DOI 10.17480/psk.2016.60.5.243
B16 흑색종세포에서 디아족사이드에 의한 멜라닌 합성 촉진효과에 미치는 세포내 칼슘의 역할
이 용 수# 덕성여자대학교 약학대학
(Received July 13, 2016; Revised September 2, 2016; Accepted September 7, 2016)
Role of Intracellular Ca
2+in the Diazoxide-induced Stimulation of Melanin Synthesis in B16 Melanoma Cells
Yong Soo Lee#
College of Pharmacy, Duksung Women's University, Seoul 01369, Korea
Abstract — The effect of diazoxide on melanogenesis was investigated in B16 melanoma cells. Diazoxide stimulated the production of melanin in a dose-dependent manner. Diazoxide increased intracellular Ca2+ concentration, and BAPTA/AM significantly inhibited the diazoxide-induced melanogenesis. Diazoxide-induced intracellular Ca2+ increase and melano- genesis was significantly inhibited by EGTA. Taken together, these results suggest that intracellular Ca2+ influx may medi- ate the diazoxide-induced melanogenesis, and that diazoxide may be useful in the management of pigmentation disorder.
Keywords □ diazoxide, melanogenesis, Ca2+ signal, K+ channel, B16 melanoma cell
동물은 물론 사람의 피부색을 결정하는 색소인 멜라닌은 피부 기저층에 존재하는 멜라닌세포(melanocytes)에 의해 생성된 후 각질형성세포(keratinocytes)의 각화 과정에 의해 각질층으로 이 동된다.1)멜라닌의 합성은 멜라닌세포의 멜라노좀(melanosomes) 에 존재하는 여러 가지 효소에 의해 일어나며 그 중 티로시나제 (tyrosinase)가 율속 단계에 작용하는 효소로 알려져 있다.2)국소 적으로 너무 많이 생성되거나, 노화 등에 의해 피부의 기능이 감 소하면 멜라닌이 피부 표면에 침착되어 기미, 주근깨 등의 과색 소침착증(hyperpigmentation)을 일으킬 수 있다.3) 현재 국내·외 에서 티로시나제의 활성을 억제함으로써 멜라닌 합성을 저해하 는 효과를 지닌 물질을 식물로부터 추출하여 미백 효과를 나타 내는 기능성 화장품의 원료로 개발하려는 연구가 많이 이루어지 고 있다.4)
칼륨 통로는 진핵생물에는 어디에나 존재하며 안정막 전위, 활 동 전위 빈도 및 막전위의 재분극 속도의 조절 등과 같은 다 양한 세포 기능을 가지고 있다.5)또한 칼륨 통로는 세포의 분 열,6)분화7)및 세포 사멸8)의 조절에 깊이 관여하고 있음이 밝혀 졌다. 멜라닌세포에서도 다양한 칼륨 통로가 발현하고 있다는 사 실이 알려지고 있으나,9)멜라닌 합성과의 관련성은 우리가 조사 한 바로는 아직 보고된 바가 없다.
디아족사이드(Fig. 1)는 칼륨 통로의 한 종류인 ATP-민감성 칼륨 통로에 대한 활성제로 작용하며,10) 임상적으로는 급성 고혈압 또 는 악성 고혈압의 치료에서 혈관확장제로 사용되고 있다.11)디아 족사이드는 췌장에서 인슐린의 분비를 억제하는 기능이 있어 인슐 린종(insulinoma)이나 선천성 과인슐린증(congenital hyperinsulinism) 에서 나타나는 저혈당을 교정하는 목적으로도 사용된다.12)디아 족사이드는 AMPA 및 kainate 수용체의 양성 알로스텔릭 조절 인자(positive allosteric modulator)로 작용하여 인지 기능을 증 강시키는 효과를 나타내기도 한다.13) 또한, 간세포를 이용한 실 험에서는 디아족사이드가 세포내 칼슘 농도의 증가를 초래한다 는 사실도 보고된 바 있다.14)
#
Corresponding Author Yong Soo Lee
College of Pharmacy, Duksung Women’s University, Seoul 01369, Korea
Tel.: 02-901-8396 Fax.: 02-901-8386
E-mail: [email protected]
Short Report
종설따라서 본 연구에서는 디아족사이드가 멜라닌세포에서 멜라닌 합성에 미치는 영향을 조사하고, 그 기전을 칼륨 통로와 칼슘 신 호의 역할을 중심으로 규명하고자 하였다.
