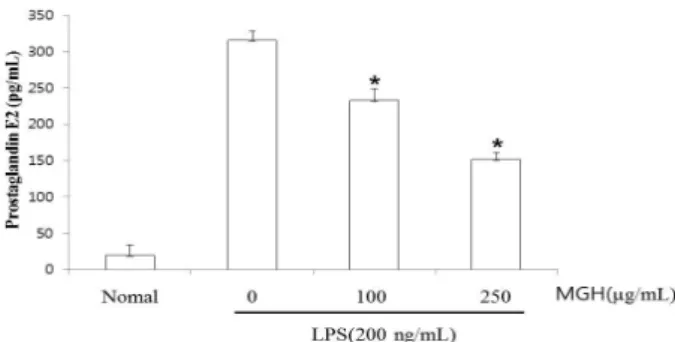
Fig. 4. Effect of MGH on LPS-induced PGE
2production in RAW 264.7 cells. RAW 264.7 cells were pretreated with the in- dicated concentration of MGH for 30 minutes before being in- cubated with LPS (200 ng/ml) for 24 hours. The culture super- natant was subsequently isolated and analyzed for LPS treated group. *P<0.05, when compared to the LPS treated group.
Significant differences between treated groups were determined using the Dunnett’s t-test. Values are the mean±S.E. of dupli- cate determinations from three separate experiments.
3. iNOS mRNA 발현에 대한 MGH의 억제 효과
NO는 iNOS에 의해 생성되어 염증 상태에서 혈관 투과 성, 부종 등의 염증반응을 촉진시킬 뿐만 아니라 염증매 개체의 생합성을 촉진하여 염증을 심화시키는 것으로 알 려져 있다. 따라서 NO를 생성하게 하는 iNOS의 mRNA 발현을 알아 보기위해 RT-PCR을 수행하였다. 그 결과, LPS 처리에 의해 형성되는 iNOS mRNA 발현 모두 100, 250μg/ml에서 현저한 억제 양상을 보였다(Fig. 3).
4. PGE
2
생성에 대한 MGH의 억제 효과대식세포 매개 PGE2는 COX-1과 COX-2로부터 생성되 는데, 소량의 경우 생체대사에 필수적이지만 유도성 COX-2 의 과발현에 의한 과량의 PGE2는 강력한 염증 매개물질 로 작용하는 것으로 보고되어 있다. 따라서 본 연구에서 는 PGE2를 대상으로 MGH의 약리학적인 효능 평가를 실 시하였다. MGH를 각각 100μg/ml, 250μg/ml의 농도로 30분 동안 처리한 후, 그람-음성 박테리아 내독소인 LPS (200 ng/ml)로 RAW 264.7 대식세포주를 자극한 후 세포 부유액을 원심분리하여 세포들을 침전시켜 상층액을 수 집하고, 상층액 내 PGE2 생성량을 측정하였다. 그 결과, control에서는 PGE2 생성량이 매우 낮게 측정되었으며, LPS에 의해 현저히 증가되었다. MGH는 100μg/ml, 250 μg/ml의 처리농도에서 각각 농도 의존적으로 PGE2 생 성이 유의성 있게(p<0.05) 억제되는 것을 확인할 수 있 었다(Fig. 4).
5. COX-2 mRNA 발현에 대한 MGH의 억제 효과
MGH의 COX-2 mRNA 발현에 대한 효과를 알아보기 위해 Western blot과 RT-PCR을 수행하였다. 그 결과, LPS 처리에 의해 증가된 COX-2 mRNA 발현은 100μg/ml, 250μg/ml 농도에서 현저하게 억제되는 것을 확인하였다 (Fig. 5).
6. IL-6 생성에 대한 MGH의 억제 효과
IL-6는 염증단계에서 중추적 역할을 한다고 알려져 있 다. 대식세포주인 RAW 264.7 세포로부터 IL-6의 생성을
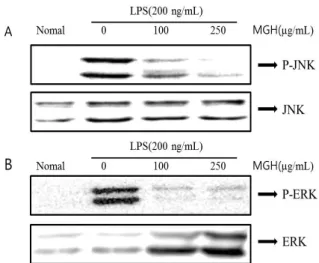
Fig. 7. Effect of MGH on the phosphorylation (P-) of MAPKs in LPS-stimulated RAW 264.7 cells. RAW 264.7 cells were treated with the indicated concentrations of MGH for 30 mi- nutes before being incubated with LPS (200 ng/ml) for 30 minutes. Whole-cell lysates were analyzed by western blot analysis. The experiment was repeated three times, and similar results were obtained.
