200 책임저자:김송자, 314-701, 충남 공주시 신관동 182
공주대학교 자연과학대학 생명과학과 Tel: 041-850-8507, Fax: 041-850-0927 E-mail: [email protected]
접수일:2011년 7월 6일, 1차 수정일:2011년 7월 12일, 2차 수정일:2011년 7월 15일, 게재승인일:2011년 7월 18일
Correspondence to:Song-Ja Kim
Department of Biological Sciences, College of Natural Sciences, Kongju National University, 182, Singwan-dong, Gongju 314-701, Korea Tel: +82-41-850-8507, Fax: +82-41-850-0927
E-mail: [email protected]
Gallotannin (GT)에 의한 MDA-MB-231 유방암 세포의 PI3-kinase/p38 MAP Kinase 신호전달 경로를 통한 세포사멸과 염증반응 조절
공주대학교 자연과학대학 생명과학과
유선미ㆍ김송자
Gallotannin (GT) Regulates Apoptosis and Inflammation via PI3-kinase/p38 MAP Kinase Pathway in MDA-MB-231 Human Breast Cancer Cells
Seon-Mi Yu and Song-Ja Kim
Department of Biological Sciences, College of Natural Sciences, Kongju National University, Gongju 314-701, Korea
Gallotannin (GT) is a type of tannic acid derived from plant polyphenol, it is usually involved in plant defense mechanism. GT has been used in medical agents such as anti-cancer, anti-inflammatory, anti-virial, anti-vacterial and anti-parastic in a wide range of cells. We have investigated the effects of GT on apoptosis and inflammation in MDA-MB-231 human breast cancer cells. Treatment of GT induced apoptosis and reduced proliferation as detected by MTT assay, and increased the expression of active caspase-3 as determined by Western blot analysis. Also, treatment of GT increased the expression of cyclooxygenase-2 (COX-2) and the production of prostaglandin E
2(PGE
2) in a dose- and time-dependent manner as determined by Western blot analysis and PGE
2assay. We confirmed that GT dramatically caused phosphorylation of Akt and p38 kinase. Inhibition of PI3-kinase and p38 kinase with LY294002 and SB203580 prevented GT-induced apoptosis and COX-2 expression. Our results suggested that GT regulates apoptosis and COX-2 expression through PI3-kinase and p38 kinase pathway in MDA-MB-231 human breast cancer cells. (Cancer Prev Res 16, 200-209, 2011)
Key Words: Gallotannin (GT), Apoptosis, Cyclooxygenase-2 (COX-2), PI3-kinase, p38 kinase
서 론
우리나라에서 암은 사망 원인 중 첫 번째로 높은 빈도 를 나타내는 질병이다. 암은 정상적인 세포의 성장과 다 르게 일반적인 규칙을 따르지 않고 비정상정인 성장을 나타냄으로써 정상적인 세포의 파괴를 가져오게 된다.
이 중 유방암은 전세계적으로 여성에게 일어나는 암 중
첫 번째로 발병률이 높은 것으로 알려져 있다.
1)암의 전 이는 암의 치료를 어렵게 할 뿐만 아니라 암 환자의 생 존율을 낮게 만든다. 그러므로 암의 초기 발생을 막는 것이 가장 중요하며, 암 발생 이후에 각 진행단계를 차단 시켜 치료를 용이하게 하는 것 또한 중요하다.
2)MDA- MB-231 51세 환자로부터 처음 추출한 유방암 세포로서 p53단백질에 돌연변이가 일어난 세포이다.
COX는 arachidonic acid (AA)를 prostanoids로 바꾸는 일
련의 과정 중 반응속도를 조절하는 주요 효소이며, 지방 산 호르몬인 PGE
2의 생산을 촉진하는 역할을 한다. COX 는 대표적으로 2종류의 아형이 존재하는데, 그 중 하나 인 COX-1은 대부분의 정상 조직에서 발현되어 세포나 조직의 항상성을 유지하는 기능을 하는 것으로 알려져 있다. 한편, COX-2는 일반적으로 cytokine, lipopolysaccha- ride (LPS), Growth factor, oncogene 등과 같은 여러 자극에 반응함으로써
3∼5)발현량이 증가하며 특히 염증이나 종 양이 발생된 조직에서의 발현이 증가된다.
6)통증과 염증 을 일으키는 것으로 알려진 PGE
2는 암을 비롯한 여러 질 환들과 관련되어 있으며, PGE
2는 특히 혈관 형성을 촉진 하여 종양의 형성, 전이, 침습에 관여한다.
7,8)최근 다양한 천연물이 항암효과를 보인다는 연구결과 들이 발표됨으로써 천연물을 사용한 연구가 활발히 이 루어지고 있으며, 이와 같은 천연물은 부작용을 일으키 지 않으므로 암 치료에 있어서 효과적인 물질로 알려져 있다.
9)이러한 천연물에 속하는 GT는 다양한 식물, 과일, 음 식에 광범위하게 존재하며 가수분해형으로 수산기를 가 진 페놀을 합성하며 gallic acid라고도 불린다. 이러한 가 수분해형 탄닌은 의학적인 용도로써 항암,
10,11)함염
증,
12,13)항바이러스,
14,15)항박테리아
16)등의 기능으로 사
용된다.
PI3-kinase및 p38 kinase 신호 경로는 세포사멸, 세포분 화, 물질대사, 염증반응을 포함하는 세포 내 다양한 반응 을 조절하는데 관여한다.
17∼19)PI3K/Akt는 PIP
3에 의해 조절되는 serine-threonine protein kinase로써 섬유모세포, 상피세포, 신경세포 등의 다양한 세포에서 생존과 관련 되어 있는 신호전달 경로를 활성화시킨다.