실험 방법
시약
Diazoxide, glibenclamide, tolbutamide, dantrolene, 3,4,5- trimethoxybenzoic acid-8-(diethylamino)-octyl ester(TMB-8), ethylene glycol-bis-(aminoethyl ether)N,N,N',N'-tetraacetic acid (EGTA), Dulbecco’s modified Eagle’s medium(DMEM), 3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide (MTT) 및 각종 용매와 염류는 Sigma–Aldrich(미국)에서 구입하 였다. Fetal bovine serum(FBS)와 penicillin-streptomycin 혼합 액은 GIBCO(미국)에서 구입하였다. 1-(2,5-carboxyoxazol-2-yl- 6-aminobenzfuran-5-oxyl)-2-(2’-amino-methylphenoxy)-ethane- N,N,N’N’-tetraacetoxylmethyl ester(Fura-2/AM), bis-(o- aminophenoxy)-ethane-N,N,N',N'-tetraacetic acid/acetoxymethyl ester(BAPTA/AM) 및 potassium-binding benzofuran isophthalate/
aceoxylmethyl ester(PBFI/AM)은 Molecular Probes(미국)에서 구입하였다. 형광 탐침은 dimethyl sulfoxide(DMSO)에 녹여 농 축액으로 만든 후 물로 희석하여 사용하였다.
세포 배양
B16 흑색종 세포는 서울대학교 한국세포주은행에서 구입하였 다. 구입한 세포는 10% FBS와 penicillin 100 IU/mL와 streptomycin 50μg/mL을 함유한 DMEM 용액으로 37oC로 유지되는 5% CO2 배양기(Forma, 미국)에서 배양하였다.
세포 생존율 측정
살아있는 세포는 미토콘드리아에 존재하는 탈수소 효소에 의 해 MTT가 MTT-formazan으로 전환되므로 이것의 양을 재면 살 아있는 세포의 수를 측정할 수 있다. 세포는 1 mL 배지에 디아 족사이드를 최종 농도가 1∼200 μM이 되도록 처리하여 24 well
plate에서 48시간 동안 배양하였다. 배양이 끝난 세포에 MTT 용 액(2.5 mg/mL H2O) 100μL를 첨가하여 37oC에서 4시간 동안 반응시켰다. 생성된 MTT-formazan 결정체를 분리하기 위하여 세포 현탁액을 eppendorf tube에 옮겨 원심분리하였다(1500 rpm, 4분). 상등액을 조심스럽게 제거하고 DMSO 100 μL를 첨가하여 결정체를 용해시킨 후 540 nm에서 ELISA reader (Molecular Device, 미국)로 흡광도를 측정하였다. 세포 생존율은 대조군과 비교하여 흡광도의 백분율로 나타내었다.
멜라닌 정량
B16 세포에서 멜라닌을 측정할 때에는 phenol red가 없는 DMEM을 사용하여 24 well plate에 mL 당 5×104개로 분주하 고 12시간이 지난 뒤 세포가 plate에 완전히 부착된 것을 확인한 후 디아족사이드 등 약물을 처리하고 48시간 지난 뒤에 세포를 수집하여 세포 수를 측정하고, 1200 rpm에서 5분간 원심 분리하 여 침전한 후, 1 mL 균질완충액(50 mM sodium phosphate pH 6.5, 1% Triton X-100, 2 mM PMSF)로 용해시켰다. 여기서 얻은 세포 pellet을 1 N NaOH, 10% DMSO 용액 200 μL를 첨가하고 vortex 후 멜라닌을 완전히 녹인 다음 96 well plate에 옮긴 후 ELISA reader(Molecular Device, 미국)를 사용하여 405 nm에서 흡광도를 측정하였다. 멜라닌 표준품을 이용한 표준 검량선을 이 용하여 멜라닌 양을 산출하였다. 멜라닌 생성량은 단위세포(104 개)에서의 멜라닌 생성량으로 비교하였다.
