Copyright ⓒ 2014, The Microbiological Society of Korea
Chondromyces crocatus 자실체 형성의 정량적 유도 방법 개발
이차율․신혜진․조경연*
호서대학교 생명공학과
Development of a Quantitative Induction Method for Chondromyces crocatus Fruiting Body Formation
Chayul Lee, Hyejin Shin, and Kungyun Cho*
Department of Biotechnology, Hoseo University, Asan 336-795, Republic of Korea (Received July 14, 2014 / Accepted July 26, 2014)
We have developed a method for the preparation of dispersed cell suspensions of Chondromyces crocatus, which is essential for quantitative studies of fruiting body formation. Cells of C. crocatus have a tendency to aggregate in liquid, hindering quantitative studies. However, cells grown on casitone-yeast extract agar plates, containing 3%
agar, allowed the preparation of well-dispersed cell suspensions. Cell suspensions at a concentration of 2 × 108 cells/ml, obtained by using this method, developed typical C. crocatus fruiting bodies when placed as 20 μl spots on agar plates with no nutrient supplementation. The addition of nutrients such as casitone altered or inhibited fruiting body formation. Fruiting body branch formation increased with increasing agar content. Under optimum conditions, the formation of fruiting body structure in C. crocatus KYC2823 was completed within 24 h.
Keywords: Chondromyces crocatus, fruiting body, myxobacteria
*For correspondence. E-mail: [email protected]; Tel.: +82-41-540- 5627; Fax: +82-41-548-6231
점액세균(myxobacteria)은 다세포 자실체를 형성하는 세균으로 잘 알려져 있다(Reichenbach, 2005; Shimkets et al., 2006; Kaiser et al., 2010). 점액세균은 영양분이 풍부한 상태에서 간균 형태의 개별 세포들이 다른 세균들과 마찬가지로 분열하여 영양생장을 한 다. 하지만 영양분이 결핍된 환경에서는 수십만 마리의 점액세균 들이 협력하여 작은 버섯과 같은 모양의 자실체 구조를 형성하고, 각각의 세포들이 자실체 구조 안에서 구형 또는 타원형 모양의 포자로 변화됨으로써 성숙한 자실체를 형성한다. 영양분이 다시 풍부해지는 환경이 되면 자실체 내의 포자들이 동시에 발아하여 다시 집단의 영양세포로 변하여 생장하게 된다. 점액세균에 의한 다세포 자실체의 형성은 수많은 유전자가 관여하는 매우 정교하 고 체계적인 과정이다. 따라서 점액세균은 세균의 발달을 연구하 는 주요 재료의 하나로 이용되어 왔다(Kaiser et al., 2010).
점액세균은 종에 따라서 고유한 형태의 다세포 자실체를 형성 한다(Dawid, 2000; Reichenbach, 2005; Shimkets et al., 2006).
Chondromyces crocatus는 점액세균 종류 중 가장 정교하고 복 잡한 자실체를 형성하는 종의 하나로 흡사 나무에 큰 열매가 달려 있는 형태의 자실체를 형성한다(Grilione and Pangborn, 1975).
하지만 자실체 형성에 관한 연구는 대부분 단순한 반구형 자실
체를 형성하는 Myxococcus xanthus를 대상으로 이루어졌는데, 이는 M. xanthus가 상대적으로 분리 및 배양이 쉬우며, 액체 및 고체 배지에서 빠르게 성장할 뿐만 아니라 분산되어 성장하는 균주가 존재하여 정량적인 분석과 유전자 조작이 가능하기 때문 이다(Dworkin, 1962; Wall et al., 1999). 반면에, C. crocatus의 자실체 형성에 대한 연구는 거의 이루어지지 않았는데, 이는 순 수 분리하기가 매우 어려워 전 세계적으로 극소수 균주만이 순 수 분리되었으며, 액체배지에서 잘 성장하지 않거나 성장하더라 도 매우 느리게 배양용기의 표면에 세포덩어리 형태로 성장하기 때문이다(Hesseltine and Fennell, 1955; McCurdy, 1964; Nellis and Garner, 1964). 덩어리로 성장한 세포들은 세포의 상태도 알 수 없고, 균일하게 현탁되지도 않기 때문에 정량적이며 재현성 있는 자실체 형성 연구를 곤란하게 한다.
