서 론
잉어목 미꾸리과에 속하는 왕종개 Iksookimia longicorpa는 한국고유종으로서, 물이 맑고 유속이 빠른 돌과 자갈 바닥에 서 서식하면서 주로 수서곤충 및 부착조류를 먹는 것으로 알 려져 있다(Kim, 1997; Kim and Park, 2002).
어류의 안구(eyeball)는 외부조직인 눈꺼풀이 없고 구형 의 수정체(lens)를 가지고 있으며, 이러한 수정체는 포유류와 다르게 탄력성이 없기 때문에 수정체 후인근(retractor lentis muscle)을 통해 전후이동으로 초점거리를 맞추는 것으로 보고 되어 있다(Nicol, 1989; Kim, 1997). 또한 안구와 연결되는 시 신경은 고등한 척추동물에서는 교차부에서 서로 접합되어 있 는 반면 어류의 시신경은 좌·우안이 각각 교차되어 뇌에 연 결되어 있고 교차부에서 서로 접합되진 않는 것으로 알려져 있다(George and Robert, 2001).
척추동물의 안구는 각막(cornea)과 공막(sclera)으로 구성되 는 외막(outer coat)과 홍체(iris), 모양체(ciliary body), 맥락막 (choroid)으로 구성된 중막(middle coat) 그리고 망막(retina)이 존재하는 내막(inner coat)으로 구분되며(George and Robert, 2001; You and Park, 2008), 망막은 안구에서 내벽에 위치하 며 외부로부터 들어오는 빛을 감지하는 신경조직의 연합층으 로 알려져 있다(Polyak, 1957; Hagedorn et al., 1998). 망막의 한 층인 시각세포층에는 원추세포(cone cell)와 간상세포(rod cell)가 존재하며 원추세포는 비교적 강한 파장의 빛을 감지하 여 상을 구분하게 되고 간상세포는 약한 파장의 빛을 감지해 명암을 구별하는 역할을 한다(Peter and Joseph, 1996). 시각세 포의 종류는 각각 동물의 서식방법과 밀접한 연관을 가지는 것으로 보고되어 있는데 주로 낮에 활동하는 척추동물은 원추 세포와 간상세포 모두를 가지고 있지만 원추세포의 밀도가 더 높은 반면에 야행성 척추동물의 망막에는 거의 간상세포로 구 성되어 있다(Audesirk et al., 2008).
경골어류의 안구에 관한 연구는 광학현미경을 이용한 시 각세포의 형태학적 분석(Lyall, 1957; Collins and MacNichol,
—257 — http://www.fishkorea.or.kr
* Corresponding author: Jong Young Park Tel: 82-63-270-3344, Fax: 82-63-270-3362, E-mail: [email protected]
ISSn: 1225-8598(Print), 2288-3371(online)
accepted: December 21, 2015
한국산 미꾸리과 어류 왕종개 Iksookimia longicorpa 망막의 시각세포
김재구
·
박종영*
전북대학교 자연과학대학 생물학과, 전북대학교 생물다양성연구소
Visual Cells in the Retina of Iksookimia longicorpa(Pisces; Cobitidae) of Korea by Jae Goo Kim and Jong Young Park*(faculty of Biological Science and Institute for Biodiversity research, college of natural Sciences, chonbuk national university, Jeonju 54896, Korea)
ABSTRACT The visual cells in the retina of Iksookimia longicorpa(Pisces, Cobitidae) were investigated by light and scanning electron microscopes. The retina(216.42±13.36µm) has several layers, and the visual cell layer consists of unequal double cones and large rods. In a double cone, two members are unequal such that one cone is longer than the other (long element 26.42±1.7 μm, short element 16.82±1.1µm). The cones form a row mosaic pattern in which the partners of double cones are linearly oriented with a large rod. The visual cells observed have an outer segment (hematophilic), inner segment(eosinophilic). In scanning electron microscopy, the outer segment links to inner segment by so-called calyceal piles(calyceal processes) of membrane discs surrounded by double membranes.