세포내 칼륨 이온 농도 측정
세포내 칼륨 이온 농도는 형광 탐침인 PBFI/AM을 이용하여 측정하였다. 비이온성 계면 활성제인 0.02% pluronic F-127을 포 함한 Hank 용액의 세포 현탁액에 5 μM의 PBFI/AM을 가해 37oC 에서 2시간 동안 진탕 배양하여 세포내로 봉입시킨 후, 봉입되지 않은 PBFI/AM은 원심분리하여 제거시켰다. 다시 세포를 Krebs- Ringer 완충액(125 mM NaCl, 5 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 5 mM NaHCO3, 25 mM HEPES, 6 mM glucose, pH 7.4)에 현탁시킨 후 cuvette에 옮겨 340 nm 및 380 nm 파장에서 excitation 시켜, 500 nm 파장에서 나오는 형광을 형광 분석기로 측정하였다. 이 때 cuvette 내의 세포 현탁액을 계속 교 반하여 세포가 가라앉는 것을 방지하였다. 세포내 칼륨 농도는 두 파장에서 나온 형광 세기의 비로써 나타내었다. 약물 투여는 원하는 최종 농도가 되게 원액(stock solution)을 사용하여 10 μL 이내로 정밀 주사기를 이용하여 cuvette 내로 주입하였다.
세포내 칼슘 이온 농도 측정
B16 세포를 5 μM Fura-2/AM을 가해 37oC에서 30분간 배양 하였고 비봉입 Fura-2/AM은 150 g에서 30분간 원심분리하여 제 거하였다. 세포를 Krebs-Ringer 완충액(125 mM NaCl, 5 mM
Fig. 1 − Chemical structure of diazoxide.
KCl, 1.3 mM CaCl2, 1.2 mM KH2PO4, 1.2 mM MgSO4, 5 mM NaHCO3, 25 mM HEPES, 6 mM glucose 및 2.5 mM probenecid, pH 7.4)에 세포 밀도가 mL당 2×106개가 되도록 현탁시켰다.
Fura-2가 봉입된 세포를 형광 측정 전까지 25oC에서 90분간 유 지시킨 후 교반봉이 들어있는 quartz cuvette에 0.5 mL 세포 현 탁액을 주입하여 Hitachi F-4500 형광 분석기를 사용하여 여기 파장을 340 및 380 nm에서 반복하게 하고 형광 발광을 510 nm 에서 측정하였다. 세포내 칼슘 농도는 340:380 nm 형광비로 나타 내었다. 약물 투여는 원하는 최종 농도가 되게 원액을 사용하여 10μL 이내로 정밀 주사기를 이용하여 cuvette 내로 주입하였다.
자료 분석 및 통계적 검정
모든 실험은 네 번 반복해서 실시하고 실험 결과는 대조군의 조건에 대한 백분율로 나타내었다. 실험 결과는 평균값±SEM으 로 표시하고 ANOVA로 분석하며 각각의 유의성 비교는 Student- Newman-Keul’s test를 이용하여 실시하였다. P값이 0.05 이하 인 경우에만 통계학적 유의성이 있는 것으로 간주하였다.
실험 결과 및 고찰
디아족사이드의 세포 독성
디아족사이드가 세포 독성에 미치는 농도를 조사하여 향후 실 험에 사용될 농도 범위를 결정하기 위해서 MTT 분석을 시행하 였다. B16 흑색종 세포에 대한 디아족사이드의 세포 독성을 측 정한 결과, 70 μM 이하의 농도로 처리했을 때 세포 생존율은 유 의성 있는 변화를 보이지 않았으며, 100 μM 이상을 처리했을 때 생존율이 유의성 있게 저하되었다(Fig. 2). 따라서, 향 후 실험에 서 디아족사이드는 50 μM 이하의 농도로 사용하였다.