본 연구에서는 국내에서 순수 분리한 C. crocatus KYC2823 (Lee et al., 2009)을 대상으로 세포를 균일하게 현탁할 수 있는 방법을 개발하였으며, 이를 바탕으로 C. crocatus의 정량적 자실 체 형성 조건을 조사하였다.
재료 및 방법 사용 균주, 배지, 배양 조건
C. crocatus KYC2823은 본 연구팀이 국내에서 순수 분리한
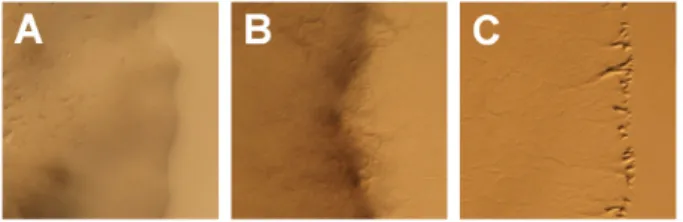
Fig. 1. Growth of C. crocatus KYC2823 on agar plates. C. crocatus KYC2823 was inoculated on CY medium containing 0.5% (A), 1.5%
(B), and 3.0% (C) agar and incubated at 32℃ for 5 days. Bar, 1 mm.
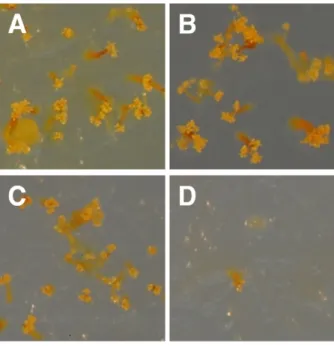
Fig. 2. Effects of cell density on C. crocatus KYC2823 fruiting body development. Twenty-microliter spots of cells at concentrations of 1 × 109 cells/ml (A, E), 2 × 108 cells/ml (B, F), 4 × 107 cells/ml (C, G), and 8 × 106 cells/ml (D, H), which were cultured on CY plates, were placed on ST21 (A–D) and WC (E–H) plates containing 3.0% agar and incubated at 32℃ for 2 days. Bar, 0.2 mm.
균주이다(Lee et al., 2009). C. crocatus의 일반적인 배양에는 0.3% (w/v) casitone, 0.1% yeast extract, 0.1% CaCl2․ 2H2O, 3.0%
agar를 함유한 casitone-yeast extract (CY) 평판배지를 사용하였 으며, 자실체 형성을 위해서는 mineral salts (ST21) 배지와 water-calcium (WC) 배지를 사용하였다(Shimkets et al., 2006).
배양은 32℃에서 이루어졌다. ST21 배지는 0.1% K2HPO4, 0.002%
yeast extract, 0.1% KNO3, 0.1% MgSO4·7H2O, 0.02% FeCl3, 0.01% MnSO4·7H2O, 0.1% 미량원소용액을 함유하고 있다. WC 배지는 10 mM 3-[N-morpholino]propanesulfonic acid (MOPS, pH 7.6), 0.1% CaCl2⋅2H2O을 함유하고 있다. 일반적인 평판 배지의 제조를 위해서는 한천(BD, USA)을 사용하였으며 칼슘 이온 농도가 자실체 형성에 미치는 영향을 위해서는 agarose (Sigma, USA)를 사용하였다.
자실체 형성 유도
C. crocatus KYC2823을 3.0% 한천을 함유한 CY 배지에서 5일 동안 배양한 다음 세포들을 회수하여 멸균 증류수에 현탁하 였다. 원심분리하여 상등액을 제거하고, 다시 멸균 증류수에 현 탁하여 세포의 농도가 2 × 108 cells/ml가 되도록 하였다. 20 μl 를 배지 위에 올려놓고 32℃에서 배양하였다. 세포 수는 세균계
수판을 이용하여 측정하였다.
형태학적 특성조사
자실체의 관찰에는 Nikon SMZ1000 입체현미경을 사용하였으 며, Nikon Digital sight DS-Fi1 카메라를 사용하여 촬영하였다.
