DOI 10.17480/psk.2016.60.6.290
B16 흑색종세포에서 아라키돈산에 의한 멜라닌 합성 촉진 효과에 미치는 칼륨-염소이온-수송체의 역할
이 용 수# 덕성여자대학교 약학대학
(Received September 6, 2016; Revised October 18, 2016; Accepted November 15, 2016)
Role of K
+-Cl
--cotranspoter in the Arachidonic Acid-induced Stimulation of Melanin Synthesis in B16 Melanoma Cells
Yong Soo Lee#
College of Pharmacy, Duksung Women's University, Seoul 01369, Korea
Abstract — The effect of arachidonic acid (AA) on melanogenesis was investigated in B16 melanoma cells. AA induced melanogenesis and K+ efflux in a dose-dependent manner, which was significantly inhibited by DIOA, a specific inhibitor of K+-Cl--cotransporter (KCC). These effects of AA were not altered by inhibitors of cyclooxygenase and lipoxygenase.
Treatment with ETYA, a non-metabolizable analog of AA induced the same effects as AA did. Taken together, these results suggest that AA itself induced melanogenesis, and that KCC may mediate the effects of AA.
Keywords □ arachidonic acid, melanogenesis, K+-Cl--cotransporter, B16 melanoma cell
피부 기저층에 존재하는 멜라닌세포(melanocyte)의 멜라노좀 (melanosomes)에서 합성되는 멜라닌은 사람의 피부색을 결정하 는 색소로 작용할 뿐만 아니라 자외선 조사나 독성 약물 및 화 학 물질 등으로 인한 피부 손상을 방어하는 역할과 같은 다양한 기능을 가지고 있다.1) 기미(melasma), 흑색점(lentigo), 모반세포 모반(nevocellular nevi), 악성 흑색종(malignant melanoma) 등 에서는 멜라닌 합성이 과하게 일어나 피부에 과색소침착증 (hyperpigmentation)이 일어나지만, 반대로 멜라닌세포의 기능을 상실하면 백반증(vitiligo)을 유발하게 된다.2)따라서, 피부에서 멜 라닌 합성의 균형을 적절히 조절하는 것이 중요하다.
멜라닌의 합성은 티로시나제(tyrosinase), tyrosinase-related protein 1(TRP-1) 및 tyrosinase-related protein 2(TRP-2) 등 여 러 효소에 의해 일어나며, 이 중 티로시나제가 율속효소(rate-
limiting enzyme)인데, 이 효소는 티로신이 수산화되어 3,4- dihydroxyphenylalanine(DOPA)을 형성하는 반응과 DOPA가 산 화되어 DOPA quinone으로 되는 반응에 관여한다.3)그 후 몇 단 계 반응을 거쳐 최종적으로 멜라닌이 합성된다. 현재 국내 · 외에 서 천연물로부터 티로시나제의 활성을 조절함으로써 색소질환에 효과가 있는 물질을 얻으려는 연구가 많이 이루어지고 있다.4)
Albert 등의 연구에서 보면 포도막(uveal tract)의 염증이 백반 증의 발생과 연관되어 있고,5)다른 연구에서는 염증 반응이 오 히려 과색소침착증과 관련이 있는 것 같다.6)한편, 염증과 관련 된 효소인 phospholipase A27)에 의해 생성되는 아라키돈산 자체 가 기니아픽 피부를 사용한 실험에서 멜라닌 합성을 촉진하고,8) 또한 아라키돈산 대사체가 티로시나제 활성을 증가시킨다는 연 구 결과가 발표되었다.9)하지만, 아직까지 아라키돈산에 의한 멜 라닌 합성 촉진 작용의 기전은 명확하지 않고 이에 관한 연구도 미비한 실정이다.
따라서, 본 연구에서는 B16 흑색종 세포를 이용하여 아라키돈 산이 멜라닌 합성에 미치는 영향을 알아보고 그 작용 기전을 규 명하고자 하였다.