20)또한 췌장 암세포를 포함한 다양한 암세포에서의 Akt의 과발현이 보고되었으며, 이러한 Akt의 과발현은 세포사멸에 덜 민 감하게 반응하도록 세포를 조절한다.
21,22)MAP kinases인 extracellular signal-regulated protein kinase (ERK), p38 MAP kinase, c-Jun N-terminal kinase (JNK)는 포유동물에서 세포 분열, 이동, 물질대사 및 세포사멸 등의 주요한 반응을 매개하는 신호전달경로로 알려져 있다.
23,24)따라서 본 연구에서는 MDA-MB-231 유방암 세포에 GT를 처리하여 유방암세포의 염증반응 및 세포사멸에 미치는 GT의 효과 및 신호 전달 경로를 규명하기 위한 연구를 수행하였다.
재료 및 방법
1. Cell culture실험에 사용된 인체 유방암 세포인 MDA-MB-231 유방 암세포는 American Type Culture Collection (ATCC, Rockville, MD, USA)에서 구입하였다. 세포는 37
oC, 5% CO
2in- cubator에서 RPMI-1640 (GibcoBRL, Gaithersburg, MD, USA) 에 10% fetal bovine serum (GibcoBRL), 50μg/ml의 strepto- mycin (Sigma, MO, saint louis, USA), 50 units/ml의 Penicillin (Sigma)이 함유된 RPMI-1640 (GibcoBRL)에 섞은 후 배양 접시에 1.5×10
5cells/cm
2의 밀도로 배양하였다. 배지는 2
∼3일마다 교환하였으며, 배양접시의 세포밀도가 약 80%가 되었을 때 본 연구에 사용된 시약을 처리하였다.
2. Western blot analysis
실험에 사용된 세포는 단백질의 발현 양상을 확인하 기 위하여 50 mM Tris-HCl, pH 7.4, 150 mM NaCl, 1%
nonidet P-40, 0.1% sodium dodecylsulfate가 함유된 cell lysis buffer에 단백질 분해효소 억제제[10 μg/ml leupeptin, 10 μg/ml pepstatin A, 10μg/ml aprotinin, 1 mM 4-(2-amino- ethyl) benzensulfonyl fluoride]와 인산 분해효소 억제제[1 mM NaF, 1 mM Na
2VO
4]를 첨가하여 세포 단백질을 추출 하였다. 추출된 단백질은 SDS-polyacrylamide gel에 전기 영동하여 단백질을 크기별로 분리하였으며 니트로셀룰 로오스 막으로 이동시켰다. 그 후 각 단백질 발현의 분석 을 위하여 각 단백질에 특이적인 항체인 COX-2 (Cayman Chemical, Ann Arbor, MI, USA), caspase-3 (Cell Signaling Technology, Denvers, MA, USA), pp38 (Cell Signaling Technology), pAkt (Cell Signaling Technology), and β-actin (Santa Cruz, CA, USA)을 사용하였다.
3. Cell proliferation assay
GT의 농도 증가에 따른 세포의 증식에 미치는 효과를 알아보기 위해 MTT assay를 실시하였다. 96 well plate에 유방암 세포를 1×10
5cells/well로 분주하여 37
oC에서 24시 간 동안 배양 한 후 MTT reagent I을 10μl/well씩 넣고 4시간 동안 배양한 후, 다시 solubilization solution인 MTT reagent II을 넣어 12시간 더 배양을 하였다. 후에 ELISA reader를 사용하여 흡광도(550∼600 nm) 값을 측정하였 다.
4. PGE2 assay
COX-2에 의한 PGE
2의 농도 변화를 assay kit (Assay de-
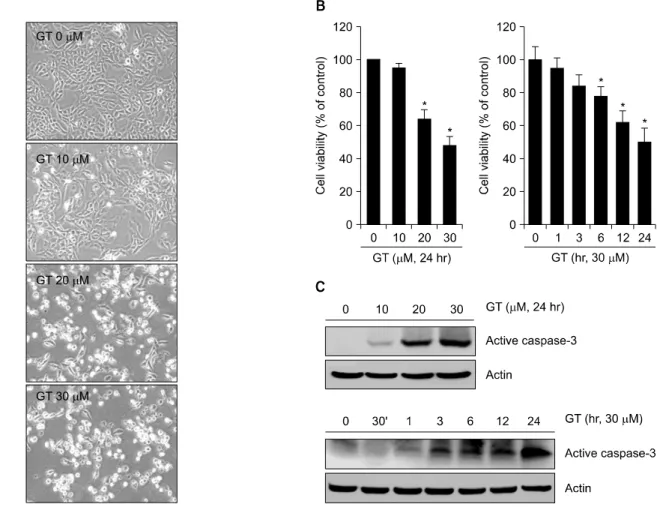
Fig. 1. GT induces apoptosis in MDA-MB-231 cells. MDA-MB-231 cells were untreated (control) or treated with the various concentrations of GT for 24 hr. Cells photographs were taken using phase contrast microscope (magnification, x100) (A).
MDA-MB-231 cells were untreated (control) or treated with the various concentrations of GT for 24 hr or with the 30μM for the indicated time periods. Cell viability was determined by MTT assay (B). MDA-MB-231 cells were untreated (control) or treated with the various concentrations of GT for 24 hr or with the 30μM of GT for the indicated time periods. Expressions of active caspase-3 and actin were determined by Western blot analysis (C). The data in B represent mean values and SD (n=6), and in C represent results of a typical experiment from four independent experiments with similar results. *p<0.05 compared with untreated cells.
sign, USA)를 사용하여 enzyme-linked immunoabsorbent as- say (ELISA) 방법으로 제조사의 방법에 따라 정량하였다.