결 과
한천 농도가 C. crocatus KYC2823의 성장 및 분산에 미치는 효과
CY 한천 배지는 대부분의 용균성 점액세균이 성장할 수 있는 배지이다. C. crocatus KYC2823은 CY 액체배지에서는 성장하 지 않고 배양용기 내부 표면에 엉겨 붙지만, CY 한천배지에서는 잘 성장한다. 하지만 1.5% 한천을 함유한 CY 배지 상에서 배양 할 경우 세포 집락이 한천을 파고들면서 성장하였으며(Fig. 1B), 물에 현탁할 경우 세포들이 덩어리지는 현상을 보였다. 정량적 이고 재현성 있는 자실체 형성 연구를 위해서는 분산된 세포의 확보가 필수적이다. 따라서 다양한 배지를 대상으로 C. crocatus KYC2823의 성장 여부를 조사하여 보았지만 대부분 CY 배지에 비해 성장속도가 느리거나 세포들이 덩어리지는 정도가 더 심하 였다. 한 예로, C. crocatus KYC2823은 M. xanthus의 배양에 많 이 사용되는 CYE 배지(Hagen et al., 1978)에서는 성장하지 못 하였다.
CY 배지를 대상으로 배양조건을 조사하던 중, 한천 농도를 다르게 하면 어떠한 형태로 성장하는지를 알아보기 위해 CY 배 지의 한천 농도를 다르게 하여 배양한 결과, 3.0%의 한천이 함유 된 CY 평판배지에서 C. crocatus KYC2823 세포들이 모두 배지 표면에서 얇은 층으로 퍼져 성장함을 관찰할 수 있었다(Fig. 1C).
반면에, 0.5% 한천이 함유된 배지에서는 세포들이 1.5% 한천을 함유한 배지에서 보다도 더 심하게 한천 내부로 파고드는 모습
Fig. 3. Effects of agar content on C. crocatus KYC2823 fruiting body development. Twenty-microliter spots of cells at a concentration of 2 × 108 cells/ml, which were cultured on CY plates, were placed on WC plates containing 0.5% (A), 1.5% (B), and 3.0% (C) agar and incubated at 32℃ for 2 days. Bar, 0.2 mm.
Fig. 4. Effects of calcium ion content on C. crocatus KYC2823 fruiting body development. Twenty-microliter spots of cells at a concentration of 2 × 108 cells/ml, which were cultured on CY plates, were placed on WC plates containing 3.0% agarose and 0 mM (A), 6.8 mM (B), 34 mM (C), and 136 mM (D) CaCl2 and incubated at 32℃ for 3 days. Bar, 0.2 mm.
을 관찰할 수 있었다. 또한, 0.5% 한천이 함유된 배지에서 배양 된 세포들은 1.5% 한천을 함유한 배지에서 배양한 세포들과 마 찬가지로 증류수에 균일하게 현탁되지 않았으나, 3.0% 한천을 함유한 배지에서 성장한 세포들은 한천배지 표면에서 성장한 까 닭에 증류수에 균일하게 현탁할 수 있었다. 현탁한 세포들은 자 실체 형성 배지에서 전형적인 형태의 자실체를 형성하였다. 따 라서 본 발견으로 인해 C. crocatus 세포들이 균일하게 분산된 세포현탁액을 제조할 수 있게 되었으며, 이를 사용하여 정량적 이면서도 재현성 있는 자실체 형성 실험이 가능하게 되었다.
세포농도가 자실체 형성에 미치는 영향
점액세균이 자실체를 형성하기 위해서는 일정 수 이상의 세포 농도가 필요하다. 자실체 형성기작 연구의 모델균주인 M. xanthus DZ2의 경우에는 일반적으로 세포를 5 × 109 cells/ml로 농축한 후, 20 μl를 자실체 유도 배지 상에 올려놓고 배양함으로써 자실 체 형성을 유도한다(Cho and Zusman, 1999). C. crocatus의 자 실체 형성을 위해 필요한 세포농도를 알아보기 위해 C. crocatus KYC2823을 3.0% 한천을 함유한 CY 배지에서 5일간 배양한 후, 증류수에 1 × 109 cells/ml로 현탁하였다. 그리고 20 μl (2 × 107 cells)를 ST21 배지와 WC 배지에 올려놓고 32℃에서 배양하였다.