#
Corresponding Author Yong Soo Lee
College of Pharmacy, Duksung Women’s University, Seoul 01369, Korea
Tel.: 02-901-8396 Fax.: 02-901-8386
E-mail: [email protected]
Short Report
종설실험 방법
시약
Arachidonic acid(AA), indomethacin(Indo), nordihydroguaiaretic acid(NDGA), Dulbecco’s modified Eagle’s medium(DMEM), 3- (4,5-dimethylthiazol-2-yl)-2,5-diphenyl tetrazolium bromide(MTT) 및 각종 용매와 염류는 Sigma–Aldrich(미국)에서 구입하였다. R- (+)-[(2-n-butyl-6,7-dichloro-2-cyclopentyl-2,3-dihydro-1-oxo-1H- inden-5-yl)oxy]acetic acid(DIOA)와 5,8,11,14-eicosatetraynoic acid(ETYA)는 BIOMOL Research Laboratories(미국)에서, fetal bovine serum(FBS)와 penicillin-streptomycin 혼합액은 GIBCO(미 국)에서 구입하였다. Potassium-binding benzofuran isophthalate/
aceoxylmethyl ester(PBFI/AM)은 Molecular Probes(미국)에서 구 입하였다. PBFI/AM은 dimethyl sulfoxide(DMSO)에 녹여 농축액 으로 만든 후 물로 희석하여 사용하였다.
세포 배양
실험에 사용한 B16 흑색종 세포의 배양은 과거 본 연구실에 서 보고한 통상적인 방법으로 시행하였다.10) 간단히 설명하면, 서 울대학교 한국세포주은행에서 구입한 B16 흑색종 세포는 10%
FBS를 함유한 DMEM 용액으로 37oC, 5% CO2배양기(Forma, 미국)에서 배양하였다.
세포 생존율 측정
세포 생존율은 MTT 방법을 이용하여 측정하였다.10) 간단히 설 명하면, 여러 가지 약물을 처리하여 48 시간 동안 배양한 후 MTT 용액을 첨가하여 4 시간 더 반응시키고 생성된 MTT-formazan 결 정체를 DMSO로 용해시킨 후 ELISA reader(Molecular Device, 미국)로 측정한 흡광도를 대조군과 비교하여 세포 생존율을 백분 율로 나타내었다.
멜라닌 정량
B16 세포에서 멜라닌의 정량은 통상적인 흡광도 측정법을 이 용하여 시행하였다.10)간단히 설명하면, 시료를 처리한 세포를 48 시간 배양 후 원심분리로 세포 pellet을 얻은 후 DMSO 용액 으로 멜라닌을 녹인 다음, ELISA reader(Molecular Device, 미 국)를 사용하여 흡광도를 측정하고 멜라닌 표준품을 이용하여 그 린 표준 검량선으로 부터 멜라닌 양을 산출하였다.
세포내 칼륨 이온 농도 측정
세포내 칼륨 이온 농도는 형광 탐침인 PBFI/AM을 이용하여 측정하였다.11)비이온성 계면 활성제인 0.02% pluronic F-127을 포함한 Hank 용액의 세포 현탁액에 5 µM의 PBFI/AM을 가해 37oC에서 2 시간 동안 진탕 배양하여 세포내로 봉입시킨 후, 봉
입되지 않은 PBFI/AM은 원심분리하여 제거시켰다. 다시 세포를 Krebs-Ringer 완충액(125 mM NaCl, 5 mM KCl, 1.2 mM KH2PO4, 1.2 mM MgSO4, 5 mM NaHCO3, 25 mM HEPES, 6 mM glucose, pH 7.4)에 현탁시킨 후 cuvette에 옮겨 340 nm 및 380 nm 파장에서 excitation 시켜, 500 nm 파장에서 나오는 형광을 형광 분석기로 측정하였다. 이 때 cuvette 내의 세포 현 탁액을 계속 교반하여 세포가 가라앉는 것을 방지하였다. 세포 내 칼륨 농도는 두 파장에서 나온 형광 세기의 비로써 나타내었다.