96 well plate에 세포를 분주하여 24시간 배양한 후 gallo- tannin을 여러 농도와 시간에 따라 처리하였다. 시료의 PGE
2와 첨가된 PGE
2-peroxidase conjugate와의 경쟁반응으 로 흡광도 변화와 농도와의 관계를 나타내는 표준 검량 선을 이용하여 각 세포배양액에 함유된 PGE
2의 함량을 계산하였다.
5. Statistics
본 실험의 결과는 여러 차례의 실험 중 평균적인 결과를 사용하였다. 통계적 검증은 one-way-ANOVA test를 사용하 고, 신뢰구간은 5%로 하여 p값이 0.005보다 낮은 경우를
*로, 표시하여 통계학적으로 유의하다고 해석하였다.
결 과
1. GT는 MDA-MB-231 유방암 세포의 세포사멸 및 염증반응을 유도한다
GT가 유방암세포의 세포사멸 및 염증반응에 미치는
효과를 알아보았다. 우선 GT를 농도 및 시간별로 처리
하여 세포사멸에 미치는 GT의 효과를 위상차 현미경과
MTT assay를 통해 확인해 보았다. 그 결과 GT는 농도 및
시간 의존적으로 세포사멸을 유도하는 것을 확인할 수
있었으며, 30μM 처리한 실험군에서는 약 50% 정도의
세포사멸이 유도되는 것을 확인할 수 있었다(Fig. 1A, B).
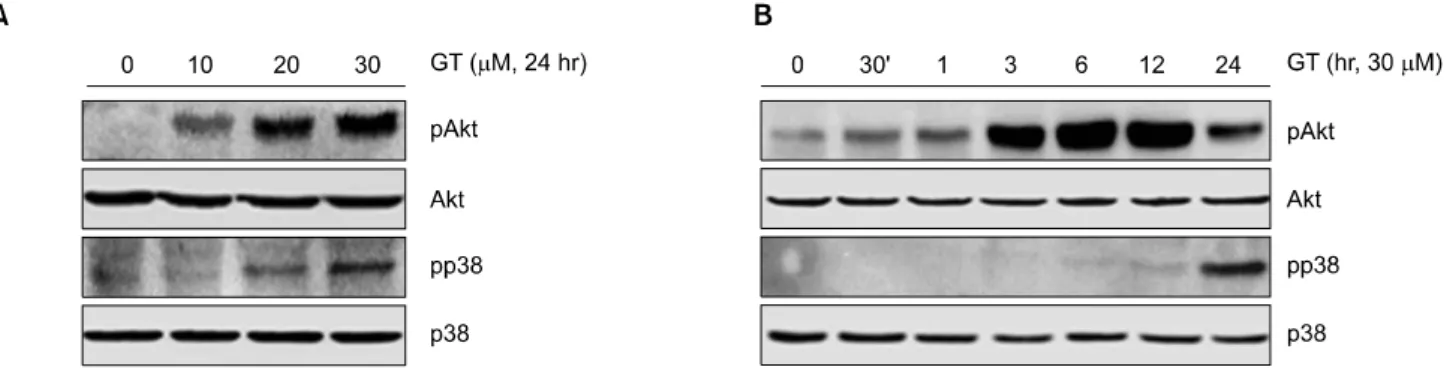
Fig. 3. GT leads to the activation of PI3kinase and p38 kinase in MDA-MB-231 cells. MDA-MB-231 cells were untreated (control) or treated with the indicated concentrations of GT for 24 hr. Expressions of pAkt, pp38 and actin were detected by Western blot analysis (A). MDA-MB-231 cells were untreated (control) or treated with the 30μM of GT for the indicated time periods. Expressions of pAkt, pp38 and actin were detected by Western blot analysis (B). The data represent the results of a typical experiment conducted at three times.
Fig. 2. GT causes the expression of COX-2 in MDA-MB-231 cells. MDA-MB-231 cells were untreated (control) or treated with the indicated concentrations of GT for 24 hr. Expressions of COX-2 and actin were detected by Western blot analysis (A).
MDA-MB-231 cells were untreated (control) or treated with the 30μM of GT for the indicated time periods. Expressions of COX-2 and actin were determined by Western blot analysis (B). MDA-MB-231 cells were untreated (control) or treated with the indicated concentrations of GT for 24 hr. PGE2 production was measured using a PGE2 assay kit (C). MDA-MB-231 cells were untreated (control) or treated with the 30μM of GT for the indicated time periods. PGE2 production was measured using a PGE2 assay kit (D). The data represent results of a typical experiment (A, B) or mean values and SD from five independent experiments (C, D). *p<0.05 compared with untreated cells.
또한 Western blot analysis를 통해 caspase-3의 활성을 확인 해 본 결과 농도 및 시간 의존적으로 active caspase-3의 발현량이 증가하는 것을 확인할 수 있었다(Fig. 1C).