여러 종류의 세포에서 디아족사이드는 본 실험에서 사용한 비 슷한 농도의 범위에서는 독성을 나타내지 않았으며, 오히려 본 연구실에서 시행한 조골세포 실험에서 과산화수소에 의한 세포 독성을 억제하는 결과를 보였고15)또한, 신경세포에서도 베타-아 밀로이드(1-42)에 의한 세포 독성을 억제하는 효과를 나타내었다.16)
디아족사이드의 멜라닌 생성 촉진 효과
본 연구에서는 멜라닌 합성에 미치는 칼륨 통로의 역할을 알 아보고자 하였다. 이를 위하여 칼륨 통로에 직접 작용하여 통로 를 열어 주는 활성이 있다고 알려진 디아족사이드10)를 이용하여 이 약물이 멜라닌 합성에 미치는 영향을 확인하였다. 디아족사 이드를 처리하고 48시간 동안 배양 후 그 효과를 관찰하였는데 그 이유는 이 시간대에서 대조군, 즉 약물을 가하지 않은 상태에 서 분석이 가능한 멜라닌 합성이 이루어지기 때문이다.17)배양 후 B16 세포를 수집하여 멜라닌 양을 측정한 결과, 디아족사이 드를 처리한 군에서 농도 의존적으로 멜라닌 합성이 증가되었으
며 20 μM 및 50 μM 농도 처리군에서 대조군에 비해 각각 약 2.2 및 2.7배가 증가하여 통계적으로 유의한 촉진 효과를 나타내 었다(Fig. 3). 우리가 조사한 바로는 멜라닌 합성에 미치는 칼륨 이온 자체 혹은 칼륨 통로 조절 약물의 영향에 대한 연구 결과 는 아직까지 보고된 바가 없다.
본 연구실에서는 B16 세포에서 식물성 플라보노이드 성분인 아피제닌에 의한 멜라닌 합성 촉진 작용이 칼륨-염소이온수송체 의 활성화를 통해서 일어난다는 사실을 확인한 바 있다.18) 아피 제닌에 의한 칼륨-염소이온수송체의 활성화는 세포막을 통하여 칼륨과 염소 이온을 동시에 이동시키지만 이 중 염소 이온의 변 화가 멜라닌 합성과 연관되어 있는 것을 밝힌 바 있다.18)
디아족사이드에 의한 멜라닌 생성 촉진 작용 기전
디아족사이드는 ATP-민감성 칼륨 통로를 직접적으로 여는 약 물10)로 알려져 있기 때문에 먼저 세포내 칼륨 농도에 미치는 디 아족사이드의 영향을 조사하였다. B16 세포에서 디아족사이드 (50μM)를 처리했을 때 세포내 칼륨 농도의 변화는 관찰할 수 없 었다(Fig. 4A). 또한, ATP-민감성 칼륨 통로의 억제제로 알려진 glibenclamide(100μM) 및 tolbutamide(100 μM)를 전처리했을 때 디아족사이드에 의한 멜라닌 합성 촉진 효과에 유의성 있는 영향 을 미치지 않았다(Fig. 4B). 이 결과는 디아족사이드의 ATP-민감 성 칼륨 통로 활성화와 멜라닌 합성 촉진 효과와는 무관함을 시 사한다. 본 실험에 사용한 B16 세포에서 칼륨 통로의 한 아형인
Fig. 2 − Effects of diazoxide on cell viability in B16 melanoma cells.
Cells were incubated with or without each concentration of
diazoxide for 48 hr. Cell viability assay was done by the MTT
staining method. Results are expressed as a percentage of
control condition in which cells were grown in medium
without drugs. Data points represent the mean values of
four replications with bars indicating SEM.
*P<0.05
compared to control in which the cells were incubated with
diazoxide-free medium.
칼슘-불활성 칼륨 통로(Ca2+-inactivated K+ channel)의 존재가 확 인 바19)있지만 아직까지 ATP-민감성 칼륨 통로의 존재에 대해 선 보고된 바 없다. 따라서, B16 세포에서 디아족사이드의 작용 은 ATP-민감성 칼륨 통로의 활성화와는 무관한 것 같다.
본 연구실에서는 멜라닌 합성 기전에 칼슘 신호가 중요한 역 할을 담당하고 있다는 사실을 밝힌 바20)있고 또한, 간세포를 이 용한 실험에서는 디아족사이드가 세포내 칼슘 농도의 증가를 초 래한다는 사실을 보고한 바14)있으므로 디아족사이드에 의한 멜 라닌 합성 촉진 작용에 미치는 칼슘 신호의 역할을 조사하였다.