자료 분석 및 통계적 검정
실험 결과는 대조군에 대한 백분율로 나타내었으며 네 번 반 복 실험에서 얻은 실험 결과를 평균값±SEM으로 표시하였고 통 계 처리는 ANOVA로 분석한 후 Student-Newman-Keul’s test를 이용하여 각 군끼리의 유의성 유무를 비교하였다. 통계학적 유 의성은 P값이 0.05 이하인 경우로 한정하였다.
실험 결과 및 고찰
아라킨돈산의 세포 독성
B16 흑색종 세포에 대한 아라키돈산의 세포 독성을 측정한 결 과, 20 µM 이하의 농도로 처리했을 때 세포 생존율은 유의성 있 는 변화를 보이지 않았으며, 50 µM 이상을 처리했을 때 생존율 이 유의성 있게 저하되었다(Fig. 1). 따라서, 향후 실험에서 아라 키돈산은 10 µM 이하의 농도로 사용하였다. 사람 흑색종 세포를 사용한 실험에서도 아라키돈산은 본 실험에서 사용한 비슷한 농 도의 범위에서는 독성을 나타내지 않았다.12)
아라키돈산의 멜라닌 생성 촉진 효과
B16 세포를 이용하여 아라키돈산이 멜라닌 합성에 미치는 영 향을 확인하였다. 아라키돈산을 처리하고 48시간 동안 배양 후 그 효과를 관찰하였는데 그 이유는 이 시간대에서 대조군, 즉 약 물을 가하지 않은 상태에서 분석이 가능한 멜라닌 합성이 이루 어지기 때문이다.10)배양 후 B16 세포를 수집하여 멜라닌 양을 측정한 결과, 아라키돈산을 처리한 군에서 농도 의존적으로 멜 라닌 합성이 증가되었으며 5 µM 및 10 µM 농도 처리군에서 대 조군에 비해 각각 약 2.3 및 2.9배가 증가하여 통계적으로 유의 한 촉진 효과를 나타내었다(Fig. 2). 다른 연구에서도 기관-배양 된(organ-cultured) 기니아픽 피부에서 아라키돈산이 멜라닌 합 성을 촉진한다는 사실이 보고되었다.8)
아라키돈산에 의한 멜라닌 합성 촉진 작용에 미치는 칼륨-염소- 수송체의 역할
본 연구실에서는 간세포에서 아라키돈산이 칼륨-염소-수송체 (K+-Cl−-cotransporter)를 활성화시키며,13) B16 세포에서 식물성
플라보노이드 성분인 아피제닌에 의한 멜라닌 합성 촉진 작용이 칼륨-염소-수송체의 활성화를 통해서 일어난다14)는 사실을 밝힌
바 있다.따라서, 아라키돈산에 의한 멜라닌 합성 촉진 작용에 미 치는 칼륨-염소-수송체의 역할을 조사하였다. 이를 위하여 먼저, 칼륨-염소-수송체의 활성 유무를 검정하는 지표로 사용하는 칼륨 이온 유출(K+ efflux)15)에 미치는 아라키돈산의 영향을 관찰하였다.
아라키돈산(5 µM)은 세포내 칼륨이온 농도를 서서히 감소시켰 으며 이 작용은 칼륨-염소-수송체를 선택적으로 억제한다고 알려
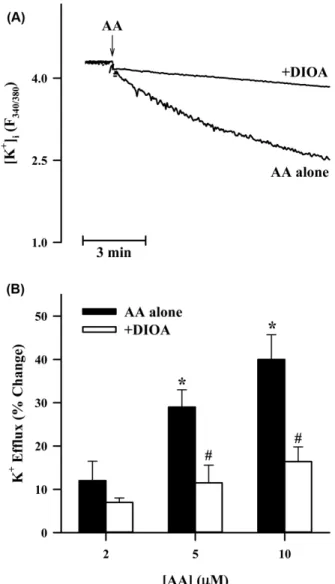
Fig. 3 − Effects of DIOA on the AA-induced K
+efflux in B16 melanoma cells. Intracellular K
+concentration was assessed by PBFI fluorescence probe, and the data (A) represent intracellular K
+changes with time. The arrow shows the time point for addition of AA (5 µM). In the data (B), quantitative changes were expressed as a percentage of the maximum decease in intracellular K
+concentration induced by the drug compared to control condition in which the cells were treated with a drug-free vehicle. In these experiments 100 µM DIOA was used. Each column represents the mean value of four replications with bars indicating SEM.