Keywords : Iksookimia longicorpa, retina, visual cells, unequal double cone, rod cell, row mosaic model
1978; Monica, 2001; Thomas and Craig, 2010)과 원추세포의 모자이크 모델에 관한 연구(Lyall, 1956; Rossetto et al., 1992;
Yuko et al., 1997), 명암조건에 따른 원추세포의 변화(Yvette, 1980; Fernald, 1982; Frank et al., 2001) 등에 대하여 비교적 활발한 연구가 진행되어 왔으나 국내의 연구에는 생태적 차이 에 의한 두 종 간의 망막 조직의 비교연구(Lee and Lim, 2005;
You and Park, 2008) 이외에는 매우 미흡한 실정이며 특히 간 상세포와 원추세포의 정확한 형태에 관한 연구는 전무한 실정 이다.
따라서 본 연구는 한국고유종인 왕종개의 망막구조 및 시각 세포들의 형태를 확인하여 한국고유종의 미세서식처를 포함 한 수중환경과 망막 및 시각세포와의 상관관계를 규명하고자 하며 담수어류의 망막구조에 대한 기초자료를 제시하고자 하 였다.
재료 및 방법
1. 실험재료
왕종개(n=10, 131.40±4.9mm SL)는 2014년 11월에 전라 북도 순창군 복흥면 반월리(35°26ʹ52 N; 126°55ʹ1 E) 일대에서 족대(망목 4×4mm)를 이용하여 채집하였다.
2. 실험방법
왕종개의 안구와 외부형태를 조사한 후 망막의 시각세포를 관찰하기 위해 광학현미경과 주사전자현미경을 이용하여 망 막구조 및 시각세포의 형태를 확인하였다.
1) 광학현미경 관찰
채집된 개체의 안구를 적출하여 4°C의 10% formalin에 12 시간 고정하고 12시간 수세하였다. 70~100%의 ethanol을 이
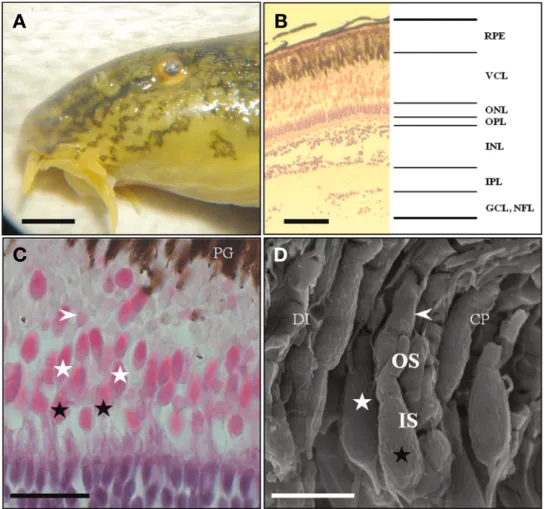
Fig. 1. External morphology and gross sections of the retina in a Harris hematoxylin and counter-stained with eosin used by light microscope and scanning electron microscope of Iksookimia longicorpa. (A) Left eye side view(bar=4mm). (B, C) Longitudinal sections of the retina(bars=
50, 20μm). (D) Scanning electron micrograph(bar=10μm). CP, calyceal process; DI, disc; GCL, ganglion cell layer; INL, inner nuclear layer;
IS, inner segment; IPL, inner plexiform layer; NFL, nerve fiber layer; ONL, outer nuclear layer; OPL, outer plexiform layer; OS, outer segment;
PG, pigment granules; RPE, retina pigment epithelium; VCL, visual cell layer. Black asterisks, short cones; white asterisks, big cones; arrow- heads, rod cells.
A B
C D
용하여 탈수하였고 xylene으로 치환하여 paraffin으로 포매하 였다. 완성된 block은 microtome(Leica-820, Germany)으로 5 μm 간격으로 절단하여 hematoxylin과 eosin으로 이중 염색하 였으며, 광학현미경(Carl zeiss-AX10, Germany)으로 관찰하였 다. 시각세포층을 확인하고자 수직으로 절단하였으며, 모자이 크 패턴을 확인하기 위해 수평으로 절단하였다. 고배율의 해 상도를 위하여 일부 수직단면은 epon block을 제작하여 1μm 이하로 semi-thin section한 시료를 toluidine으로 염색한 후 광 학현미경으로 관찰하였다.