다음으로는 GT를 농도 및 시간별로 처리하여 염증반
응에 미치는 GT의 효과를 확인해 보았다. 그 결과 GT는
농도 및 시간 의존적으로 염증반응 물질인 COX-2 및
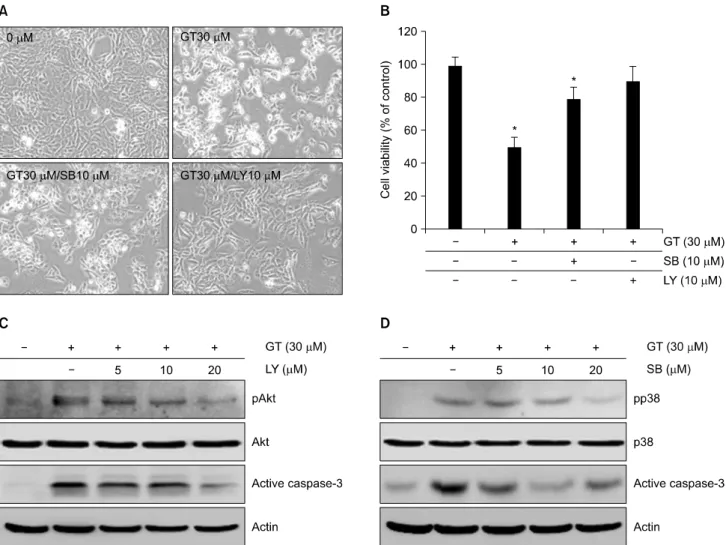
Fig. 4. PI3kinase and p38 kinase regulates GT-induced apoptosis in MDA-MB-231 cells. MDA-MB-231 cells were untreated (control) or treated with 30μM GT in the absence or presence of 10μM SB203580 or 10μM LY294002 for 24 hr. Cells photographs were taken using phase contrast microscope (mignification, ×100) (A). MDA-MB-231 cells were untreated (control) or treated with 30μM GT in the absence or presence of 10μM SB203580 or 10μM LY294002 for 24 hr. Apoptotic cells were determined by MTT assay (B). MDA-MB-231 cells were untreated (control) or treated with 30μM GT in the absence or presence of 10μM SB203580 or 10μM LY294002 for 24 hr. Expressions of pAkt, active caspase-3 and actin were detected by Western blot analysis (C). The data represent results of a typical experiment (C, D) or mean values±S.D. from at least four independent experiments (B). *p<0.05 compared with untreated cells.
PGE
2의 발현을 증가시키는 것을 Western blot analysis와 PGE
2assay를 통해 확인할 수 있었다(Fig. 2A, B). GT에 의한 COX-2 및 PGE
2의 발현은 12시간째 가장 많이 발현 하는 것을 확인할 수 있었으며, PGE
2의 발현이 최고조에 이르렀을 때는 대조군 세포에 비해 PGE
2의 발현량이 약 7∼8배 정도 증가하는 것을 확인할 수 있었다(Fig. 1B).
이와 같은 결과는 GT가 유방암 세포에서 세포 사멸 및 염증 반응을 유도한다는 것을 말해 주는 결과라 할 수 있다.
2. GT는 Akt와 p38의 활성을 증가시킨다
유방암세포에서 GT가 유도하는 세포사멸과 염증반응
을 조절하는 신호전달경로를 알아보았다. 기존에 암세
포의 세포사멸과 염증반응을 조절하는데 밀접하게 관련
되어 있다고 알려져 있는 신호전달 경로 중 PI3K와
MAPK의 활성을 확인해 보았다. 그 결과 GT처리에 의해
p38의 활성이 농도 및 시간 의존적으로 증가하는 것을
확인할 수 있었다(Fig. 3A, B). 또한 GT의 농도가 증가할
수록 Akt의 활성 역시 증가하는 것을 확인할 수 있었으
며, GT처리 후 12시간째 Akt의 활성이 가장 많이 증가하
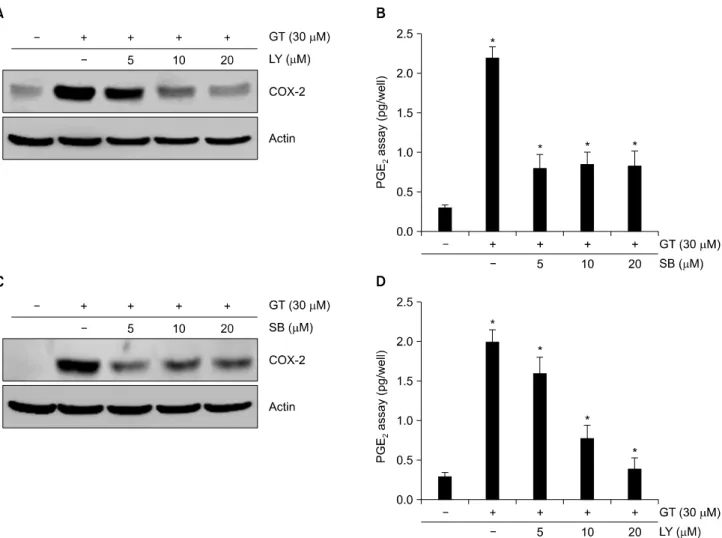
Fig. 5. PI3kinase and p38 kinase modulates GT-induced the expression of COX-2 in MDA-MB-231 cells. MDA-MB-231 cells were untreated (control) or treated with 30μM GT in the absence or presence of SB203580 or LY294002 for 24 hr. Expression of COX-2 was detected using Western blot analysis (A, C). PGE2 production was measured using a PGE2 assay kit (B, D). The data represent a typical result of experiment carried out more than five times with similar results (A, C) or mean values±S.D.
from at least five independent experiments (B, D). *p<0.05 compared with untreated cells.
였다. 그러나 MAPK 중 ERK의 활성에는 별다른 효과를 보이지 않았다(data not shown). 이와 같은 결과는 GT가 유 도하는 세포사멸과 염증반응이 Akt와 p38의 활성이 관 련되어 있음을 말해 주는 결과라 할 수 있다.