*P<0.05 compared to negative control.
#P<0.05 compared to AA alone.
Fig. 1 − Effects of AA on cell viability in B16 melanoma cells. Cells were incubated with or without each concentration of AA for 48 hr. Cell viability assay was done by the MTT staining method. Results are expressed as a percentage of control condition in which cells were grown in medium without drugs.
Data points represent the mean values of four replications with bars indicating SEM.
*P<0.05 compared to control in which the cells were incubated with AA-free medium.
Fig. 2 − Effect of AA on the production of melanin in B16
melanoma cells. Melanin content was measured by the
method described in the Method section. Briefly, the cells
were initially grown for 12 hr in order to attach them to the
bottom of the culture flasks. Then, the cells were
incubated with each concentration of AA for 48 hr. Each
column represents the mean value of four replications with
bars indicating SEM.
*P<0.05 compared to control in
which the cells were incubated with AA-free medium.
진 약물인 DIOA(100 µM)16)의 전처리에 의해 확연히 차단되었다 (Fig. 3A). 아라키돈산에 의한 칼륨이온 유출은 농도-의존적으로 나타났고, 5 µM 투여시 통계적으로 유의한 결과를 보였으며, 아 라키돈산의 효과는 DIOA의 전처리에 의해 유의성 있게 차단되 었다(Fig. 3B). 또한, DIOA는 그 자체로는 멜라닌 합성에 영향 을 주지 않지만, 아라키돈산에 의한 멜라닌 합성 촉진 작용에는 농도-의존적으로 차단하는 효과를 나타내었다(Fig. 4). 이 결과는 B16 세포에 칼륨-염소-수송체가 기능적으로 존재하고 있고 아라 키돈산에 의해 활성화되며, 나아가 아라키돈산에 의한 멜라닌 합 성 촉진 작용을 매개한다는 사실을 시사한다.
칼륨-염소-수송체는 적혈구에서 팽창-의존성 칼륨 유출을 매개 한다는 사실17)이 처음 밝혀진 이래, 많은 종류의 세포에서 그 존 재가 확인되었고 그 주된 기능은 세포의 체적을 조절하는 것으 로 알려지고 있다.18)이러한 칼륨-염소-수송체의 기능 변화가 심 혈관계 질환 등 여러 질병의 발병에 관여하고 있다는 사실도 밝 혀지고 있다.19)본 연구실의 실험에서도 사람 간암세포에서 세 포 사멸(apoptosis) 기전에 칼륨-염소-수송체가 중요한 매개체로 작용하고 있음을 밝힌 바 있다.20)본 연구의 결과로써 아피제닌 과 같은 외부에서 투여되는 물질 뿐만 아니라,14)세포 내부에서 생성되는 아라키돈산에 의해서도 칼륨-염소-수송체의 활성이 조 절되며, 이 작용으로 인해 멜라닌 합성이 촉진된다는 새로운 사 실이 확인되었다.
아라키돈산에 의한 멜라닌 합성 촉진 작용에 미치는 아라키돈 산 대사체의 역할
아라키돈산은 포유류의 세포막에 에스테르화되어 존재하며, phospholipase A2(PLA2) 효소에 의하여 sn-2 위치에서 acyl 결합 의 가수분해를 통해 세포내로 유리된다.21)또한, 아라키돈산이 cyclooxygenase(COX) 및 lipoxygenase(LOX)의 작용을 받으면 프 로스타노이드(prostanoid) 및 류코트리엔(leukotriene)이 각각 합 성된다.22)이렇게 생성되는 프로스타글란딘의 한 종류인 PGE2가 멜라닌 합성을 촉진한다는 사실이 보도된 바 있다.23)따라서, 아 라키돈산에 의한 멜라닌 합성 촉진 작용이 아라키돈산 대사체에 의한 것인 지를 확인하기 위하여 COX 및 LOX를 각각 억제하 는 약물인 Indo24)및 NDGA25)를 활용하여 이들 약물이 미치는 영향을 관찰하였다.