Fura-2 형광법을 통한 세포내 칼슘 측정에서 디아족사이드 (50μM)는 빠르게 세포내 칼슘 농도를 증가시켰으며(Fig. 5A), 이 작용은 농도 의존적으로 나타났다(Fig. 5B). 또한, 세포내 칼 슘 킬레이트인 BAPTA/AM(1 μM)을 전처리한 결과 디아족사이 드에 의한 멜라닌 합성 촉진 작용을 유의성 있게 억제하였다(Fig.
5C). 이 결과로 미루어 보아 디아족사이드에 의해 일어나는 멜 라닌 합성 촉진 작용은 세포내 칼슘 증가에 의해 매개되는 것 같다.
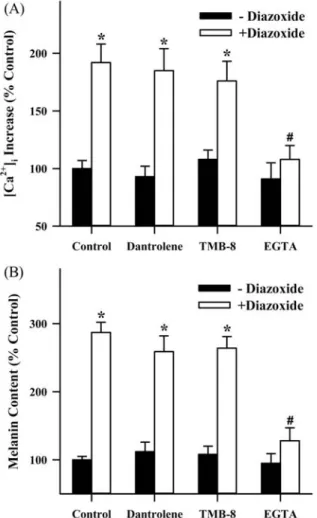
다음은 디아족사이드에 의한 세포내 칼슘 증가가 어떤 경로를 통해 일어나는 지를 추적하기 위해 먼저 EGTA를 첨가하여 세 포내 칼슘 변화를 관찰하였다. EGTA는 칼슘 이온을 붙여 착화합 물을 형성하므로 세포외 칼슘 농도를 효과적으로 감소할 수 있기
때문에 EGTA의 첨가로 칼슘 유입 과정을 차단할 수가 있다. 실 험결과를 보면 EGTA의 첨가는 디아족사이드(50 μM)에 의한 세 포내 칼슘 증가(Fig. 6A)와 멜라닌 합성 촉진 작용(Fig. 6B)을 유 의성 있게 차단하였다. 하지만, 칼슘 통로로 작용하여 소포체막 으로부터 칼슘을 유리시키는 ryanodine 수용체와 IP3수용체에 대하여 각각 길항 효과를 나타내는 dantrolene(50 μM) 및 TMB- 8(20μM)을 처리한 결과는 디아족사이드에 의한 칼슘 증가와(Fig.
Fig. 3 − Effect of diazoxide on the production of melanin in B16 melanoma cells. Melanin content was measured by the method described in the Method section. Briefly, the cells were initially grown for 12 hr in order to attach them to the bottom of the culture flasks. Then, the cells were incubated with each concentration of diazoxide for 48 hr. Each column represents the mean value of four replications with bars indicating SEM.
*P<0.05 compared to control in which the cells were incubated with diazoxide-free medium.
Fig. 4 − No involvement of ATP-sensitive K
+channels in the
diazoxide-induced production of melanin in B16 melanoma
cells. Intracellular K
+concentration was assessed by PBFI
fluorescence probe, and the data (A) represent intracellular
K
+changes with time. The arrow shows the time point for
addition of diazoxide (50 µM). The data (B) represent the
effect of blockers of ATP-sensitive K
+channels on the
production of melanin induced by diazoxide. Glibenclamide
(100 µM) and tolbutamide (100 µM) were added 30 min
before treatment with diazoxide (50 µM). Each column
represents the mean value of four replications with bars
indicating SEM.
*P<0.05 compared to negative control.
6A) 멜라닌 합성 촉진 작용(Fig. 6B)에 유의성 있는 영향을 미치 지 않는 것으로 나타났다. 이 결과는 세포 내에서 칼슘 유리가 아닌 세포 바깥으로부터 칼슘 유입이 디아족사이드에 의한 멜라 닌 합성 촉진 작용을 매개하는 중요한 신호라는 사실을 입증한다.
세포내 칼슘 신호는 세포의 분열, 분화 및 사멸은 물론 근육 수축과 호르몬이나 신경전달물질의 외포 작용 등 세포 기능에 중
Fig. 5 − Role of intracellular Ca
2+in the diazoxide-induced production
of melanin in B16 melanoma cells. Intracellular Ca
2+concentration was assessed by Fura-2 fluorescence technique, and the data (A) represent intracellular Ca
2+changes with time. The arrow shows the time point for addition of diazoxide (50
μM). In the data (B) intracellular Ca
2+increases are expressed as a percentage of control condition in which the cells were incubated with diazoxide-free medium. Each column represents the mean value of four replications with bars indicating SEM.