또 세포현탁액을 5배(2 × 108 cells/ml), 25배(4 × 107 cells/ml), 125배(8 × 106 cells/ml)로 희석하여 20 μl씩을 ST21 배지와 WC 배지에 올려놓고 배양하였다. ST21 배지는 C. crocatus를 순수분리하는 과정에서 자실체 유도를 위해 사용하였던 배지이 며 WC 배지는 일반적인 점액세균의 자실체 유도를 위해 사용하 는 배지이다. 8 × 106 cells/ml에서는 ST21와 WC 배지에서 모두 자실체를 형성하지 않았으며, 4 × 107 cells/ml의 경우에는 ST21 에서는 정상적인 자실체 형태를 형성하지 못하였지만 WC에서는 매우 적은 수의 자실체를 형성하였다(Fig. 2). 2 × 108 cells/ml과 1 × 109 cells/ml에서는 모두 자실체를 형성하였는데, 세포농도 가 높은 경우에 더 많은 수의 자실체를 형성하였다. WC 배지에 서는 정상적인 자실체를 형성하였으나, ST21에서는 대부분의 자실체에서 포자낭 덩어리 위에 새로운 가지가 뿔처럼 솟아나는 모습을 보였으며, 일반적인 C. crocatus의 야생형 자실체 모양에 서 벗어나 있었다(Fig. 2). 따라서 C. crocatus KYC2823의 자실 체 형성에는 ST21 배지보다는 WC 배지가 더 적합하며, 세포의 농도는 2×108 cells/ml 이상이 적합한 것으로 나타났다.
한천 농도가 자실체 형성에 미치는 영향
전술한 바와 같이 한천 농도는 C. crocatus KYC2823의 영양 성장 형태에 영향을 미친다. 한천의 농도가 자실체 형성에는 어 떤 영향을 미치는지 조사하기 위하여 20 μl의 2 × 108 cells/ml C. crocatus KYC2823을 0.5%, 1.5%, 또는 3.0% 한천을 함유한 WC 배지 상에 올려놓고 2일 동안 배양한 후, 자실체 형성 양상 을 조사하였다. 0.5% 한천이 함유된 배지에서는 대부분의 자실 체들이 가지가 전혀 없는 작은 자실체를 많은 수로 형성한 반면(
Fig. 3A), 3.0% 한천을 함유된 WC 배지에서는 여러 갈래로 가 지 친 큰 자실체를 적은 수 형성하였다(Fig. 3C). 1.5% 한천을 함 유한 WC 배지에서는 가지 없는 자실체와 가지 친 자실체들이 섞여 있었다(Fig. 3B). 따라서 C. crocatus KYC2823는 한천의 농도의 농도가 높아질수록 자실체 내 가지 형성이 증가하는 것 으로 나타났다.
세포의 상태와 자실체 형성
자실체 형성에 요구되는 세포의 상태를 알아보기 위하여 CY 배지에서 다양한 기간 동안 배양한 세포들을 대상으로 자실체 형성을 조사하였다. 그 결과, 3–5일 배양한 세포들은 정상적인 자실체를 형성하였으나, 6일 이상 배양한 세포들은 정상적인 자 실체를 형성하지 못하였다.
칼슘이온 및 casitone이 자실체 형성에 미치는 영향
WC 배지는 6.8 mM 칼슘이온을 함유하고 있으므로 칼슘이 온이 C. crocatus KYC2823 균주의 자실체 형성에 어떠한 영향
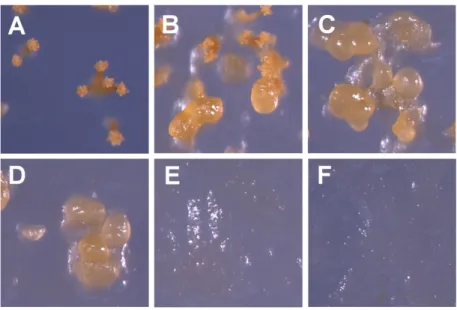
Fig. 5. Effects of casitone content on C. crocatus KYC2823 fruiting body development. Twenty-microliter spots of cells at a concentration of 2
× 108 cells/ml, which were cultured on CY plates, were placed on WC plates containing 3.0% agar and 0% (A), 0.01% (B), 0.02% (C), 0.08%
(D), 0.16% (E), and 0.34% (F) casitone and incubated at 32℃ for 2 days. Bar, 0.2 mm.