Fig. 6. Effect of MGH on LPS-induced IL-6 production in RAW 264.7 cells. RAW 264.7 cells were pretreated with the in- dicated concentrations of MGH for 30 minutes before being in- cubated with LPS (200 ng/ml) for 24 hours. Production of IL-6 was measured by ELISA. Cells were pretreated with the in- dicated concentrations of MGH for 30 minutes before being in- cubated with LPS (200 ng/ml) for 6 hours. *p<0.05, when compared to the LPS treated group. Significant differences be- tween treated groups were determined using the Dunnett’s t-test. Values are the mean±S.E. of duplicate determinations from three separate experiments.
ELISA를 이용하여 실험하였다. 그 결과, LPS 자극원을 처 리하였을 때 IL-6 생성이 현저히 증가하였고, MGH를 처 리하였을 때 IL-6 생성 억제 효과가 유의성 있게(p<0.05) 나타났다(Fig. 6).
7. MAPKs의 인산화에 대한 MGH의 억제 효과
MAPKs는 세포의 성장과 분화 및 cytokine과 stress 제 어에 중요한 역할을 한다. 따라서 MGH의 억제 메커니즘 이 MAPKs를 경유하는지 알아보기 위해 MAPKs의 인산화 를 Western blot을 통해 확인하였다. 그 결과 LPS에 의해 활성화된 RAW 264.7 대식세포에 MGH를 100, 250μg/ml 을 처리한 경우 JNK1/2와 ERK1/2 인산화를 억제하는 것 을 확인하였다(Fig. 7). 그러나 p38의 인산화에는 영향이 없는 것을 확인할 수 있었다.
고찰»»»
戊己丸은 宋代 「太平惠民和劑局方」8)에 처음 기록된 처 방으로 黃連, 吳茱萸, 白芍藥으로 구성되어 脾受濕氣 泄痢 不止 米穀遲化 臍腹刺痛을 치료한다고 하였다. 또한 許9) 는 「東醫寶鑑」에 痢疾의 한 종류인 濕痢를 치료한다 하
였다.
戊己丸의 구성 약재의 본초학적 효능으로 黃連은 大苦 大寒하면서 質이 燥하여 淸熱燥濕의 효능이 매우 강력할 뿐 아니라 解毒 작용을 겸하고 있기 때문에 胃腸의 濕熱 로 인한 嘔吐, 瀉痢, 痔瘡 등의 병증에 응용하며, 吳茱萸 는 性味가 辛苦熱하면서 燥하여 溫散開鬱의 효능이 있으 며, 熱證에 속한 병증에도 황련과 같은 寒冷藥物을 배합 하여 응용한다. 또한 白芍藥은 濕熱下痢증으로 인한 腹 痛, 裏急後重의 병증에 黃芩, 黃連, 大黃, 木香, 甘草 등의 調和氣血, 淸熱解毒, 止痢의 효능이 있는 약물을 배합하 여 응용할 수 있다18).
이러한 약재들의 효능과 배합으로 볼 때 戊己丸은 이 질과 같은 증상뿐만 아니라 염증성 질환을 치료하는데 사 용할 수 있을 것으로 생각되어, 본 연구에서는 설치류 대 식세포주인 RAW 264.7 세포를 이용하여 LPS로 유도된 염증반응에서 戊己丸이 어떤 작용 기전을 통해 항염증 효 과를 나타내는지 연구하였다.
염증은 자극에 대한 생체조직의 방어반응의 하나로서 감염에 의한 외인적인 요인 및 조직의 stress와 기능부전 에 의한 내인적인 요인에 의해 유도되며19) 손상된 조직을 수복하거나 재생하려는 기전이다20). 그러나 지속적인 염 증반응은 점막손상을 촉진시켜 결과적으로 통증, 부종,
발적, 발열 등을 일으켜 기능장애를 유발하며 당뇨병, 동 맥경화증, 관절염 및 암 등의 발생과 깊은 연관을 갖고 있다21,22).