*P<0.05 compared to control. The data (C) represent the effect of BAPTA/AM, an intracellular Ca
2+chelator, on the production of melanin induced by diazoxide.
BAPTA/AM (1
μM) was added 30 min before treatment with diazoxide (50
μM). Each column represents the mean value of four replications with bars indicating SEM.
*P<0.05 compared to negative control.
#P<0.05 compared to diazoxide alone.
Fig. 6 − Role of Ca
2+influx in the diazoxide-induced intracellular Ca
2+increase and melanin synthesis in B16 melanoma cells.
Intracellular Ca
2+concentration (A) and melanin content (B) were assessed by the techniques, as explained in Fig. 4 and 2, respectively. In the experiments of (A) a nominal Ca
2+-free medium containing 100 µM EGTA was used. Dantrolene (50 µM) and TMB-8 (20 µM) were added 5 min before diazoxide application. In the data the diazoxide (50 µM)- induced changes in intracellular Ca
2+levels are expressed as a percentage of control condition in which the cells were incubated with diazoxide-free medium. In the experiments of (B), dantrolene (50 µM), TMB-8 (20 µM) and EGTA (1 mM) were added 30 min before diazoxide (50 µM) treatment. The data (A and B) represent the mean values of four replications with bars indicating SEM. *p<0.05 compared to control.
#
p<0.05 compared to diazoxide alone.
요한 역할을 담당하고 있다고 잘 알려져 있으며, 최근 연구에서 는 멜라닌 합성 기전에도 관여하고 있음이 밝혀지고 있다.
Carsberg 등의 연구에서 보면 자외선의 조사에 의해 세포내 칼 슘 농도 및 멜라닌 합성이 증가되고 인위적으로 세포외 칼슘 농 도를 높였을 때도 멜라닌 합성이 증가되었다.21)또한, 스타틴 계 열 약물인 로바스타틴에 의한 멜라닌 합성 촉진 작용에 세포내 칼슘 증가가 중요한 역할을 한다는 사실도 밝혀졌다.20)이와는 반대로 칼슘 통로 억제제인 verapamil이 멜라닌 합성을 촉진시 킨다는 결과도 발표되었다.22) 이러한 상반된 결과가 실험에 사용 한 세포 종류에 기인한 것인 지 또는 다른 이유에 의한 것인 지 는 현재로선 정확히 알 수 없으며 추후 연구에서 밝혀할 과제이다.
칼슘 이온의 증가가 어떻게 멜라닌 합성을 촉진시키는 지에 대해서는 아직 확실히 이해되고 있지 않다. 그럼에도 불구하고, 그 이유를 밝혀진 사실에 근거하여 추측해 본다면 다음과 같다.
하나의 가능성은 칼슘 이온의 증가가 멜라닌세포에 존재한다고 알려진 protein kinase C-β23)를 활성화시켜 일어날 수 있다는 것 이다. 왜냐하면, 이 효소는 멜라닌 합성에 중요한 티로시나제를 인산화하여 효소 활성을 증가시킬 수 있으므로,24)멜라닌 합성 을 유도할 수 있을 것이다.
다른 가능성은 칼슘 이온이 멜라노좀에 작용하여 멜라닌 합성 을 촉진할 수 있다는 것이다. 멜라노좀 내부에는 실제로 칼슘 이 온이 많이 존재하고 있는데,25) 이것은 멜라닌 자체가 칼슘 이온 에 잘 결합하는 성질 때문이다.26)이 경우 세포막을 통해 칼슘 이온이 유입되면 멜라노좀막에 존재하는 칼슘 펌프를 통해 멜라 노좀 내로 수송되는데 이 때 멜라노좀 막전압과 내강 pH의 변 화를 유발하게 되고,27)이것이 멜라닌 합성에 관여하는 여러 효 소의 활성을 조절한다는 가설이다.28)종합적으로, 세포내 칼슘 신호가 멜라닌 합성 과정에 모종의 역할을 할 것으로는 생각되 지만 좀 더 확실한 결론을 도출하기 위해서는 이를 입증하는 많 은 연구가 필요할 것으로 사료된다.