Fig. 4 − Effects of DIOA on the AA-induced stimulation of melanin synthesis in B16 melanoma cells. Experiments were done by the same method, as shown in Fig. 2. In these experiments 10 µM AA was used. Each column represents the mean value of four replications with bars indicating SEM.
*P<0.05 compared to negative control.
#
P<0.05 compared to AA alone.
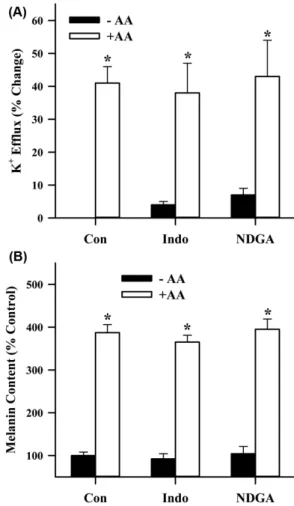
Fig. 5 − Role of AA metabolism in the AA-induced K
+efflux (A) and
melanin synthesis (B) in B16 melanoma cells. Intracellular
K
+concentration and melanin content were assessed by the
techniques, as explained in Fig. 3 and 2, respectively. In the
experiments Indo (30 µM) and NDGA (50 µM) were used
as a COX and LOX inhibitor, respectively. These drugs were
added 5 min before AA (10 µM) application. The data
represent the mean values of four replications with bars
indicating SEM. *p<0.05 compared to negative control.
실험 결과를 보면, Indo(30 µM)와 NDGA(50 µM)를 전처리했 을 때 아라키돈산에 의한 칼륨 유출(Fig. 5A) 및 멜라닌 합성 촉 진 작용(Fig. 5B)이 아라키돈산을 단독으로 처리했을 때와 비교 하여 통계적으로 유의한 변화를 초래하지 않았다.
또한, 아라키돈산과 유사한 작용은 나타내지만 대사는 되지 않 는 물질로 알려진 ETYA26)를 활용하여 이 물질이 아라키돈산과 같이 칼륨 유출과 멜라닌 합성을 촉진하는 지를 조사하였다. ETYA 는 농도-의존적으로 칼륨유출을 증가시켰으며, 이 작용은 칼륨-염 소-수송체 억제제인 DIOA(100 µM)에 의해 효과적으로 차단되었 다(Fig. 6A). 마찬가지로, ETYA는 농도-의존적으로 멜라닌 합성 도 촉진하였으며, 이 작용 역시 DIOA(100 µM)에 의해 효과적으 로 차단되었다(Fig. 6B). 이러한 사실로 미루어 보아 적어도 본 실
험에 사용한 세포에서는 아라키돈산에 의한 칼륨-염소-수송체의 활성화 및 멜라닌 합성 촉진 작용이 그 대사체에 의해서가 아니 라 그 자체의 작용에 의해서 일어난다는 것을 알 수 있다. 본 실 험 결과는 아라키돈산의 대사체인 PGE2가 멜라닌 합성을 유도한 다는 다른 실험실의 결과23)와는 일치하지 않는다. 그 이유에 대 해서는 실험에 사용한 세포 종류의 차이에 기인한 것인 지, 또는 다른 원인에 의한 것인 지는 현재로선 정확히 알 수 없다.