2) 주사전자현미경 관찰
채집된 개체의 안구를 적출하여 0.1M phosphate buffer(pH 7.4)로 조정된 2.5% glutaraldehyde에 4°C에서 4시간 전고 정 후 1% osmium tetroxide(OSO4)로 90분간 후고정하였으 며 ethanol 상승 순으로 탈수시켰다. Tert-butyl alcohol(TBA) 을 이용하여 동결 건조시킨 뒤 해부현미경(Carl Zeiss, Stemi- DV4, Germany)을 이용하여 망막 부분을 상위로 stub에 부 착시켰다. 이후 stub를 OSO4로 증착시켜 주사전자현미경
(Hitachi, SN-3000, Japan)을 이용하여 20kV에서 관찰 및 촬 영하였다.
결 과
1. 안구의 외부형태
왕종개의 안구는 3.53±0.2mm의 크기였으며 두장대 안경 비율은 14.08±0.7이였다. 가로안경과 세로안경이 거의 유사 한 구형의 안구를 가지고 있었다(Fig. 1A).
2. 광학현미경적 관찰
1) 수직단면(longitudinal plane)의 관찰
왕종개의 망막은 가장 안쪽부터 색소포가 많은 망막 색소 상피층(retina pigment epithelium)이 존재하였고 망막 색소 상피층과 만나는 부분부터 시각세포들이 존재하는 시각세 포층(visual cell layer), 그리고 이 시각세포의 핵들이 존재하
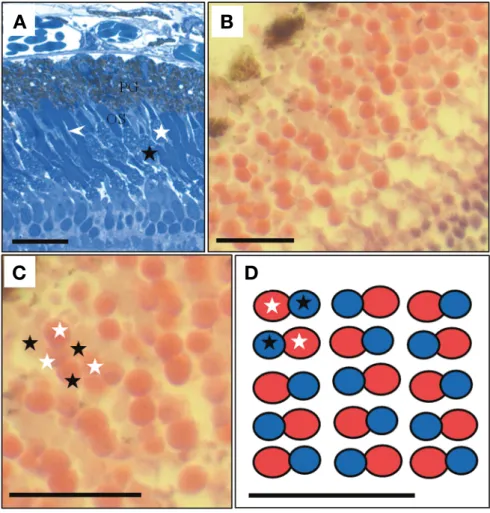
Fig. 2. Visual cell layer by semi-thin sections in the Iksookimia longicorpa retina. (A) Longitudinal sections of the visual cell layer. (B, C) Trans- verse sections showing a mosaic model in the visual cells by light microscopy. (D) Diagram based on the left micrograph. OS, outer segment;
PG, pigment granules. Black asterisks, short cones; white asterisks, big cones; arrowheads, rod cells(bars=20μm).
A B
C D
는 바깥핵층(outer nuclear layer), 바깥얼기층(outer plexiform layer), 속핵층(inner nuclear layer) 등으로 명확히 구분되는 신 경연합층의 구조를 가지고 있었다(Fig. 1B). 전체 망막의 두께 는 216.42(±13.36)μm였으며, 시각세포의 핵이 존재하는 바 깥핵층의 두께는 13.42(±2.16)μm였다(Figs. 1B, 2A). 시각 세포층은 긴 원추세포와 짧은 원추세포로 구성된 이형 이중 원추세포와 간상세포로 구성되어 있었다. 시각세포들은 내절 (inner segment)과 외절(outer segment)로 구분되었으며 내절 은 호염성 염색액인 eosin에 강하게 염색되었고 외절은 호산 성 염색액인 hematoxylin에 염색되어 나타났다(Fig. 1C). 이형 이중 원추세포 중 긴 원추세포는 26.42±1.7μm의 길이였으며 직경은 5.12±0.6μm로 확인되었고, 짧은 원추세포는 16.82±
1.1μm의 길이였으며 직경은 긴 원추세포와 유사한 5.10±
0.31μm로 확인되었다. 간상세포는 길이 40.13±2.4μm였으며 직경 3.95±0.4μm로 확인되었다(Fig. 1C, D). 따라서 가장 긴 세포는 간상세포였으며 직경은 원추세포들이 더 두껍게 확인 되었다.