결 론
본 연구에서는 디아족사이드가 멜라닌 합성에 미치는 영향을 알아보고 그 기전을 규명하고자 하였다. 실험 결과에서 디아족 사이드는 B16 세포에서 농도 의존적으로 멜라닌 합성을 증가시 켰다. 디아족사이드는 세포내 칼슘 농도를 용량 의존적으로 증 가시켰으며 세포내 칼슘 킬레이트인 BAPTA/AM의 전처리에 의 해 디아족사이드에 의한 세포내 칼슘 증가와 멜라닌 합성 촉진 작용이 유의성 있게 감소되었다. 디아족사이드에 의한 세포내 칼 슘 증가와 멜라닌 합성 촉진 작용은 세포외 칼슘 킬레이트인 EGTA에 의해 차단되었다. 하지만, 소포체로부터 칼슘 유리를 억 제하는 약물인 dantrolene과 TMB-8에 의해서는 변화가 없었다.
이 결과는 디아족사이드에 의한 멜라닌 합성 촉진 작용이 세포
내 칼슘 유입에 의해 매개된다는 사실을 시사한다. 또한, 이 연 구의 결과로써 디아족사이드는 멜라닌 합성 이상에 의한 피부 색 소 질환의 치료제 개발 연구에 활용될 가치가 있다고 판단된다.
감사의 말씀
본 연구는 덕성여자대학교 2015년도 교내연구비 지원에 의해 수행되었음.
References
1) Simon, J. D., Peles, D., Wakamatsu, K. and Ito, S. : Current challenges in understanding melanogenesis: bridging chemistry, biological control, morphology, and function. Pigment Cell Melanoma Res. 22, 563 (2009).
2) Casanola-Martin, G. M., Le-Thi-Thu, H., Marrero-Ponce, Y., Castillo-Garit, J. A., Torrens, F., Rescigno, A., Abad, C. and Khan, M. T. : Tyrosinase enzyme: an overview on a pharmacological target. Curr. Top. Med. Chem. 14, 1494 (2014).
3) Speeckaert, R., Van Gele, M., Speeckaert, M. M., Lambert, J. and van Geel, N. : The biology of hyperpigmentation syndromes. Pigment Cell Melanoma Res. 27, 512 (2014).
4) Alexis, A. F. : New and emerging treatments for hyperpigmentation. J. Drugs Dermatol. 13, 382 (2014).
5) Buckingham, S. D., Kidd, J. F., Law, R. J., Franks, C. J. and Sattelle, D. B. : Structure and function of two-pore-domain K+ channels: contributions from genetic model organisms.
Trends Pharmacol. Sci. 26, 361 (2005).
6) Urrego, D., Tomczak, A. P., Zahed, F., Stuhmer, W., Pardo, L. A. : Potassium channels in cell cycle and cell prolifera- tion. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 369, 20130094 (2014).
7) Song, M. S., Choi, S. Y., Ryu, P. D. and Lee, S. Y. : Voltage- gated K+ channel, Kv3.3 is involved in hemin-induced K562 differentiation. PLoS One 11, e0148633 (2016).
8) Demidchik, V., Straltsova, D., Medvedev, S. S., Pozhvanov, G. A., Sokolik, A. and Yurin, V. : Stress-induced electrolyte leakage: the role of K+-permeable channels and involvement in programmed cell death and metabolic adjustment. J. Exp.
Bot. 65, 1259 (2014).
9) Allen, D. H., Lepple-Wienhues, A. and Cahalan, M. D. : Ion channel phenotype of melanoma cell lines. J. Membr. Biol.
155, 27 (1997).
10) Challinor-Rogers, J. L. and McPherson, G. A. : Potassium channel openers and other regulators of KATP channels.
Clin. Exp. Pharmacol. Physiol. 21, 583 (1994).
11) van Hamersvelt, H. W., Kloke, H. J., de Jong, D. J.,
Koene, R. A. and Huysmans, F. T. : Oedema formation with the vasodilators nifedipine and diazoxide: direct local effect or sodium retention? J. Hypertens. 14, 1041 (1996).