선천 면역을 담당하는 대식세포는 능동 및 수동 면역 반응에서 매우 중요한 역할을 한다. 또한, 초기 염증반응 에 관여하는 대표적인 면역세포로서 여러 자극이나 면역 세포들이 분비하는 cytokine 등에 의해 활성화되어 TNF- α, IL-1, IL-6와 같은 pro-inflammatory cytokine을 비롯 하여 eicosanoids, ROS, NO, PGE2, superoxide (O2−
) 등 의 다양한 염증 매개물질들을 생성함으로써 통증, 부종, 열 등의 염증 반응을 유발하고, 염증부위로 면역세포의 이동을 촉진시킨다23-26). 최근, LPS에 의한 대식세포의 활 성은 toll-like receptor (TLR) 4의 발현을 조절한다고 알 려져 있다. LPS와 TLR4가 결합하여 활성화되면 세포질 조절 단백질인 MyD88이 모이게 되고 여러 신호전달기전 을 통하여 전사인자인 nuclear factor-κB (NF-κB)가 세 포질내로 이동하여 활성화된다고 보고되고 있다27). 이러 한 NF-κB 전사인자의 활성화는 염증매개물질들인 NO, prostaglandins (PGs) 그리고 pro-inflammatory cytokine 등을 조절한다28).
NO는 반응성이 높은 물질로 NO synthase (NOS)에 의 해 L-arginine으로부터 생성되며, 신경전달과 혈관확장 등 생리적인 기능을 조절하는 중요한 역할을 하지만, 과발현 된 NO는 혈관 투과성, 부종, 염증을 심화시켜 조직을 손 상시키고 암으로의 진행을 촉진한다29,30). NOS는 consti- tutive NOS와 inducible NOS (iNOS)로 나누어지는데, 특 히 iNOS는 외부자극이나 pro-inflammatory cytokine 등 에 의해 자극 받게 되면 hepatocytes, smooth muscle cells, bone marrow cells, monocytes, macrophages 등 다양한 세포에서 발현되어 다량의 NO를 생성한다고 보고 되고 있다31).
염증반응의 중요한 매개체인 prostaglandin (PG) E2는 COX에 의해 생성되는데, COX에는 COX-1과 COX-2 두 종류가 있으며, COX-1은 생체 내의 대부분의 조직에 존 재하며 일정하게 발현되는 반면, COX-2는 염증자극원에 의해 macrophage와 같은 염증성 세포에서 유도되며, PGE2와 같은 다량의 PGs를 생성함으로써 염증관련 질병 을 유발하는 것으로 밝혀져 있다32). 또한 IL-6, IL-1β, TNF-α는 염증반응을 촉진하며 면역계를 자극하는 염증 유발인자로 종양의 침습에도 관련이 있다고 알려져 있
다33).
이러한 사실에 기초하여 MGH가 RAW 264.7 세포에서 LPS에 의해 유도된 NO의 생성을 저해함을 확인하였다.
또한 Western blot으로 분석한 결과 MGH에 의한 iNOS 의 발현 억제는 NO 형성 억제와 유사한 경향을 나타냄으 로써 NO 형성 억제는 iNOS의 발현 저해를 경유한 것임 을 알 수 있었다. MGH가 LPS에 의해 유도된 iNOS 단백 질 및 mRNA 발현을 저해되는 것으로 보아 MGH가 LPS 에 의해 유도된 iNOS의 전사와 번역 모두를 저해한다고 추정된다. 다수의 항염증 약물들의 작용기전은 prosta- glandin 합성을 억제하며 이는 COX-2의 생성 및 효소 활 성저해에 의한 것이다. 따라서 COX-2에 의한 prosta- glandin의 합성은 염증반응을 매개하는 것으로 여겨지고 있다고 보고 MGH가 LPS에 의해 형성되는 PGE2를 유의 성있게 감소시키며 이는 COX-2 단백질과 mRNA의 발현 저해에 의한 것임을 확인할 수 있었다. 염증매개물질인 IL-6는 in vivo 및 in vitro 모두에서 염증반응을 조절하는 것으로 알려져 있다. 이러한 cytokine들은 서로 상호작용 이 있는 것으로 알려져 있으며 LPS등의 염증 자극 물질에 의해 생성이 유도된다고 보고되었다34). 따라서 본 연구에 서 LPS에 의해 유도된 IL-6의 생성에 대한 MGH의 억제 효과를 실험하였다. 그 결과, IL-6의 생성을 ELISA로 분석 하고, 이러한 COX-2, iNOS 그리고 pro-inflammatory cy- tokines의 발현에 대해 조사하였다.