본 연구의 결과에서 아라키돈산에 의한 멜라닌 합성 촉진작용 이 칼륨-염소-수송체의 활성화를 통해서 일어난다는 사실이 확인 되었지만 어떻게 칼륨-염소-수송체의 활성화에 의해 멜라닌 합성 이 촉진될 수 있는 지에 대해선 아직 정확히 밝혀진 바 없다. 다 만, 그 기전으로서 “칼륨-염소-수송체의 활성화에 의한 염소이온 의 세포내 농도 변화가 멜라닌 합성을 촉진한다”는 가설을 세울 수 있다. 왜냐하면, 염소이온이 멜라닌 합성에 중요한 효소인 티로 시나제의 활성을 억제하는 기능을 가지고 있기 때문이다.27)또한, 염소이온 통로인 GABA 수용체28)와 cystic fibrosis transmembrane conductance regulator(CFTR)29)이 멜라닌 합성에 연루되어 있다는 보고도 있다. 따라서, 칼륨-염소-수송체의 활성화에 의한 세포내 염소이온의 유출은 멜라닌 합성을 촉진하는 결과를 초래할 수 있 을 것으로 추측된다.
또 다른 가능성은 “칼륨-염소-수송체의 활성화가 세포막에 존 재하는 NADPH oxidase에 의한 활성산소종의 발생을 유도하여 멜라닌 합성을 촉진한다”는 것이다. 중성구에서 활성산소종을 발 생시켜 살균 작용에 관여하는 NADPH oxidase의 활성이 칼륨- 염소-수송체와 밀접한 관계가 있다는 사실이 보고되었다.30)활성 산소종이 멜라닌 합성을 촉진하며, 활성산소종의 발생을 억제하 는 물질은 자외선 조사에 의한 멜라닌 합성을 차단한다는 사실 은 잘 알려져 있다.31)따라서, 칼륨-염소-수송체의 활성화는 NADPH oxidase에 의한 활성산소종의 발생을 유도하고 이로 인 해 멜라닌 합성이 촉진된다고 추측할 수 있다.
종합적으로, 칼륨-염소-수송체의 활성화와 멜라닌 합성과의 상 관관계를 설명하는 여러 가능성에 대한 확실한 결론을 도출하기 위해서는 향후 이를 입증하는 많은 연구가 필요하다.
결 론
본 연구에서는 아라키돈산이 멜라닌 합성에 미치는 영향을 알 아보고 그 기전을 규명하고자 하였다. 실험 결과에서 아라키돈산 은 B16 세포에서 농도 의존적으로 멜라닌 합성을 증가시켰다. 아 라키돈산은 세포내 칼륨 농도를 용량 의존적으로 감소시켰으며 칼륨-염소-수송체 억제제인 DIOA의 전처리에 의해 유의성 있게 차단되었다. 또한 DIOA는 아라키돈산에 의한 멜라닌 합성 촉진 작용도 농도-의존적으로 억제하였다. 아라키돈산의 이러한 작용 은 COX 및 LOX 억제제인 Indo 및 NDGA에 의해서는 변화가
Fig. 6 − Effects of ETYA, a non-metaboliazable analogue of AA, on
the KCC activation (A) and melanin synthesis (B) in B16
melanoma cells. The data were obtained by the same
methods, as used in Fig. 5. In the experiments 100 µM
DIOA was used. The drug was added 5 min before ETYA
application. The data represent the mean values of four
replications with bars indicating SEM. *p<0.05 compared
to negative control.
#P<0.05 compared to ETYA alone.
없었다. 또한, 대사가 되지 않는 아라키돈산 유사체인 ETYA는 칼륨 유출과 멜라닌 합성을 촉진하여 아라키돈산과 같은 작용을 나타내었다. 이 결과는 아라키돈산에 의한 멜라닌 합성 촉진 작 용이 칼륨-염소-수송체의 활성화에 의해 매개된다는 사실을 시사 한다. 또한, 이 연구의 결과로써 칼륨-염소-수송체는 멜라닌 합성 이상에 의한 피부 색소 질환의 치료제 개발 연구에 중요한 타깃 분자(target molecule)로 활용될 가치가 있다고 판단된다.
감사의 말씀
본 연구는 덕성여자대학교 2016년도 교내연구비 지원에 의해 수행되었음.
