B16 흑색종세포에서 아피제닌에 의한 멜라닌 합성 촉진효과에 미치는 칼륨-염소이온수송체의 역할
이 용 수# 덕성여자대학교 약학대학
(Received November 5, 2008; Revised November 21, 2008)
Role of K
+-Cl
--cotransporter in the Apigenin-induced Stimulation of Melanogenesis in B16 Melanoma Cells
Yong Soo Lee#
College of Pharmacy, Duksung Women’s University, Seoul 132714, Korea
Abstract
— Apigenin, a natural flavonoid found in a variety of vegetables and fruits, has been shown to possess many bio- logical functions. In this study we found that apigenin stimulated melanin synthesis in a dose-dependent manner in B16 murine melanoma cells. Since in our previous study K
+-Cl
--cotransport (KCC) has been shown to mediate the mechanism of action of apigenin in neuronal cells, we further investigated the role of KCC in the melanogenesis-stimulating effect of apigenin in B16 cells. At nontoxic concentrations apigenin induced Cl
--dependent K
+efflux, a hallmark of KCC activity, which was markedly prevented by a specific KCC inhibitor R-(+)-[(2-n-butyl-6,7-dichloro-2-cyclopentyl-2,3-dihydro-1-oxo- 1H-inden-5-yl)oxy]acetic acid (DIOA). These results indicate that KCC is functionally present, and activated by apigenin in the B16 cells. In addition, the apigenin-induced stimulation of melanogenesis was also significantly inhibited by DIOA. N- Ethylmaleimide (NEM), a known KCC activator, induced Cl
-efflux and stimulated melanogenesis in a concentration-depen- dent fashion. Both effects of NEM were significantly inhibited by DIOA. Taken together, these results suggest that apigenin can modulate melanogenesis through the activation of a membrane ion transporter, KCC in B16 cells. These results further suggest that apigenin may be a good candidate in the therapeutic strategy for hypopigmentation disorders, such as vitiligo.
Keywords □
K
+-Cl
--cotransport, apigenin, B16 melanoma cell, melanogenesis, vitiligo
아피제닌은자연에존재하는플라보노이드류로서파슬리
,
박 하,
레몬,
들깨,
딸기류및여러과일에다량함유되어있다.
1)아피제닌은본연구실및다른연구실의실험결과로서여러조직 에서항염
,
2,3)항발암4)및항암작용5-7)을가진다는사실이밝혀 졌다.
지금까지의연구에서보면식물성플라보노이드류는세포실험에서활성산소종의발생을억제하는항산화효과를나타낸다 는사실이알려지고있다
.
8-11)하지만,
최근아피제닌은전립선암 세포및간암세포의사멸을유도하고이작용기전에활성산소종 이관여한다는연구결과가발표되었다.
7,12)또한신경아세종에서 는아피제닌이세포막칼륨-
염소이온수송체를활성화시키며 이작용은활성산소종에의해매개된다는새로운사실이발표되었다
.
13)피부의색은피부속에존재하는멜라닌의함량에의해결정 되며피부의기저층에존재하는멜라닌생성세포에의해합성된 멜라닌은세포질돌기를통하여각질형성세포의각화과정에의 해각질층으로이동한다
.
14)멜라닌은태양광선으로부터들어오는자외선을차단하여피부를보호하는역할을담당하고있지만 만일국소적으로과도하게생성되거나
,
노화등에의해피부의 생리기능이떨어지게되면멜라닌이피부표면에침착되어기미,
주근깨또는다양한색소침착을유발하게된다
.
15)멜라닌생성 세포에서멜라닌합성은멜라노좀에서일어나며여기에존재하 는여러cascade
효소반응에의해생성된다.