3. GT는 Akt와 p38 신호전달경로를 통한 caspase- 3 활성을 조절하여 세포 사멸을 조절한다
다음으로는 GT에 의해 유도되는 세포사멸이 Akt와 p38 신호전달경로에 의해 조절되는지 알아보기 위하여 Akt 상위신호전달경로인 PI3Kinase의 저해제인 LY294002 (LY)와 p38 kinase의 저해제인 SB203580 (SB)을 각각 1시 간 전 처리 후 GT를 처리하여 세포 생존율을 위상차 현 미경과 MTT assay를 수행하여 생존율을 확인해 보았다 (Fig. 4A, B). 그 결과 GT에 의해 증가하였던 세포사멸이
LY와 SB에 의해 억제되고, 세포 생존율이 회복되는 것을 확인 할 수 있었다(Fig. 4A). MTT assay 결과 역시 GT에 의한 세포 사멸률이 50% 정도였던 반면 LY와 SB 처리에 의해 각각 약 30%, 40% 정도 생존율이 회복되어 각각의 세포 사멸률이 10∼20%인 것을 확인할 수 있었다(Fig.
4B).
또한 세포사멸 관련 단백질인 caspase-3의 발현량을 확
인하기 위하여, LY와 SB를 농도별로 전 처리하여 active
caspase-3의 발현량을 확인 해 보았다(Fig. 4C, D). 그 결과
LY와 SB의 농도가 증가함에 따라 GT에 의해 증가하였던
active caspase-3의 발현량이 감소하였다. 이와 같은 결과
는 GT가 유도하는 세포사멸은 caspase-3 의존적인 신호
전달 경로를 통해 조절된다는 것을 말해 주는 결과라 할
수 있다.
4. GT는 Akt와 p38 신호전달경로를 통해 염증반응을 조절한다
다음으로는 LY와 SB를 처리하여 GT가 유도하는 염증 반응에 미치는 효과에 관한 연구를 수행하였다(Fig. 5).
보다 명확한 결과를 얻기 위하여 LY와 SB를 농도별로 1시간 전처리한 후 GT를 처리하였다. 그 결과, GT에 의 해 증가하였던 COX-2및 PGE
2의 발현이 LY와 SB의해 감 소되는 것을 확인할 수 있었다(Fig. 5). 특히 LY는 COX-2 및 PGE
2의 발현이 농도 의존적으로 감소시키는 것을 확 인할 수 있었다. 반면에 SB는 저 농도인 5μM처리만으 로도 GT가 증가시켰던 COX-2 및 PGE
2의 발현을 눈에 띄게 감소시키는 것을 확인할 수 있었으며 이러한 발현 량은 20μM까지 비슷한 수준으로 감소되는 것을 확인할 수 있었다(Fig. 5C, D). 이는 GT에 의한 염증반응이 Akt와 p38 신호전달경로를 통해 조절된다는 것을 말해주는 결 과이다.
결과를 종합해 보면, 유방암세포 MDA-MB-231에서 GT는 Akt와 p38 신호전달경로를 통해 세포 사멸 및 염 증반응을 조절한다는 것을 말해 준다. 염증반응과 세포 사멸과의 관련성에 있어서는 보다 많은 연구가 수행 되 어야 할 것으로 생각되며, 본 연구를 통한 유방암세포에 서 일어나는 세포사멸과 염증반응과 관련한 분자적 메 커니즘의 이해는 앞으로 유방암세포를 치료하는데 있어 서 귀중한 기초 자료로 제공될 수 있을 것이다.
고 찰
본 연구에서는 암세포의 증식, 생존, 혈관신생 및 전이 억제에 중요한 역할을 하는 것으로 알려져 있는 GT를 MDA-MB-231세포에 처리하여 암세포의 세포사멸 및 COX-2의 발현에 미치는 효과를 알아보았다.
10,25,26)COX-2 는 방광암, 위암, 결장암, 유방암, 전립선암, 간암을 포함 한 대부분의 암세포에서 정상세포보다 더욱 증가하여 발현되는 것을 확인할 수 있었다.
25∼28)COX-2의 산화력 은 발암전구물질을 발암물질로 바꾸는 능력을 가지고 있으며 종양의 형성을 개시하는 능력을 가지고 있다.
29,30)또한 COX-2는 여러 종류의 세포주의 종양성장과 전이 에 있어서 중요한 역할을 한다고 알려져 있으며, COX-2 를 knockout시킨 mouse에서 종양형성이 억제되는 것을 확인할 수 있었다.
31)따라서 celecoxib, aspirin, NSAIDS (Non-steroidal anti-inflammatory drugs)와 같은 다양한 COX-2 억제제를 세포 내에 처리하였을 때, 암세포의 성 장이 억제되는 것을 확인할 수 있었다.
32,33)COX-2를 세
포 내 과발현 시켰을 때 세포사멸이 감소된다는 연구결 과는 COX-2가 p53의 발현 및 전사 활성을 조절한다는 것을 증명해주는 결과라 할 수 있다.
34)또한, COX-2 활성 의 억제는 활성화된 p53의 발현을 유도한다는 결과 역시 발표된 바 있다. 반면에 다른 연구결과에서는 COX-2의 발현이 p53이 유도하는 세포사멸을 촉진시킨다는 연구 결과 역시 발표된 바 있다.
35)이와 같은 결과는 암세포에 서 COX-2의 발현과 세포사멸의 조절이 서로 밀접하게 연관되어 있음을 말해준다.
GT는 다양한 세포에서 p53의 발현을 촉진시킴으로써 세포 사멸을 유도하는 것으로 알려져 있다. Human mon- ocytic cell line U937에서 GT는 NFkB의 발현을 조절하여 p53의 발현을 증가시킴으로써 세포 성장을 억제하고, 세 포 사멸을 유도시키는 것을 확인할 수 있었다.
36)또한 A549세포에서caspase-3의 활성을 조절하여 세포사멸을 유도시키는 연구 결과 역시 확인할 수 있었다.
37)이와 같 은 이전 연구 결과는 GT가 다양한 암세포의 세포 사멸 을 조절한다는 것을 말해준다.