Fig. 6. Fruiting body development of C. crocatus KYC2823. Twenty-microliter spots of cells at a concentration of 2 × 108 cells/ml, which were cultured on CY plates, were placed on WC plates containing 3.0% agar and incubated at 32℃. Bar, 0.1 mm.
이 있는지를 알아보았다. 이를 위해 일반적인 한천에 포함되어 있을지 모르는 극미량 원소들을 배제하기 위해 한천 대신 고순 도 아가로스를 사용하였다. 3.0% 아가로스 배지에 WC 배지보 다 칼슘이온이 5배(34 mM), 20배(136 mM) 많은 배지와 칼슘이 온이 전혀 없는 배지에서 자실체 형성을 조사하였다. 그 결과, 칼 슘이온이 136 mM 함유된 배지에서는 자실체 형성이 억제된 반면 (Fig. 4D), 칼슘이온이 전혀 없는 경우와 6.8 mM을 함유한 경우 에서는 정상적인 자실체를 형성하였다(Figs. 4A and 4B).
일반적으로 점액세균은 영양분이 결핍된 상태에서 자실체 형 성을 개시하기 때문에 일정 양 이상의 영양분이 존재하는 경우 에는 자실체를 형성하지 않는다. 하지만 소량의 영양분은 자실 체 형성에 도움을 주므로 점액세균 M. xanthus DZ2 균주의 자실 체 형성 유도 배지인 CF에는 0.015% casitone이 함유되어 있다 (Shimkets et al., 2006).
KYC2823 균주에 대해 자실체 형성을 억제하는 영양분의 농 도를 알아보기 위하여 WC 배지에 0.01%, 0.02%, 0.04%, 0.08%, 0.16%, 0.34%의 casitone을 첨가하고 자실체 형성 여부
를 조사하였다. 그 결과, casitone이 전혀 존재하지 않는 경우에 서는 정상적인 자실체를 형성하는 반면에 casitone이 0.01%만 존재하여도 정상적인 자실체가 아닌 부분적으로 비정상적인 자 실체를 형성함을 보였고, 0.02% 이상의 casitone을 함유한 배지 에서는 전혀 자실체 형태를 띠지 않는 세포 덩어리가 형성되었 다(Fig. 5).
C. crocatus KYC2823에 의한 자실체 형성 단계 관찰 최적 상태에서의 자실체 형성 과정을 관찰하기 위하여 2×108 cells/ml의 C. crocatus KYC2823 세포현탁액 20 μl를 3.0% 한 천을 함유한 WC 배지에 올려놓고 자실체 형성과정을 관찰하였 다. 배양 시작 후 약 12시간이 되었을 때 세포의 집단으로 이루 어진 끝이 뭉툭한 기둥이 형성되었으며, 16시간 이후에는 끝에 구가 있는 기둥이 형성되었고, 18시간 이후에는 구가 갈라져 여 러 개의 포자낭이 생성되었다. 가지의 분지는 16시간 이후에 이 루어졌다. 이런 과정을 통해 24시간 이전에 자실체의 외형이 완 성되었다(Fig. 6).
고 찰
C. crocatus는 다른 점액세균 종류에 비해 복잡하고 아름다운 자 실체를 형성하는 까닭에 학문적 관심이 많다(Hesseltine and Fennell, 1955; Grilione and Pangborn, 1975; Kaiser et al., 2010). 하지 만 세포들이 분산되지 않는 특성으로 인해 정량적인 분석이 불 가능하여 자실체 형성 연구에 큰 진전이 없었다. 본 연구에서는 3.0% 한천이 함유된 평판배지에서 배양한 C. crocatus 세포들이 액체에서 균일하게 분산되는 특성을 이용하여 세포현탁액을 준 비함으로써 C. crocatus의 정량적 자실체 형성 연구를 실시할 수 있는 방법을 개발하였다.