16,17)이들효소중tyrosinase
는멜라닌합성의속도를결정하는효소로서, tyrosine
을3,4-dihydroxyphenylalanin(DOPA)
로전환하는tyrosine hydroxylase
활성과
DOPA
를DOPA quinone
으로산화하는DOPA oxidase
활성을가지고있다
. Tyrosinase-related protein(TRP)-1
은5,6-
#본논문에관한문의는저자에게로
(
전화) 02-901-8396 (
팩스) 02-901-8386
(E-mail) [email protected]
dihydroxyindole-2-carboxylic acid(DHICA)
를indole-5,6-quinone- 2-carboxylic acid
로산화하는효소이며dopachrome tautomerase (DCT)
는 초기에TRP-2
로불려졌던효소로서dopachrome
을DHICA
로이성화하는효소이다.
멜라닌은흑·갈색의eumlanin
과적·노랑색의
phaeomelanin
으로나뉘는데tyrosinase
는이들 두 가지 타입의melanin
합성에 필요하며, TRP1
과DCT
는eumelanin
의합성에더많이관여하는것으로알려져있다.
18)최근다양한식물추출물이
tyrosinase
활성저해나활성산소종의발 생억제등을통하여멜라닌합성을저해함으로써미백기능성화 장품의원료물질로개발되고있다.
19,20)칼륨
-
염소이온수송체는적혈구에서팽창-
의존성칼륨유출기 전을매개하는기능이있다는사실21,22)이처음밝혀진이래기 능적및생리적인실험적증명을통하여여러조직에존재하고 있다는것이밝혀졌다.
즉,
칼륨-
염소이온수송체는상피,
23,24) 내 피,
25)혈관평활근,
26)심장,
27)골격근28)및신경29)에그존재가확 인되었다.
칼륨-
염소이온수송체는여러생리활성을나타내는데조절체적감소
(regulatory volume decrease),
30)상피세포간염류 흡수,
23)허혈심근에서칼륨손실,
27)신경에서염소농도조절29)및신장에서칼륨분비31)등의기능을가지고있다
.
또한최근본연구실에서도칼륨
-
염소이온수송체가사람간암세포의사멸기전 에중요한매개체로작용하고있음을박힌바있다.
32)본연구에서는이러한기존연구결과를바탕으로하여아피제 닌이멜라닌합성에미치는영향을관찰하고이에미치는세포 막칼륨
-
염소이온수송체의역할을규명하고자하였다.
실험 방법
시약
Dulbecco‘s modified Eagle’s medium(DMEM), 3-(4,5-di- methylthiazol-2-yl)-2,5-diphenyltetrazoliumbromide(MTT), apigenin, Nethylmaleimide(NEM)
및각종용매와염류는Sigma Chemical Co.(
미국)
에서 구입하였고, R-(+)-[(2-n-butyl-6,7- dichloro-2-cyclopentyl-2,3-dihydro-1-oxo-1H-inden-5- yl)oxy]acetic acid(DIOA)
는Alexis Biochemicals(
미국)
에서fetal bovine serum(FBS)
와penicillin-streptomysin
혼합액은GIBCO (
미국)
에서 구입하였으며, N-(6-methoxyquinolyl) acetoethyl ester(MQAE)
및potassium-binding benzofuran isophthalate/
aceoxylmethyl ester(PBFI/AM)
은Molecular Probes, Inc.(
미국)
에서구입하여사용하였다
.
세포배양
B16
흑색종세포는서울대학교한국세포주은행에서구입하였다
.
구입한세포는10% FBS
와penicillin 100 IU/m
l와strep- tomysin 50
µg/m
l을함유한DMEM
용액으로37
oC
로유지되는5% CO
2배양기(Forma,
미국)
에서배양하였다.
세포생존율측정
살아있는세포는미토콘드리아에존재하는탈수소효소에의 해
MTT
가MTT-formazan
으로전환되므로이것의양을재면살아있는세포의수를측정할수있다
.
세포는1 m
l배지에여러가지약물또는약물을녹인용매만을처리하여
24 well plate
에서
48
시간동안배양하였다.
배양이끝난세포에MTT
용액(2.5 mg/m
lH
2O) 100
ml를첨가하여37
oC
에서4
시간동안반응시켰 다.
생성된MTT-formazan
결정체를분리하기위하여세포현탁액을
eppendorf tube
에옮겨원심분리하였다(1500 rpm, 4
분).