2) 수평단면(transverse plane)의 관찰
왕종개의 시각세포층을 수평 절단하여 관찰한 결과, 이형 이중 원추세포가 규칙적인 배열로 인해 열 배열(row-type)의 모자이크 형태로 나타났다(Fig. 2B, C, D). 긴 원추세포와 짧 은 원추세포가 하나의 쌍을 이루었으며 긴 원추세포는 내절이 절단되어 eosin에 염색되었으며 짧은 원추세포는 외절이 절단 되어 hematoxylin에 염색되었다. 간상세포는 원추세포보다 상 위에 존재하기 때문에 수평단면 관찰에서는 확인할 수 없었다 (Fig. 2C).
3. 전자현미경적 관찰
왕종개 시각세포를 자세히 확인하기 위해 주사전자현미경 을 이용하여 관찰한 결과, 긴 원추세포와 짧은 원추세포가 명 확히 구분되었으며 간상세포 또한 확인할 수 있었다(Fig. 1D).
원추세포와 간상세포는 광학현미경 관찰 결과와 같이 내절과 외절로 구분되었고 원추세포의 내절은 장타원형으로 확인되 었다(Fig. 1C, D). 외절에는 디스크구조의 판상을 확인할 수 있 었으며 내절과 외절을 연결해주는 배상극(calyceal processes) 들이 내절과 외절 가장자리로 분포하고 있었다. 간상세포의 외절은 원추세포의 외절보다 길었으며, 내절은 원추세포의 내 절이 더 큰 것으로 확인되었다. 또한 원추세포와 간상세포 모 두 내절 표면은 무수히 많은 과립형태가 관찰되었다.
고 찰
한국고유종인 왕종개의 시각세포를 광학현미경과 전자현미
경을 이용하여 조사한 결과, 왕종개의 안구는 33.53±0.2mm 의 크기였으며 두장대 안경비율은 14.08±0.7이였다. 가로안 경과 세로안경이 거의 유사한 구형의 안구를 가지고 있었다. Kim(1997)의 문헌에서도 왕종개는 전체 체장 및 두장에 대한 안경비율은 매우 낮은 종으로 안구의 크기가 작은 편에 속하 였다.
왕종개의 망막은 다른 어류와 같이 기본적인 망막구조인 신경연합층을 가지고 있었다. 시각세포들이 분포하는 시각 세포층과 이들의 핵이 존재하는 바깥핵층의 두께는 다소 차 이를 보였다(Hagedorn et al., 1998; Lee and Lim, 2005; You and Park, 2008). 한국산 담수어류 중 고유종인 꺽지(Kim et al., 2014)의 연구결과와 비교한 결과, 꺽지의 망막은 약 277.8 μm의 두께였으나 왕종개는 216.4μm로 얇았다. 비교적 긴 간 상세포의 외절을 가지고 있는 꺽지는 전체적인 망막의 두께 는 왕종개보다 두꺼웠다. 이러한 결과는 망막 내의 시각세포 층에 존재하는 시각세포의 차이에 의한 것으로 사료된다. 꺽 지는 동형 이중 원추세포를 가지고 있으며 짧은 단일원추세 포, 간상세포로 구성된다(Kim et al., 2014). 왕종개는 이형 이 중 원추세포와 간상세포로만 구성되기 때문에 바깥핵층의 두 께와 전체적인 망막 및 시각세포층의 두께가 얇은 것으로 사 료된다. 왕종개의 시각세포는 이형 이중 원추세포 중 긴 원 추세포(26.42±1.7μm), 짧은 원추세포(16.82±1.1μm) 및 간 상세포(40.13±2.4μm)로 구성되었다. Lyall(1957)은 경골어 류의 한 종인 Phoxinus laevis의 시각세포층에서 단일원추세 포, 이중원추세포, 삼중원추세포(triple cone cell), 사중원추세 포(quadruple cone cell)까지 총 4가지 형태의 원추세포를 확 인하였으며 Collins and MacNichol(1978)은 3종의 어류에서 삼중원추세포(triple cone cell)를 광학현미경상에서 확인하였 다. 일반적으로 어류는 물 속에서 서식하기 때문에 공기 중에 서와 다르게 빛은 수표면의 굴절로 인해 분산된다(Fernald, 1988; Meer van der, 1992). 따라서 분산된 다양한 파장대를 흡 수하는 여러 종류의 원추세포가 존재하게 된다(Audesirk et al., 2008). 단일 원추세포는 주로 단파장(short wavelength)에 서 최대 흡수력을 보이며 이중 원추세포는 중파장과 장파장 (middle and long wavelength)에서 최대 흡수력을 갖는다고 알 려져 있다(Marc and Sperling, 1976; Meer van der, 1992). 왕종 개는 비교적 낮은 수심의 담수환경에서 서식하기 때문에 긴 파장을 흡수하는 긴 원추세포와 짧은 원추세포로 구성되며, 하천의 바닥에서 모래 및 자갈 사이에 서식하기 때문에 낮은 빛환경에서 간상세포를 이용하여 명암 구별이 가능할 것으로 사료된다.