M. xanthus를 대상으로 한 연구에 의하면 점액세균은 A-운동 성(개별 세포의 독립 활주운동)과 S-운동성(집단 내에서의 활주 운동)의 두 활주운동기구를 지니고 있는데(Cho, 2002; Mauriello et al., 2010), 0.5% 한천에서는 A-운동성이 억제되는 반면 S-운 동성이 활발해지며, 3.0% 한천에서는 S-운동성이 억제되고 A- 운동성이 활발해지는 것으로 알려져 있다(Shi and Zusman, 1993). S-운동성은 type IV pili가 신장되어 다른 세포 표면 또는 주변 물질에 부착한 다음 수축에 의해 세포 이동이 이루어진다 (Mauriello et al., 2010). S-운동성을 지닌 세포들은 세포들끼리 서로 부착하므로 덩어리 형태로 존재한다. 반면에 S-운동성 관 여 유전자가 불활성화된 돌연변이의 경우에는 대부분 완전하게 분산되어 성장한다(Shimkets, 1986; Wall et al., 1999). C.
crocatus의 경우에도 A와 S-운동성을 지닌 것으로 관찰되는데, 한천 농도가 낮은 배지에서는 S-운동성이 활성화되어 세포들이 응집되는 반면, 한천 농도가 높은 배지에서는 S-운동성이 억제 되어 세포들이 서로 응집되지 않아 물에 현탁할 경우 균일하게 분산되는 것으로 사료된다.
C. crocatus의 자실체 형성을 위해서는 세포 농도가 2 × 108 cells/ml 이상인 젊은 세포들이 필수적인 것으로 분석되었으며 (Fig. 2), 영양분이 전혀 없는 상황에서 자실체를 가장 잘 형성하 였다. Casitone과 같은 영양분이 있는 경우에는 자실체 형성이 억제되었다(Fig. 5). M. xanthus의 경우에도 충분한 영양분이 존 재하는 환경에서는 영양생장만 하고 자실체를 형성하지 않으며 필요한 아미노산 중 하나라도 농도가 0.01 mM 이하로 떨어질 때 자실체 형성이 이루어진다(Hemphill and Zahler, 1968). 특정 아 미노산들은 자실체 형성에 서로 다른 효과를 미치는데(Hemphill and Zahler, 1968; Cho and Zusman, 1999), 후속 연구를 통해 이 러한 아미노산들이 C. crocatus의 자실체 형성에는 어떠한 영향 을 미치는지 조사될 것이다. 자실체 형성은 한천의 농도에 영향 을 받아 한천 함량이 높을수록 분지가 많아지고 자실체의 크기 가 커지는 반면 한천 함량이 낮은 환경에서는 한 점으로 모인 세 포 수가 적어 가지가 분지되지 않는 작은 자실체를 생성하였다 (Fig. 3). 이는 C. crocatus가 고체 표면 위를 활주운동으로 움직 이는데, 단단한 표면에서는 넓은 지역의 세포들이 한 점으로 모 여 분지가 많은 큰 자실체를 형성하는 반면, 무른 표면에서의 자 실체 유도는 더 좁은 지역의 세포들이 모여 분지수가 적은 작은 자실체를 형성하는 것으로 추정되어진다.
자실체 형성을 위한 구조물의 형성은 자실체 형성 유도 시작
시간으로부터 약 12시간 이후에 시작되었는데, 먼저 세포의 집 단으로 구성된 기둥이 생성되고, 머리 부분에 구가 발달한 다음, 구가 갈라져 포자낭이 형성됨으로써 24시간 이내에 자실체 외형 을 완성하였다(Fig. 6). 반구형의 단순한 형태의 자실체를 형성 하는 M. xanthus의 경우에는 빠르면 24시간 내에 자실체 구조를 형성한다(Shimkets, 1990). C. crocatus은 M. xanthus에 비해 성 장이 느리며 구조도 복잡하므로 자실체 형성에 오랜 시간이 걸 릴 것으로 예상되었다. 하지만 이러한 선입관과는 달리 조건이 충족되면 M. xanthus와 마찬가지로 24시간 내로 자실체 구조를 형성하는 것으로 나타났다. 본 연구에서 개발한 세포의 분산, 이 를 이용한 정량적인 자실체 형성 유도, 그리고 자실체 형성을 위 한 최적 조건은 향후 C. crocatus의 자실체 형성 연구에 중요한 기반이 될 것으로 기대한다.
적 요
Chondromyces crocatus의 자실체 형성에 대한 정량적 연구 를 위하여 필수적인 분산된 세포현탁액 제조방법을 개발하였다.