상 징액을조심스럽게제거하고DMSO 100
µl를첨가하여결정체 를용해시킨후540 nm
에서ELISA reader(Molecular Device,
미국
)
로흡광도를측정하였다.
세포생존율은대조군과비교하여 흡광도의백분율로나타내었다.
멜라닌정량
B16
세포에서멜라닌을측정할때에는phenol red
가없는DMEM
을사용하여24 well plate
에5
×10
4cells/m
l로분주하고12
시간이지난뒤세포가plate
에완전히부착된것을확인한후시료를처리하고
48
시간지난뒤에세포를수집하여세포수를측정하고
, 1200 rpm
에서5
분간원심분리하여침전한후, 1 m
lhomogenization buffer(50 mM sodium phosphate pH 6.5, 1%
Triton X-100, 2 mM PMSF)
로용해시켰다.
여기서얻은세포pellet
을1 N NaOH(+10% DMSO) 200
µl를 첨가하고vortex
후멜라닌을완전히녹인다음
96 well plate
에옮긴후ELISA reader(Molecular Device,
미국)
를사용하여405 nm
에서흡광도를측정하였다
.
멜라닌표준품을이용한표준검량선을이용하여멜라닌양을산출하였다
.
멜라닌생성량은단위세포(10
4cells)
에서의멜라닌생성량으로비교하였다
.
세포내염소이온농도의측정
세포내염소이온농도의변화는염소이온에민감한형광탐침인
MQAE
를사용해조사하였다.
세포를두번Hank
용액으로씻은 후4
×10
5cells/m
l되게현탁시킨후MQAE
를세포내로봉입시 키기위해최종농도가5 mM
로하여실온에서밤새배양하였다.
형광
(excitation wavelength=365 nm; emission wavelength=
450 nm)
을교반을한상태의cuvette
을이용하여측정하였다.
실 험을실온에서시행하여형광탐침이새어나오는것을최소화하 였다.
실험결과는세포내염소이온과비례하는값인상대적형 광(Fo/F; Fo=
염소이온이없을때형광치; F=
시간에따른형광 치)
으로나타내었다.
모든형광값은MQAE
이온선택적신호를 최대로제거하는
5
µM valinomycin
을 포함하는HEPES-
buffered KSCN
용액을 이용하여따로 측정한 배경 형광치(background fluorescence)
에대하여보정하였다.
다른실험으로Fo
값은10
µM tributyltin
및10
µM nigericin
을포함한염소-
제거
KNO
3용액에세포를현탁시켜측정하였다.
세포내칼륨농도의측정
세포내칼륨농도는형광탐침인
PBFI/AM
을이용하여측정하 였다.
비이온성계면활성제인0.02% pluronic F-127
을포함한Hank
용액의세포현탁액에5
µM
의PBFI/AM
을가해37
oC
에서2
시간동안진탕배양하여세포내로봉입시킨후,
봉입되지않은PBFI/AM
은원심분리하여제거시켰다.
다시세포를Krebs-Ringer
완충액
(125 mM NaCl, 5 mM KCl, 1.2 mM KH
2PO
4, 1.2 mM MgSO
4, 5 mM NaHCO
3, 25 mM HEPES, 6 mM glucose, pH 7.4)
에현탁시킨후cuvette
에옮겨340 nm
및380 nm
파장에서excitation
시켜, 500 nm
파장에서나오는형광을형광분석기로 측정하였다.
이때cuvette
내의 세포현탁액을계속교반하여 세포가가라앉는것을방지하였다.
세포내칼륨농도는두파장에서나온형광세기의비로써나타내었다
.
자료분석및통계적검정
모든실험은네번반복해서실시하고실험결과는대조군의 조건에대한백분율로나타내었다
. Data
는평균값±SEM
으로표시하고
ANOVA
로분석하며 각각의유의성비교는Student-
Newman-Keul's test
를이용하여실시하였다. P
값이0.05
이하인 경우에만통계학적유의성이있는것으로간주하였다.