12) Huang, Q., Bu, S., Yu, Y., Guo, Z., Ghatnekar, G., Bu, M., Yang, L., Lu, B., Feng, Z., Liu, S. and Wang, F. : Diazoxide prevents diabetes through inhibiting pancreatic β-cells from apoptosis via Bcl-2/Bax rate and p38-β mitogen-activated protein kinase. Endocrinology 148, 81 (2007).
13) Randle, J. C., Biton, C. and Lepagnol, J. M. : Allosteric potentiation by diazoxide of AMPA receptor currents and synaptic potentials. Eur. J. Pharmacol. 247, 257 (1993).
14) Lee, Y. S. : Mechanism of apoptosis induced by diazoxide, a K+ channel opener, in HepG2 human hepatoma cells. Arch.
Pharm. Res. 27, 305 (2004).
15) Choi, E. M., Kim, G. H. and Lee, Y. S. : Diazoxide protects against hydrogen peroxide-induced toxicity in the osteoblastic MC3T3-E1 cells. Eur. J. Pharmacol. 624, 45 (2009).
16) Zeng, X., Wang, T., Jiang, L., Ma, G., Tan, S., Li, J., Gao, J., Liu, K. and Zhang, Y. : Diazoxide and cyclosporin A protect primary cholinergic neurons against β-amyloid (1-42)- induced cytotoxicity. Neurol. Res. 35, 529 (2013).
17) Lee, Y. S. : Role of Ca2+-activated Cl- channels in the stimulation of melanin synthesis induced by cyclosporin A in B16 melanoma cells. Yakhak Hoeji 59, 177 (2015).
18) Lee, Y. S. : Role of K+-Cl--cotransporter in the apigenin- induced stimulation of melanogenesis in B16 melanoma cells. Yakhak Hoeji 52, 500 (2008).
19) Ma, L. J., Liu, L. Y., Jiao, S., Wei, S. M. and Mei, Y. A. : Ca2+-inactivated K+ current is modulated by endothelin-1 in B-16 murine melanoma cells. Pigment Cell Res. 16, 463 (2003).
20) Lee, Y. S. : Role of intracellular Ca2+ in the lovastatin-
induced stimulation of melanin synthesis in B16 melanoma cells. Yakhak Hoeji 57, 24 (2013).
21) Carsberg, C. J., Jones, K. T., Sharpe, G. R. and Friedmann, P. S. : Intracellular calcium modulates the responses of human melanocytes to melanogenic stimuli. J. Dermatol. Sci.
9, 157 (1995).
22) Parker, C. and Sherbet, G. V. : The Ca2+ channel blocker verapamil enhances melanogenesis without altering metastatic potential in the B16 murine melanoma. Melanoma Res. 3, 347 (1993).
23) Park, H. Y., Wu, C., Yonemoto, L., Murphy-Smith, M., Wu, H., Stachur, C. M. and Gilchrest, B. A. : MITF mediates cAMP-induced protein kinase C-β expression in human melanocytes. Biochem. J. 395, 571 (2006).
24) Park, H. Y., Perez, J. M., Laursen, R., Hara, M. and Gilchrest, B. A. : Protein kinase C-β activates tyrosinase by phosphorylating serine residues in its cytoplasmic domain. J.
Biol. Chem. 274, 16470 (1999).
25) Ulshafer, R. J., Allen, C. B. and Rubin, M. L. : Distributions of elements in the human retinal-pigment epithelium. Arch.
Ophthalmol. 108, 113 (1990).
26) Bush, W. D. and Simon, J. D. : Quantification of Ca2+
binding to melanin supports the hypothesis that melanosomes serve a functional role in regulating calcium homeostasis.
Pigment Cell Res. 20, 134 (2007).
27) Bellono, N. W., Kammel, L. G., Zimmerman, A. L. and Oancea, E. : UV light phototransduction activates transient receptor potential A1 ion channels in human melanocytes.
Proc. Natl. Acad. Sci. U. S. A. 110, 2383 (2013).
28) Bellono, N. W. and Oancea, E. V. : Ion transport in pigmentation. Arch. Biochem. Biophys. 563, 35 (2014).