C. crocatus 세포들은 액체에서 서로 응집하는 특성이 있어 정량 적 연구가 어렵다. 하지만 3% 한천을 함유한 casitone-yeast extract 평판배지에서 배양한 세포들은 물에 잘 분산되어 분산된 세포현탁액을 제조할 수 있었다. 이렇게 제조한 2 × 108 cells/ml 농도의 세포현탁액 20 μl를 영양분이 고갈된 평판배지 위에 올 려놓고 배양하였을 때 C. crocatus 특유의 자실체를 형성하였다.
Casitone과 같은 영양분을 첨가하였을 때는 자실체 형성이 저해 되거나 변형되었으며, 한천의 농도가 높을수록 자실체의 가지 수가 증가되었다. 조사된 최적조건에서 C. crocatus KYC2823 은 24시간 내로 자실체 구조를 완성하였다.
감사의 말
이 논문은 2012년도 호서대학교의 재원으로 학술연구비 지원 을 받아 수행된 연구이며 이에 감사드립니다(2012-0253).
References
Cho, K. 2002. Bacterial gliding motility. Kor. J. Microbiol. Biotechnol.
30, 199–205.
Cho, K. and Zusman, D.R. 1999. AsgD, a new two-component regulator required for A-signalling and nutrient sensing during early development of Myxococcus xanthus. Mol. Microbiol. 34, 268–281.
Dawid, W. 2000. Biology and global distribution of myxobacteria in soil.
FEMS Microbiol. Rev. 24, 403–427.
Dworkin, M. 1962. Nutritional requirements for vegetative growth of Myxococcus xanthus. J. Bacteriol. 84, 250–257.
Grilione, P.L. and Pangborn, J. 1975. Scanning electron microscopy of fruiting body formation by myxobacteria. J. Bacteriol. 124, 1558–
1565.
Hagen, D.C., Bretscher, A.P., and Kaiser, D. 1978. Synergism between morphogenetic mutants of Myxococcus xanthus. Dev. Biol. 64, 284–
296.
Hemphill, H.E. and Zahler, S.A. 1968. Nutritional induction and
suppression of fruiting in Myxococcus xanthus FBa. J. Bacteriol. 95, 1018–1023.
Hesseltine, C.W. and Fennell, D.I. 1955. Growth of Chondromyces crocatus in pure culture. Nature 175, 213–214.
Kaiser, D., Robinson, M., and Kroos, L. 2010. Myxobacteria, polarity, and multicellular morphogenesis. Cold Spring Harb. Perspect. Biol.
2, a000380.
Lee, C., Hyun, H., Kim, D., and Cho, K. 2009. Isolation of Chondromyces crocatus in pure culture. Kor. J. Microbiol. Biotechnol. 37, 316–321.
Mauriello, E.M., T. Mignot, T., Yang, Z., and Zusman, D.R. 2010.
Gliding motility revisited: how do the myxobacteria move without flagella? Microbiol. Mol. Biol. Rev. 74, 229–249.
McCurdy, H.D.Jr. 1964. Growth of Chondromyces crocatus in pure culture. Can. J. Microbiol. 10, 935–936.
Nellis, L.F. and Garner, H.R. 1964. Methods for isolation and purification of Chondromyces. J. Bacteriol. 87, 230–231.
Reichenbach, H. 2005. Myxococcales. pp. 1059–1144. In Brenner, D.J.,
Krieg, N.R., Staley, J.T., and Garrity, G.M. (eds.), Bergey's Manual of Systematic Bacteriology, 2nd ed. Bergey's Manual Trust, East Lansing, MI, USA.
Shi, W. and Zusman, D.R. 1993. The two motility systems of Myxococcus xanthus show different selective advantages on various surfaces.
Proc. Natl. Acad. Sci. USA 90, 3378–3382.
Shimkets, L.J. 1986. Correlation of energy-dependent cell cohesion with social motility in Myxococcus xanthus. J. Bacteriol. 166, 837–841.
Shimkets, L.J. 1990. Social and developmental biology of the myxobacteria. Microbiol. Rev. 54, 473–501.
Shimkets, L.J., Dworkin, M., and Reichenbach, H. 2006. The myxobacteria. Prokaryotes 7, 31–115.
Wall, D., Kolenbrander, P.E., and Kaiser, D. 1999. The Myxococcus xanthus pilQ (sglA) gene encodes a secretin homolog required for type IV pilus biogenesis, social motility, and development. J.
Bacteriol. 181, 24–33.