실험결과 및 고찰
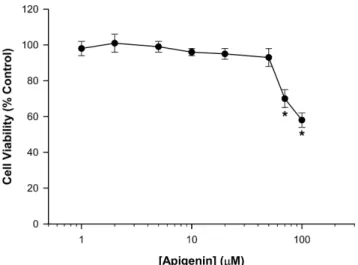
아피제닌의세포독성
아피제닌이세포독성에미치는농도를조사하여향후실험에
사용될농도범위를결정하기위해서
MTT
분석을시행하였다.
B16
흑색종세포에대한아피제닌의세포독성을측정한결과,
아피제닌은
50
µM
이하의농도로처리했을때세포생존율이90%
이상으로나타났으며
, 70
µM
을처리했을때생존율이유의성있게저하되었다
(Fig. 1).
따라서향후실험에서아피제닌은50
µM
이하의농도로사용하였다
.
아피제닌은다른세포에서도본실험에서사용한
B16
세포에서의결과와비슷한양상을보이고있으며33)특히고농도에서다른플라보노이드화합물도세포독성 을나타낸다
.
34)아피제닌의멜라닌생성촉진효과
B16
세포를이용하여아피제닌이멜라닌합성에미치는영향 을확인하였다.
준비된세포에아피제닌을농도별로처리하고48
시간동안배양하였다
. 48
시간을선택한이유는이시간대에서 대조군,
즉약물을가하지않은상태에서분석이가능한멜라닌합성이이루어지기때문이다
.
배양후B16
세포를수집하여멜 라닌양을측정한결과,
아피제닌처리군에서농도의존적으로멜라닌합성이증가되었으며
20
µM
농도처리군에서대조군에비 해약2.2
배가증가하여통계적으로유의한촉진효과를나타내었다
(Fig. 2).
식물유래플라보노이드화합물들이멜라닌합성을Fig. 1 −
Effect of apigenin on cell viability in B16 melanoma cells.
Cells were incubated with or without each concentration of apigenin for 48 hr. Cell viability assay was done by the MTT staining method. Results are expressed as percent change of control condition in which cells were grown in medium without apigenin. Each point represents the mean value of four replications with bars indicating SEM.
*p<0.05 compared to control.
Fig. 2 −
Effects of apigenin on the production of melanin in B16
melanoma cells. Melanin content was measured by the
method described in the Method section. The cells were
initially grown for 12 hr in order to attach them to the
bottom of the culture flasks. Then, the cells were incubated
with or without apigenin at each designated concentration
for 48 hr. Each column represents the mean value of four
replications with bars indicating SEM. *p<0.05 compared
to control.
저해한다는연구결과는많이발표되었다
.
35,36)그러나최근이와반대로퀄세틴과몇몇플라보노이드는오히려멜라닌합성을증 가한다는연구결과도발표되었다
.
37-41)본연구결과에서보여준 아피제닌의멜라닌합성촉진작용은후자에속하는경우로서흥 미로운결과이나플라보노이드화합물의구조적특성과멜라닌 합성에미치는영향사이에어떤상관성이존재하는지에대한 연구는전무한실정이고향후좀더체계적인연구가필요할것 으로사료된다.
아피제닌에의한멜라닌합성촉진에미치는칼륨
-
염소이온수송체의역할
본연구실에서는신경아세포종을이용한실험에서아피제닌이 활성산소종을발생시키고그작용기전에세포막에존재하는칼 륨
-
염소이온수송체가연루되어있다는사실을보고한바있다.
13)따라서후속연구는본연구에서확인한멜라닌합성촉진작용에 미치는칼륨
-
염소이온수송체의역할에대한검정을시도하였다.
실험결과에서아피제닌은
B16
세포에서칼륨-
염소이온수송체활 성유무를검정하는지표인염소이온및칼륨이온의유출을농 도의존적으로나타내었고(Fig. 3),
이작용은칼륨-
염소이온수송체를선택적으로억제한다고알려진약물인
DIOA
42)에의해유 의성있게차단되었다(Figs. 4A
및4B). 50
µM
아피제닌을투여 했을때나타나는칼륨유출양상은30
µM
투여시에비해최대유출은더증가했지만좀더느리고완만한경향을보였다
(Fig.