간상세포는 낮은 빛환경에서 명암을 구별하여 사물을 감 지할 수 있다. 심해어류에서 다층의 간상세포로만 구성된 망 막을 보유하는 종과 간상세포가 상당히 발달한 야행성어류 에 대하여 확인한 바 있다(George and Robert, 2001; Hirt and
Wagner, 2005). 기존의 국내 담수어류 연구(You and Park, 2008; Kim et al., 2014)에서는 간상세포를 확인하지 못하였으 나 본 연구에서는 왕종개의 시각세포층 내 긴 형태의 간상세 포를 관찰할 수 있었다. 이와 같이 담수 경골어류에서 간상세 포는 매우 길고 얇은 세포이기 때문에 구별 및 확인이 어렵다. 따라서 전자현미경을 이용하여 간상세포를 확인할 수 있었다.
왕종개의 시각세포층을 수평 절단하여 관찰한 결과, 이형 이중 원추세포가 규칙적인 배열로 인해 열 배열의 모자이크 형태로 확인되었다. 꺽지와 구피 Poecilia reticulata의 패턴 은 동형 이중 원추세포와 짧은 단일원추세포를 보유하기 때 문에 사각형의 패턴으로 구성되어 열 배열을 갖는 왕종개와 는 차이가 있었다(Yvette, 1980; Kim et al., 2014). 또한 계류에 서 서식한다고 알려진 인도의 미꾸리과 어류 3종의 연구에서 Nemacheilus beavani, N. devdevi는 정방형 배열로 나타내는 반면에, 비교적 유속이 빠른 곳에 서식하는 Balitora brucei에 서는 규칙적인 패턴을 찾을 수 없었다고 보고하였다(Nag and Bhattacharjee, 2002). 이들의 시각세포 구성은 왕종개와 같이 모두 이형 이중 원추세포와 간상세포로 구성되어 있었지만 시 각세포의 배열은 정방형의 사각 배열과 무작위적 배열로 나 타난다고 보고하여 왕종개와 배열의 차이를 보였다. 같은 과 (Family) 내에서의 시각세포 배열 차이는 그들의 서식환경 및 먹이를 먹는 습성에 따라 다른 것으로 사료된다. 비교적 차가 운 수온에서 서식하는 인도의 미꾸리과 Nemacheilus속은 유 속이 빠른 상류에서 돌 틈에 서식하고 주로 수서곤충을 섭식 하는 것으로 알려져 있다(Petr and Swar, 2002). 인도산 미꾸리 과 어류는 유속이 빠른 곳에서 수서곤충을 섭식하기 위해 한 국산 왕종개에 비해 먹이활동에서 시각의존성이 높을 것으로 사료된다.
주사전자현미경적 관찰에서 나타난 왕종개의 시각세포는 이형 이중 원추세포와 간상세포로 구분되었으며 내절에 관찰 된 많은 과립들로 인하여 미토콘드리아의 존재를 추정할 수 있었으며 디스크형태의 외절은 간상세포에서 길게 관찰되었 다. 또한 내절과 외절을 연결해주는 배상극은 원추세포와 간 상세포 모두 보유하고 있었다. 대부분의 어류에서 내절에는 많은 수의 미토콘드리아를 가지고 있다고 알려져 있으며 시각 세포의 형태에 따라 미토콘드리아의 밀도는 달라진다고 보고 되었다(Donatti and Fanta, 2007). 따라서 왕종개 내절에 분포 하는 과립형태는 미토콘드리아로 확인되었다. 본 연구에서 확 인된 배상극은 내절과 외절의 신호를 빠르게 전달하는 구조로 알려져 있으며 배상극의 수는 시각세포의 종류에 따라 다양한 것으로 보고된 바 있다(Frank et al., 2001; Sahly et al., 2012).