대부분의 암세포에는 종양 억제단백질로 알려져 있는 p53 단백질이 돌연변이 된 형태로 존재하고 있으며 유방 암 세포의 종류 중 약 25∼40%는 돌연변이 된 형태를 띄고 있다. MDA-MB-231 유방암세포에는 p53단백질이 발현되지 않기 때문에 세포사멸의 조절이 p53에 비의존 적인 caspases 활성을 통해 조절된다.
38)세포생존의 정상 적인 상태에서는 cytochrome c가 미토콘드리아의 막간공 간에 자리를 잡고 있으며, caspase-9/-3가 비활성 상태로 남아있다. 그러나 DNA가 손상이 되면 미토콘드리아의 손상이 일어나고, cytochrome c이 방출해서 Apaf-1과 결합 해 apoptosome을 형성한다. 이는 caspase-9/-3을 활성화 시 키며 세포사멸을 유도하게 된다.
39,40)GT는 다양한 농도로 24 시간 동안 MDA-MB-231세포
에 처리되었다. 그 결과 GT는 농도 의존적으로 세포 증
식을 감소시키고(Fig. 1A, B) caspase-3의 활성을 증가시킨
다는 것을 확인할 수 있었다(Fig. 1C). 이와 같은 결과는
GT가 caspase-3의 활성을 조절하여 세포의 증식을 억제
하고 세포사멸을 유도한다는 것을 말해 준다. 또한 GT
가 caspase의 발현을 통해 세포 사멸을 유도할 때, COX-2
의 발현이 조절되는지 확인하기 위하여 COX-2 발현 양
상의 변화를 Western blot analysis를 통해 확인하였다. 그
결과 caspase-3의 발현양상과 비슷하게, COX-2의 발현 양
상 역시 농도 의존적으로 증가되는 것을 확인 할 수 있
었다(Fig. 2A). 이와 같은 결과는 GT가 유도하는 세포 사
멸이 COX-2의 발현과 연관되어 있을 수 있음을 말해준
다.
Magnolol은 TPA가 유도하는 COX-2의 발현유도에 따 른 세포 증식 유도를 PI3kinase신호전달 경로를 통해 조 절한다는 연구가 보고된 바 있으며,
41)quercetin역시 rat liver epithelial cells에서 PI3kinase 신호전달 경로를 통해 COX-2의 발현을 조절한다는 연구 결과가 있다.
42)또한 UVB가 유도하는 mouse skin carcinogenesis의 COX-2 발현 에 있어서 p38 kinase의 활성이 증가하는 것을 확인 할 수 있었으며, 기존에 발표된 다양한 연구를 통해 PI3kin- ase와 p38 kinase는 COX-2의 발현 및 세포사멸을 조절하 는 데 중요한 신호 전달 경로라고 알려져 있다.
43)이 밖 에도 MDA-MB-231 세포에서 Serum withdrawal이 유도하 는 COX-2 mRNA의 안정화를 위해서 p38 kinase의 활성이 필요하다는 연구 결과 역시 보고된 바 있다.
44)따라서 GT에 의한 세포사멸과 COX-2의 발현이 PI3Kinase와 p38 kinase 신호 전달 경로를 통해 조절되는지 확인하기 위하 여 Akt와 p38의 활성 정도를 Western blot analysis를 통해 확인해 보았다. 그 결과, GT의 농도가 증가함에 따라 Akt와 p38의 활성 역시 증가되는 것을 확인할 수 있었으 며, LY294002와 SB203580을 통해 PI3Kinase와 Akt 신호 전달 경로를 차단하였을 때, GT에 의해 유도되었던 세 포사멸 및 COX-2의 발현이 감소되는 것을 확인 할 수 있었다. 이와 같은 결과는 PI3Kinase와 Akt 신호 전달 경 로를 통해 세포사멸 및 COX-2의 발현이 조절되어짐을 말해준다. 그러나 GT가 유도하는 세포 사멸에 있어서 COX-2의 발현이 관련되어 있는지에 관한 연구는 수행 되지 않았다. 따라서 앞으로 GT를 처리했을 때 조절되 는 세포사멸과 COX-2의 발현에 있어서 GT가 유도하는 COX-2의 발현이 세포 사멸을 조절하는지에 관한 연구 및 상호 관련성에 관하여는 더욱 연구되어야 할 것이다.
본 연구를 통해 규명된 세포 신호전달 경로는 앞으로 유 방암 치료에 있어서 중요한 자료로 사용될 수 있을 것으 로 사료된다.
결 론
Gallotannin (GT)은 식물에서 추출되는 탄닌의 일종으 로써 식물의 2차 대사산물로 식물의 방어기작에 사용되 는 폴리페놀성 중합체의 한 종류이다. GT는 의학적인 용도로써 항암, 항염증, 항바이러스, 항박테리아, 항기생 충 등의 기능으로 사용된다. 따라서 GT를 사람의 유방 암 세포인 MDA-MB-231에 처리하여 세포사멸과 염증반 응에 미치는 효과를 알아보았다. 본 연구의 결과 GT를 MDA-MB-231 세포에 처리하였을 경우, 세포 사멸을 유 도하고 세포 증식을 억제하는 것을 확인할 수 있었으며,
caspase-3의 활성 역시 증가 하는 것을 확인할 수 있었다.
또한 GT는 염증 관련 단백질인 cyclooxygenase-2 (COX-2) 및 prostaglandin E
2(PGE
2)의 발현을 증가시키는 것을 확 인 할 수 있었다. GT의 처리는 Akt와 p38의 활성을 증가 시키는 것을 확인 할 수 있었다. PI3-kinase의 억제제인 LY294002와 p38 kinase의 억제제인 SB203580를 전처리 하였을 때, GT에 의해 증가되었던 세포사멸과 COX-2의 발현이 감소하는 것을 확인할 수 있었다. 이와 같은 결과 를 통해 GT는 PI3-kinase와 p38 kinase 신호전달경로를 통 해 세포사멸 및 염증반응을 조절한다는 것을 확인 할 수 있었다.