3B).
이런양상을보이는이유에대해서는명확히알수없으나농도가다른아피제닌에대한칼륨
-
염소이온수송체의활성화기전이다른때문일가능성이있다
.
하지만이추측을명확히하기 위해서는좀더체계적인연구가필요할것이다.
그럼에도불구하고이들결과로부터
B16
세포에는칼륨-
염소이온수송체가기능적으로존재하고있으며플라보노이드인아피제닌에의해활 성화된다는사실을알수있었다
.
칼륨-
염소이온수송체의존재에 대한확증을위해칼륨-
염소이온수송체를직접적으로활성화시킨다고알려진약물인
NEM
30)에의한칼륨-
염소이온수송체의활 성화작용을확인하였다.
결과에서NEM
이100
µM
농도에서현 저한염소이온유출을나타내었으며(Fig. 4A),
이작용은100
µM DIOA
에의해통계적으로유의성있게억제되었다(Fig. 4B).
또한, NEM
은농도의존적으로멜라닌합성을촉진하였는데50
µM
농 도에서대조군에비해유의성있게증가시켰다(Fig. 4C). NEM
에의한멜라민합성촉진작용도칼륨
-
염소이온수송체억제제인DIOA(100
µM)
에의해유의성있게차단되었다(Fig. 4D).
이와더 불어50
µM
아피제닌에의해증가된멜라닌합성도DIOA(100
µ
M)
에의해유의성있게차단되었다(Fig. 4D).
종합적으로이결과는
B16
세포에서아피제닌에의한멜라닌합성촉진작용은칼륨
-
염소이온수송체의활성화를통해서일어난다는사실을시사한다.
멜라닌합성은세포내여러신호전달경로를통하여일어나 는데멜라닌합성관련실험에서흔히사용되는양성대조군화 합물인α
-melanoyte stimulating factor(
α-MSH)
에의한멜라닌합성촉진작용은
cAMP
경로를통하여일어난다고알려져있다.
멜라닌합성에있어율속단계에관여하는
tyrosinase
의발현은cAMP
에의해증가되고,
이러한tyrosinase
생성에는전사인자인microphthalmia-associated transcription factor(MITF)
의발현에 의해 조절된다고 알려져있다.
43)또한MITF
는extracellular signal-regulated kinase(ERK)
에의해인산화되면그발현이저해된다
.
44)따라서본연구의결과에서보여준아피제닌에의한 멜라닌합성촉진작용은칼륨-
염소이온수송체의활성화를통한Fig. 3 −
Effects of apigenin on the Cl
-(A) and K
+(B) fluxes in B16 melanoma cells. In figure (A) the data show changes in intracellular Cl
-concentration as a function of time, measured by using the Cl
--sensitive indicator MQAE. In the data, Fo/F values are directly proportional to intracellular Cl
-concentration. The arrow shows the time point for addition of apigenin at a designated concentration.
In figure (B) the data show changes in intracellular K
+concentration as a function of time, measured by using the K
+-sensitive
indicator PBFI/AM. In the data, PBFI fluorescence ratios are directly proportional to intracellular K
+concentration. The arrow shows
the time point for addition of apigenin at a designated concentration.
칼륨또는염소이온의세포내농도변화가멜라닌합성에관여 하는여러신호전달인자
(cAMP, MITF, ERK)
의발현또는활성을조절함으로써일어날것으로추측할수있다
.
하지만정확히 어느신호전달과정에연관이있는지를밝히기위해서는향후 추가적인연구가필요하다고생각된다.
그럼에도불구하고본연구에서밝힌아피제닌에의한멜라닌 합성촉진작용은백반과같은멜라닌합성장애에의한저색소질 환의치료용도로활용될가치가충분하다고판단된다
.