본 연구에서도 시각세포의 내절과 외절을 연결해 주는 배상극 이 관찰되었으나 이들의 정확한 수와 기능에 대하여 추가적인 연구가 필요할 것으로 사료된다.
왕종개의 시각세포층에는 이형 이중 원추세포와 간상세포
가 열 배열로 나타났으며 시각세포는 내절과 외절로 구분되 었다. 내절과 외절은 배상극으로 연결되어 있었으며 내절에는 미토콘드리아가 존재하고 외절에는 디스크 형의 구조가 관찰 되었다. 이러한 특징으로 볼 때 왕종개의 생태적인 특징 즉 낮 은 수심, 자갈 및 모래바닥에 서식하여 비교적 낮은 빛 환경에 서식하는 왕종개의 서식환경과 잘 일치하는 것으로 판단되며 추후 시각세포의 종류와 구조 등에 따른 서식환경과의 상호관 계에 관하여 지속적인 연구가 필요하다.
요 약
한국고유종인 왕종개의 안구를 관찰하고 망막에 존재하는 시각세포의 구조를 광학현미경과 주사전자현미경을 이용하 여 조사하였다. 왕종개의 안구는 3.53±0.2mm의 크기였으며 두장대 안경비율은 14.08±0.7이였다. 시각세포의 수직단면을 확인한 결과 망막은 여러 층으로 구성되어 있었으며 전체 망 막의 두께는 216.42(±13.36)μm였으며, 시각세포의 핵이 존 재하는 바깥핵층의 두께는 13.42(±2.16)μm였다. 이형 이중 원추세포 중 긴 원추세포는 26.42±1.7μm의 길이였으며 직경 은 5.12±0.6μm로 확인되었다. 짧은 원추세포는 16.82±1.1 μm의 길이였으며 직경은 긴 원추세포와 유사한 5.10±0.31 μm로 확인되었다. 간상세포는 길이 40.13±2.4μm였으며 직 경 3.95±0.4μm로 확인되었다. 시각세포의 수평단면을 확인 한 결과 이형 이중 원추세포가 규칙적인 배열로 인해 열 배열 (row-type)의 모자이크 패턴으로 나타났다. 긴 원추세포와 짧 은 원추세포가 하나의 쌍을 이루었으며 긴 원추세포는 내절이 절단되어 eosin에 염색되었으며 짧은 원추세포는 외절이 절단 되어 hematoxylin에 염색되었다. 전자현미경을 통해 미세구조 를 확인한 결과 내절과 외절로 구분되고 내절은 장타원형으로 확인되었다. 외절에는 디스크구조의 판상을 확인할 수 있었으 며 내절과 외절을 연결해주는 배상극(calyceal processes)들이 가장자리에 분포하고 있었다.
RefeRenCes
Audesirk, G., T. Audesirk and B.E. Byers. 2008. Biology : Life on Earth 8th edition. Prentice Hall, New Jersey, pp. 817-831.
Collins, B.A. and E.F. MacNichol. 1978. Triple cones found in reti- nas of 3 fish species. Cell Mol. Life Sci., 35: 106-108.
Donatti, L. and E. Fanta. 2007. Fine structure of the retinal pigment epithelium and cone of Antarctic fish Notohenia coriiceps Richardson in light and dark-conditions. Revista Brasileira de Zoologia, 24: 33-40.
Fernald, R.D. 1982. Cone mosaics in a teleost retina: No difference
between light and dark adapted states. Experientia, 38:
1337-1339.
Fernald, R.D. 1988. Aquatic adaptations in fish eyes. In: Atema, J., R.R. Fay, A.N. Popper and W.N. Tavolga(eds.), Sensory Biology of Aquatic Animals. Springer Verlag, New York, pp. 435-466.
Frank, R., R.M. Roland and S. Ulrich. 2001. Outer retinal fine structure of the garfish Belone belone(L.)(Belonidae, Tele- ostei) during light and dark adaptation-photoreceptors, cone patterns and densities. Acta Zool., 82: 89-105.
George, C.K. and K.C. Robert. 2001. Comparative anatomy of ver- tebrates. McGraw Hill, New York, pp. 387-454.