또한, 현재까지 비교적 잘 알려진 염증반응의 분자신호 전달기전은 MAPK superfamily에 속하는 세 가지 효소들 로 extracellular signal-regulated kinase (ERK), c-Jun NH2-protein kinase (JNK)/stress-activated protein kin- ase (SAPK), serine/threonine protein kinase인 p38 MAPK 등을 들 수 있다. 이들 MAPKs는 모두 다양한 세 포외 자극에 반응해 upstream MAPK kinase (MEK)에 의 해 tyrosine과 threonine에서 인산화가 일어남으로써 활 성화된다. 위의 MAPKs의 활성형은 그 후에 다른 kinase 나 전사 인자를 인산화, 활성화시키고, 결국 표적 유전자 의 발현을 변화시킨다. 어떤 보고에 의하면 RAW 264.7 세포에서 LPS로 유도된 COX-2 발현이 MAPK 저해제들인 PD98059와 SB203580에 의해 ERK1/2와 p38 MAPK 활성 화가 각각 차단됨으로써 부분적으로 억제된다는 것을 증 명했다35). 유사하게 마우스 대식세포 J774에서 phospho- lipase A2 저해제인 methyl arachidonyl fluorophospho-
nate에 의한 COX-2 유도는 PD98059와 SB203580을 포함 한 MAPK 저해제에 의해 억제되었다36). 마찬가지로 LPS 는 단핵세포에서 ERK2와 p38 MAPK의 인산화와 활성화 처럼 COX-2 단백질과 그것의 mRNA 전사물이 발현을 유 도하였다는 보고가 있다37).
따라서 본 연구에서도 MGH가 NO와 COX-2, IL-6와 같 은 염증매개체들을 어떠한 신호전달을 통해 억제하는지 알아보기 위해 MAPKs의 인산화를 억제하는지 알아보았 다. 그 결과 MGH가 MAPKs의 인산화에서는 JNK1/2와 ERK1/2의 인산화는 억제하는 반면, p38 MAPK의 인산화 에는 아무런 영향을 주지 못했다. 이는 LPS로 유도한 대 식세포의 염증반응은 JNK1/2와 ERK1/2의 신호전달 경로 를 경유하는 반면 p38의 경로와는 다르다는 것을 보여준 다.
본 실험 결과를 요약하면, MGH는 RAW 264.7 세포에 서 LPS에 의해 유도되는 iNOS, COX-2, 그리고 IL-6 유전 자의 발현, PGE2 생성을 효과적으로 저해한다. 그러므로 이러한 결과들은 한의학에서 MGH의 항염증 기전을 밝힘 으로써 그 과학적 근거를 제시할 수 있을 것으로 생각된 다.
결론»»»
설치류의 대식세포주인 RAW 264.7 세포를 LPS로 자 극하였을 때 戊己丸의 항염증 효과를 실험하여 다음과 같 은 결론을 얻었다.
1. 戊己丸은 LPS로 유도된 대식세포에서 NO 생성을 농도 의존적으로 유의성 있게(100μg/ml, 250μg/ml의 처리농도에서 각각 p<0.05, p<0.005) 억제하였다.
2. 戊己丸은 LPS로 유도된 대식세포에서 PGE2 생성을 농도 의존적으로 유의성 있게(p<0.05) 억제하였다.
3. 戊己丸은 LPS로 유도된 iNOS와 COX-2의 mRNA발 현을 억제하였다.
4. 戊己丸은 LPS로 유도된 전염증성 사이토카인 IL-6발 현을 유의성 있게(p<0.05) 억제하였다.
5. 戊己丸은 LPS로 유도된 MAPKs JNK 1/2, ERK1/2의 인산화를 억제하였으나 p38 인산화는 억제하지 못하였 다.
이와 같은 결과로 보아 戊己丸은 대식세포에 작용하여
MAPKs의 인산화 활성의 저해를 통해 NO, PGE2, IL-6의 생성과 iNOS, COX-2 발현을 억제함으로써 항염증 효과 가 있음을 알 수 있다.