결 론
다양한채소와과일에함유되어있는플라보노이드인아피제 닌은여러생리활성을나타내는데본연구에서처음으로
B16
흑색종세포에서멜라닌합성을증가시키는작용이있음을확인하 였다
.
본연구실에서는신경세포에서아피제닌의작용이세포막에존재하는칼륨
-
염소이온수송체에의하여매개된다는사실을 밝혔는바본연구에서확인한멜라닌합성촉진작용에미치는이 이온수송체의역할을검정하였다.
실험결과에서아피제닌은B16
세포에서칼륨
-
염소이온수송체활성유무를검정하는지표인염 소이온-
의존성칼륨이온유출을농도의존적으로나타내었고이작용은칼륨
-
염소이온수송체를선택적으로억제하는약물인DIOA
에의해유의성있게차단되었다
.
이결과는B16
세포에는칼륨-
염소이온수송체가기능을하고있으며아피제닌에의해활성화 된다는것을의미한다
.
칼륨-
염소이온수송체를직접적으로활성화시킨다고알려진약물인
NEM
은농도의존적으로염소이온유 출과멜라닌합성을촉진하였고이작용역시칼륨-
염소이온수Fig. 4 −
Effects of DIOA, a specific KCC inhibitor, on the apigenin and NEM, a KCC activator-induced Cl
-efflux (A and B) and melanin
production (B) in B16 melanoma cells. In figure (A) the data show changes in intracellular Cl
-concentration as a function of time,
measured by using the Cl
--sensitive indicator MQAE. In the data, Fo/F values are directly proportional to intracellular Cl
-concentration. The arrow shows the time point for addition of apigenin (50
µM) and NEM (100
µM). In figure (C) melanin content was
measured by the method described in the Method section. The cells were initially grown for 12 hr in order to attach them to the
bottom of the culture flasks. Then, the cells were incubated with or without NEM at each designated concentration for 48 hr. Each
column represents the mean value of four replications with bars indicating SEM. *p<0.05 compared to control. Quantitative changes
(figures B and D) were expressed as percent changes of the maximum decease in Fo/F values (B) and of melanin content (D) induced
by the drug compared to control condition in which the cells were treated with a drug-free vehicle. In these experiments apigenin
(50
µM), NEM (100
µM) and DIOA (100
µM) were used. Each column represents the mean value of four replications with bars
indicating SEM. *p<0.05 compared to apigenin alone.
#p<0.05 compared to NEM alone.
송체억제제인
DIOA
에의해유의성있게차단되었다.
종합적으로이결과는아피제닌은
B16
세포에서칼륨-
염소이온수송체의활성화를통해서멜라닌합성을촉진한다는사실을시사하며백 반과같은멜라닌합성장애에의한저색소질환의치료용도로활 용될가치가있다고판단된다
.
감사의 말씀
본연구는덕성여자대학교
2007
년도교내연구비지원에의해수행되었음
.
문 헌
1) Peterson, J. and Dwyer, J. : Flavonoids: dietary occurrence and biochemical activity. Nutr. Res.
18, 1995 (1988).
2) Lee, J. H., Zhou, H. Y., Cho, S. Y., Kim, Y. S., Lee, Y. S. and Jeong, C. S. : Anti-inflammatory mechanisms of apigenin:
inhibition of cyclooxygenase-2 expression, adhesion of monocytes to human umbilical vein endothelial cells, and expression of cellular adhesion molecules. Arch. Pharm. Res.
30
, 1318 (2007).
3) Rezai-Zadeh, K., Ehrhart, J., Bai, Y., Sanberg, P. R., Bickford, P., Tan, J. and Shytle, R. D. : Apigenin and luteolin modulate microglial activation via inhibition of STAT1-induced CD40 expression. J. Neuroinflammation
5, 41 (2008).
4) Psotová, J., Chlopcíková, S., Miketová, P., Hrbác, J. and Simánek, V. : Chemoprotective effect of plant phenolics against anthracycline-induced toxicity on rat cardiomyocytes. Part III.
Apigenin, baicalelin, kaempherol, luteolin and quercetin.
Phytother. Res.
18, 516 (2004).