감사의 글
이 논문은 교육과학기술부의 재원으로 한국연구재단 의 지원을 받아 수행된 기초연구 사업임(No. 2009- 0084569 & 2010-0003239).
참 고 문 헌
1) Ministry of Health and Welfare. Annual Report of the Korea Central Cancer Registry 2003.
2) Bang MH, Kim WK. Effect of curcumin on cancer invasion and matrix metalloproteinase-9 activity in MDA-MB-231 human breast cancer cell. Korean J Nutr 39, 756-761, 2006.
3) Smith WL. The eicosanoids and their biochemical mechanisms of action. Biochem J 259, 315-324, 1989.
4) Berenbaum F. Proinflammatory cytokines, prostaglandins, and the chondrocyte: mechanisms of intracellular activation. Joint Bone Spine 67, 561-564, 2000.
5) Funk CD. Prostaglandins and leukotrienes: advances in eicosa- noid biology. Science 294, 1871-1875, 2001.
6) Im JH, Kim SJ. Paclitaxel stimulates cyclooxygenase-2 expression via MAP kinase pathway in rabbit articular chondrocytes. J Exp Biomed Sci 15, 141-146, 2009.
7) Wulfing P, Diallo R, Muller C, Wulfing C, Poremba C, Heinecke A, Rody A, Greb RR, Bocker W, Kiesel L. Analysis of cyclooxygenase-2 expression in human breast cancer: high throughput tissue microarray analysis. J Cancer Res Clin Oncol 129, 375-382, 2003.
8) Howe LR, Subbaramaiah K, Brown AM, Dannenberg AJ.
Cyclooxygenase-2: a target for the prevention and treatment of breast cancer. Endocr Relat Cancer 8, 97-114, 2001.
9) Jeong SI, Zhou B, Bae JB, Kim NS, Kim SG, Kwon J, Kim DK, Shin TY, Jeon H, Lim JP, Kim H, Kim HK, Oh CH.
Apoptosis-inducing effect of akebia saponin D from the roots of Dipsacus asper Wall in U937 cells. Arch Pharm Res 31, 1399-1404, 2008.
10) Lu Y, Jiang F, Jiang H, Wu K, Zheng X, Cai Y, Katakowski
M, Chopp M, To SS. Gallic acid suppresses cell viability, proliferation, invasion and angiogenesis in human glioma cells.
Eur J Pharmacol 641, 102-107, 2010.
11) You BR, Park WH. Gallic acid-induced lung cancer cell death is related to glutathione depletion as well as reactive oxygen species increase. Toxicol In Vitro 24, 1356-1362, 2010.
12) Kassim M, Achoui M, Mustafa MR, Mohd MA, Yusoff KM.
Ellagic acid, phenolic acids, and flavonoids in Malaysian honey extracts demonstrate in vitro anti-inflammatory activity. Nutr Res 30, 650-659, 2010.
13) Bing SJ, Kim MJ, Park E, Ahn G, Kim DS, Ko RK, Lee NH, Shin T, Park JW, Jee Y. 1,2,3,4,6-penta-O-galloyl-beta- D-glucose protects splenocytes against radiation-induced apop- tosis in murine splenocytes. Biol Pharm Bull 33, 1122-1127, 2010.
14) Mahmood N, Piacente S, Pizza C, Burke A, Khan AI, Hay AJ. The anti-HIV activity and mechanisms of action of pure compounds isolated from Rosa damascena. Biochem Biophys Res Commun 229, 73-79, 1996.
15) Namba T, Kurokawa M, Kadota S, Shiraki K. Development of antiviral therapeutic agents from traditional medicines.
Yakugaku Zasshi 118, 383-400, 1998.
16) Liao SG, Zhang LJ, Sun F, Zhang JJ, Chen AY, Lan YY, Li YJ, Wang AM, He X, Xiong Y, Dong L, Chen XJ, Li YT, Zuo L, Wang YL. Antibacterial and anti-inflammatory effects of extracts and fractions from Polygonum capitatum.
J Ethnopharmacol 134, 1006-1009, 2011.
17) Kadowaki Y, Chari NS, Teo AE, Hashi A, Spurgers KB, McDonnell TJ. PI3 Kinase inhibition on TRAIL-induced apoptosis correlates with androgen-sensitivity and p21 expression in prostate cancer cells. Apoptosis 16, 627-635, 2011.
18) Xu R, Nakano K, Iwasaki H, Kumagai M, Wakabayashi R, Yamasaki A, Suzuki H, Mibu R, Onishi H, Katano M. Dual blockade of phosphatidylinositol 3'-kinase and mitogen- activated protein kinase pathways overcomes paclitaxel-resis- tance in colorectal cancer. Cancer Lett 306, 151-160, 2011.
19) Lee KM, Hwang MK, Lee DE, Lee KW, Lee HJ. Protective effect of quercetin against arsenite-induced COX-2 expression by targeting PI3K in rat liver epithelial cells. J Agric Food Chem 58, 5815-5820, 2010.
20) Kulik G, Weber MJ. Akt-dependent and -independent survival signaling pathways utilized by insulin-like growth factor I. Mol Cell Biol 18, 6711-6718, 1998.
21) Asano T, Yao Y, Zhu J, Li D, Abbruzzese JL, Reddy SA. The PI 3-kinase/Akt signaling pathway is activated due to aberrant Pten expression and targets transcription factors NF-kappaB and c-Myc in pancreatic cancer cells. Oncogene 23, 8571-8580, 2004.