Hagedorn, M., A.F. Mack, B. Evans and R.D. Fernald. 1998. The embryogenesis of rod photoreceptors in the teleost fish reti- na, Haplochromis burtoni. Dev. Brain Res., 108: 217-227.
Hirt, B. and H.J. Wagner. 2005. The organization of the inner retina in a pure-rod deep-sea fish. Brain Behav. Evolut., 65: 157- 167.
Kim, I.S. 1997. Illustrated Encyclopedia of Fauna & Flora of Korea, Vol. 37, Freshwater Fishes. Ministry of Education, 629pp.
(in Korean)
Kim, I.S. and J.Y. Park. 2002. Freshwater fishes of Korea. Kyohak Publishing, Seoul, 467pp.(in Korean)
Kim, J.G., J.Y. Park and C.H. Kim. 2014. Visual cells in the retina of the aucha perch Coreoperca herzi Herzenstein, 1896(Pi- sces; Centropomidae) of Korea. J. Appl. Ichthyol., 30: 172- 174.
Lee, M.S. and J.W. Lim. 2005. Comparative study of retinal tissues of Genus Zacco and Genus Pseudogobio in Korean fish:
Possible relationship with prey and habitat. Korean J. Anat., 38: 103-110.
Lyall, A.H. 1956. Occurrence of triple and quadruple cones in the retina of the minnow(Phoxinus laevis). Nature, 177: 1086- 1087.
Lyall, A.H. 1957. Cone arrangements in teleost retinae. Quart. J.
Micr. Sci., 98: 189-201.
Marc, R.E. and H.G. Sperling. 1976. The chromatic organization of the goldfish cone mosaic. Vision Research, 16: 1211-1224.
Meer, H.J. van der. 1992. Constructional morphology of photore- ceptor patterns in percomorph fish. Acta Biotheor., 40: 51-
85.
Monica, R.L. 2001. Morphology of the eye and visual acuities in the settlement-intervals of some coral reef fishes(Labridae, Scaridae). Environ. Biol. Fish., 62: 365-378.
Nag, T.C. and J. Bhattacharjee. 2002. Retinal cytoarchitecture in some mountain-stream teleosts of India. Environ. Biol.
Fish., 63: 435-449.
Nicol, J.A.C. 1989. The eyes of fishes. Clarendon Press, Oxford, pp. 82-142.
Peter, B.M. and J.C.J. Joseph. 1996. Fishes : an introduction to ich- thyology. Prentice Hall, New Jersey, pp. 152-156.
Petr, T. and D.B. Swar. 2002. Cold water fisheries in the trans-Hi- malayan countries. FAO Fisheries Technical Paper. No.
431, Rome, 376pp.
Polyak, S. 1957. The vertebrate visual system. University of Chica- go Press, Chicago, pp. 1-178.
Rossetto, E.S., H. Dolder and I. Sazima. 1992. Doble cone mosaic pattern in the retina of larval and adult piranha, Serrasal- mus spilopleura. Experientia, 48: 597-599.
Sahly, I., E. Dufour, S. Cataldo, M. Vincent, B. Amel, P. Isabelle, P.
Elise, E. Amrit, C. Diane, A. Asadollah, E. Inga, L. Andrea, I. Maria, H. Jean-Pierre, W. Dominique, S. José-Alain, E.A. Aziz and P. Christine. 2012. Localization of Usher 1 proteins to the photoreceptor calyceal processes, which are absent from mice. J.C.B., 199: 381-399.
Thomas, J.L. and W.H. Craig. 2010. Ocular dimensions and cone photoreceptor topography in adult Nile tilapia Oreochromis niloticus. Environ. Biol. Fish., 88: 369-376.
You, M.J. and J.Y. Park. 2008. Comparative study of the retinal structure in two Korean endemic freshwater fishes, Zacco koreanus(Cyprinidae) and Pseudobagrus koreanus(Bag- ridae) based on their habitats. Korean J. Ichthyol., 20: 97- 104.(in Korean)
Yuko, N., O. Tadashi, T. Fumio and M. Toshiteru. 1997. Three-di- mensional reconstitution of cone arrangement on the spher- ical surface of the retina in the medaka eyes. Zool. Sci., 14:
795-801.
Yvette, W.K. 1980. Cone mosaics in a teleost retina: Changes during light and dark adaptation. Experientia, 36: 1371-1374.