5) Caltagirone, S., Rossi, C., Poggi, A., Ranelletti, F. O., Natali, P. G., Brunetti, M., Aiello, F. B. and Piantelli, M. : Flavonoids apigenin and quercetin inhibit melanoma growth and metastatic potential. Int. J. Cancer
87, 595 (2000).
6) Wang, W., Heideman, L., Chung, C. S., Pelling, J. C., Koehler, K. J. and Birt, D. F. : Cell-cycle arrest at G
2/M and growth inhibition by apigenin in human colon carcinoma cell lines.
Mol. Carcinog.
28, 102 (2000).
7) Choi, S. I., Jeong, C. S., Cho, S. Y. and Lee, Y. S. : Mechanism of apoptosis induced by apigenin in HepG2 human hepatoma cells: involvement of reactive oxygen species generated by NADPH oxidase. Arch. Pharm. Res.
30, 1328 (2007).
8) Shindo, K., Saito, E., Sekiya, M., Matsui, T. and Koike, Y. : Antioxidative activity of the flower of Torenia fournieri. Nat.
Med. (Tokyo)
62, 247 (2008).
9) Ma, X., Li, Y. F., Gao, Q., Ye, Z. G., Lu, X. J., Wang, H. P., Jiang, H. D., Bruce, I. C. and Xia, Q. : Inhibition of superoxide anion- mediated impairment of endothelium by treatment with
luteolin and apigenin in rat mesenteric artery. Life Sci.
83, 110 (2008).
10) Panda, S. and Kar, A. : Apigenin (4',5,7-trihydroxyflavone) regulates hyperglycaemia, thyroid dysfunction and lipid peroxidation in alloxan-induced diabetic mice. J. Pharm.
Pharmacol.
59, 1543 (2007).
11) Sim, G. S., Lee, B. C., Cho, H. S., Lee, J. W., Kim, J. H., Lee, D. H., Kim, J. H., Pyo, H. B., Moon, D. C., Oh, K. W., Yun, Y. P. and Hong, J. T. : Structure activity relationship of antioxidative property of flavonoids and inhibitory effect on matrix metalloproteinase activity in UVA-irradiated human dermal fibroblast. Arch. Pharm. Res.
30, 290 (2007).
12) Shukla, S. and Gupta, S. : Apigenin-induced prostate cancer cell death is initiated by reactive oxygen species and p53 activation. Free Radic. Biol. Med.
44, 1833 (2008).
13) Kim, M. H., Jeong, C. S., Yoon, H. R., Kim, G. H. and Lee, Y. S. : Involvement of K
+-Cl
--cotransport in the apigenin- Induced generation of reactive oxygen species in IMR-32 human neuroblastoma cells. J. Appl. Pharmacol.
14, 137 (2006).
14) Seiberg, M. : Kerationcytemelanocyte interaction during melanosome transfer. Piment Cell Res.
14, 236 (2001).
15) Hill, H. Z., Li, W., Xin, P. and Mitchell, D. L. : Melanin: a two edged sword? Pigment Cell Res.
10, 158 (1997).
16) Cabanes, J., Chazarra, S. and GarciaCarmona, F. : Kojic acid, a cosmetic skin whitening agent, is a slowbinding inhibitor of catecholase activity of tyrosinase. J. Pharm. Pharmacol.
46, 982 (1994).
17) Marmol, V. and Beermann, F. : Tyrosinase and related protein in mammalian pigmentation. FEBS Letters
381, 165 (1996).
18) Hearing, V. J. and Tsukamoto, K. : Biochemical control of melanogenesis and melanosomal organization. J. Invest.
Dermatol. Symp. Proc.
4, 24 (1999).
19) Lee, K. E., Sim, G. S., Kim, J. H., Park, S. M., Lee. B. C., Yun, Y. P., Zhang, Y. H. and Pyo, H. B. : Effects of the Scirpi rhizoma on antixodiation and melanogenesis. Yakhak Hoeji
48, 323 (2004).