22) Semba S, Moriya T, Kimura W, Yamakawa M. Phosphory- lated Akt/PKB controls cell growth and apoptosis in intra- ductal papillary-mucinous tumor and invasive ductal adeno- carcinoma of the pancreas. Pancreas 26, 250-257, 2003.
23) Davis RJ. Signal transduction by the JNK group of MAP kinases. Cell 103, 239-252, 2000.
24) Chang L, Karin M. Mammalian MAP kinase signalling cascades. Nature 410, 37-40, 2001.
25) Koga H, Sakisaka S, Ohishi M, Kawaguchi T, Taniguchi E, Sasatomi K, Harada M, Kusaba T, Tanaka M, Kimura R, Nakashima Y, Nakashima O, Kojiro M, Kurohiji T, Sata M.
Expression of cyclooxygenase-2 in human hepatocellular carcinoma: relevance to tumor dedifferentiation. Hepatology 29, 688-696, 1999.
26) Lim HY, Joo HJ, Choi JH, Yi JW, Yang MS, Cho DY, Kim HS, Nam DK, Lee KB, Kim HC. Increased expression of cyclooxygenase-2 protein in human gastric carcinoma. Clin Cancer Res 6, 519-525, 2000.
27) Soslow RA, Dannenberg AJ, Rush D, Woerner BM, Khan KN, Masferrer J, Koki AT. COX-2 is expressed in human pulmonary, colonic, and mammary tumors. Cancer 89, 2637- 2645, 2000.
28) Yoshimura R, Sano H, Mitsuhashi M, Kohno M, Chargui J, Wada S. Expression of cyclooxygenase-2 in patients with bladder carcinoma. J Urol 165, 1468-1472, 2001.
29) Dannenberg AJ, Zakim D. Chemoprevention of colorectal cancer through inhibition of cyclooxygenase-2. Semin Oncol 26, 499-504, 1999.
30) Eling TE, Thompson DC, Foureman GL, Curtis JF, Hughes MF. Prostaglandin H synthase and xenobiotic oxidation. Annu Rev Pharmacol Toxicol 30, 1-45, 1990.
31) Oshima M, Dinchuk JE, Kargman SL, Oshima H, Hancock B, Kwong E, Trzaskos JM, Evans JF, Taketo MM. Suppress- ion of intestinal polyposis in Apc delta716 knockout mice by inhibition of cyclooxygenase 2 (COX-2). Cell 87, 803-809, 1996.
32) Shiff SJ, Koutsos MI, Qiao L, Rigas B. Nonsteroidal antiin- flammatory drugs inhibit the proliferation of colon adeno- carcinoma cells: effects on cell cycle and apoptosis. Exp Cell Res 222, 179-188, 1996.
33) Turini ME, DuBois RN. Cyclooxygenase-2: a therapeutic target. Annu Rev Med 53, 35-57, 2002.
34) Corcoran CA, He Q, Huang Y, Sheikh MS. Cyclooxygenase-2 interacts with p53 and interferes with p53-dependent trans- cription and apoptosis. Oncogene 24, 1634-1640, 2005.
35) Tang HY, Shih A, Cao HJ, Davis FB, Davis PJ, Lin HY.
Resveratrol-induced cyclooxygenase-2 facilitates p53-depen- dent apoptosis in human breast cancer cells. Mol Cancer Ther 5, 2034-2042, 2006.
36) Kim NS, Jeong SI, Hwang BS, Lee YE, Kang SH, Lee HC, Oh CH. Gallic acid inhibits cell viability and induces apoptosis in human monocytic cell line U937. J Med Food 14, 240-246, 2011.
37) Maurya DK, Nandakumar N, Devasagayam TP. Anticancer property of gallic acid in A549, a human lung adenocarcinoma cell line, and possible mechanisms. J Clin Biochem Nutr 48, 85-90, 2011.
38) Chen JC, Chang NW, Chung JG, Chen KC. Saikosaponin-A induces apoptotic mechanism in human breast MDA-MB-231 and MCF-7 cancer cells. Am J Chin Med 31, 363-377, 2003.
39) Marshman E, Ottewell PD, Potten CS, Watson AJ. Caspase activation during spontaneous and radiation-induced apoptosis in the murine intestine. J Pathol 195, 285-292, 2001.
40) Shi Y. A structural view of mitochondria-mediated apoptosis.
Nat Struct Biol 8, 394-401, 2001.
41) Kuo DH, Lai YS, Lo CY, Cheng AC, Wu H, Pan MH.
Inhibitory effect of magnolol on TPA-induced skin inflam- mation and tumor promotion in mice. J Agric Food Chem 58, 5777-5783, 2010.
42) Lee KM, Hwang MK, Lee DE, Lee KW, Lee HJ. Protective effect of quercetin against arsenite-induced COX-2 expression
by targeting PI3K in rat liver epithelial cells. J Agric Food Chem 58, 5815-5820, 2010.
43) Dickinson S E, Olson ER, Zhang J, Cooper SJ, Melton T, Criswell PJ, Casanova A, Dong Z, Hu C, Saboda K, Jacobs ET, Alberts DS, Bowden GT. p38 MAP kinase plays a functional role in UVB-induced mouse skin carcinogenesis.
Mol Carcinog 50, 469-478, 2011.
44) Jang BC, Sanchez T, Schaefers HJ, Trifan OC, Liu CH, Creminon C, Huang CK, Hla T. Serum withdrawal-induced post-transcriptional stabilization of cyclooxygenase-2 mRNA in MDA-MB-231 mammary carcinoma cells requires the activity of the p38 stress-activated protein kinase. J Biol Chem 275, 39507-39515, 2000.