20) Cho, Y. H., Lee, B. C., Kim, J. H., Kim, J. H., Pyo, H. B., Zhang, Y. H. and Park, H. D. : Effect of Aetemisia anomala S.
Moore on antioxidant activity and melanogenesis, Kor. J.
Pharmacognosy
36, 273 (2005).
21) Cossins, A. R. and Gibson, J. S. : Volume-sensitive transport systems and volume homeostasis in vertebrate red blood cells.
J. Exp. Biol.
200, 343 (1997).
22) Lauf, P. K., Bauer, J., Adragna, N. C., Fujise, H., Zade-Oppen, A. M., Ryu, K. H. and Delpire, E. : Erythrocyte K-Cl cotransport: properties and regulation. Am. J. Physiol.
263, C917 (1992).
23) Amlal, H., Paillard, M. and Bichara, M. : Cl
--dependent NH
4+transport mechanisms in medullary thick ascending limb cells.
Am. J. Physiol.
267, C1607 (1994).
24) Greger, R. and Schlatter, E. : Properties of the basolateral membrane of the cortical thick ascending limb of Henle's loop of rabbit kidney. A model for secondary active chloride transport. Pflugers Arch.
396, 325 (1983).
25) Perry, P. B. and O'Neill, W. C. : Swelling-activated K fluxes in vascular endothelial cells: volume regulation via K-Cl cotransport and K channels. Am. J. Physiol.
265, C763 (1993).
26) Adragna, N. C., White, R. E., Orlov, S. N. and Lauf, P. K. : K- Cl cotransport in vascular smooth muscle and erythrocytes:
possible implication in vasodilation. Am. J. Physiol.
278, C381 (2000).
27) Yan, G. X., Chen, J., Yamada, K. A., Kleber, A. G. and Corr, P. B. : Contribution of shrinkage of extracellular space to extracellular K
+accumulation in myocardial ischaemia of the rabbit. J. Physiol. (London)
490, 215 (1996).
28) Weil-Maslansky, E., Gutman, Y. and Sasson, S. : Insulin activates furosemide-sensitive K
+and Cl
-uptake system in BC3H1 cells. Am. J. Physiol.
267, C932 (1994).
29) Rivera, C., Voipio, J., Payne, J. A., Ruusuvuori, E., Latineen, H., Lamsa, K., Pirvola, U., Saarma, M. and Kaila, K. : The K
+/Cl
-co-transporter KCC2 renders GABA hyperpolarizing during neuronal maturation. Nature
397, 251 (1999).
30) Lauf, P. K., Bauer, J., Adragna, N. C., Fujise, H., Zade-Oppen, A. M., Ryu, K. H. and Delpire, E. : Erythrocyte K-Cl cotransport: properties and regulation. Am. J. Physiol.
263, C917 (1992).
31) Ellison, D. H., Velazquez, H. and Wright, F. S. : Stimulation of distal potassium secretion by low lumen chloride in the presence of barium. Am. J. Physiol.
248, F638 (1985).
32) Kim, J. A., Kang, Y. S. and Lee, Y. S. : Involvement of K
+-Cl
-- cotransport in the apoptosis induced by N-ethylmaleimide in HepG2 human hepatoblastoma cells. Eur. J. Pharmacol.
418, 1 (2001).
33) Wisman, K. N., Perkins, A. A., Jeffers, M. D. and Hagerman, A. E. : Accurate assessment of the bioactivities of redox-active polyphenolics in cell culture. J. Agric. Food Chem.
56, 7831 (2008).
34) Plochmann, K., Korte, G., Koutsilieri, E., Richling, E., Riederer, P., Rethwilm, A., Schreier, P. and Scheller, C. : Structure- activity relationships of flavonoid-induced cytotoxicity on
human leukemia cells. Arch. Biochem. Biophys.
460, 1 (2007).
35) Choi, M. Y., Song, H. S., Hur, H. S. and Sim, S. S. : Whitening activity of luteolin related to the inhibition of cAMP pathway in alpha-MSH-stimulated B16 melanoma cells. Arch. Pharm. Res.
31