References
1) Yamaguchi, Y., Takahashi, K., Zmudzka, B. Z., Kornhauser, A., Miller, S. A., Tadokoro, T., Berens, W., Beer, J. Z. and Hearing, V. J. : Human skin responses to UV radiation: pigment in the upper epidermis protects against DNA damage in the lower epidermis and facilitates apoptosis. FASEB J. 20, 1486 (2006).
2) Slominski, A., Tobin, D. J., Shibahara, S. and Wortsman, J. : Melanin pigmentation in mammalian skin and its hormonal regulation. Physiol. Rev. 84, 1155 (2004).
3) Simon, J. D., Peles, D., Wakamatsu, K. and Ito, S. : Current challenges in understanding melanogenesis: bridging chemistry, biological control, morphology, and function. Pigment Cell Melanoma Res. 22, 563 (2009).
4) Casanola-Martin, G. M., Le-Thi-Thu, H., Marrero-Ponce, Y., Castillo-Garit, J. A., Torrens, F., Rescigno, A., Abad, C. and Khan, M. T. : Tyrosinase enzyme: an overview on a pharmacological target. Curr. Top. Med. Chem. 14, 1494 (2014).
5) Albert, D. M., Nordlund, J. J. and Lerner, A. B. : Ocular abnormalities occurring with vitiligo. Ophthalmol. 86, 1145 (1979).
6) Cardinali, G., Kovacs. D. and Picardo, M. : Mechanisms underlying post-inflammatory hyperpigmentation: lessons from solar lentigo. Ann. Dermatol. Venereol. 139, S148 (2012).
7) Leistad, L., Feuerherm, A. J., Faxvaag, A. and Johansen, B. : Multiple phospholipase A2 enzymes participate in the inflammatory process in osteoarthritic cartilage. Scand. J.
Rheumatol. 40, 308 (2011).
8) Maeda, K., Tomita, Y., Naganuma, M. and Tagami, H. : Phospholipases induce melanogenesis in organ-cultured skin.
Photochem. Photobiol. 64, 220 (1996).
9) Tomita, Y., Maeda, K. and Tagami, H. : Melanocyte-stimulating properties of arachidonic acid metabolites: possible role in postinflammatory pigmentation. Pigment Cell Res. 5, 357 (1992).
10) Lee, Y. S. : Role of Ca2+-activated Cl- channels in the
stimulation of melanin synthesis induced by cyclosporin A in B16 melanoma cells. Yakhak Hoeji 59, 177 (2015).
11) Lee, Y. S. : Role of NADPH oxidase-mediated generation of reactive oxygen species in the apigenin-induced melanogenesis in B16 melanoma cells. Yakhak Hoeji 55, 485 (2011).
12) Zajdel, A., Wilczok, A., Chodurek, E., Gruchlik, A. and Dzierzewicz, Z. : Polyunsaturated fatty acids inhibit melanoma cell growth in vitro. Acta. Pol. Pharm. 70, 365 (2013).
13) Lee, Y. S. : Arachidonic acid activates K+-Cl--cotransport in HepG2 human hepatoblastoma cells. Kor. J. Physiol. Pharmacol.
13, 401 (2009).
14) Lee, Y. S. : Role of K+-Cl--cotransporter in the apigenin-induced stimulation of melanogenesis in B16 melanoma cells. Yakhak Hoeji 52, 500 (2008).
15) Adragna, N. C., Di Fulvio, M. and Lauf, P. K. : Regulation of K+- Cl--cotransport: from function to genes. J. Membr. Biol. 201, 109 (2004).
16) Fujii, T., Ohira, Y., Itomi, Y., Takahashi, Y., Asano, S., Morii, M., Takeguchi, N. and Sakai, H. : Inhibition of P-type ATPases by [(dihydroindenyl)oxy]acetic acid (DIOA), a K+-Cl--cotransporter inhibitor. Eur. J. Pharmacol. 560, 123 (2007).
17) Cossins, A. R. and Gibson, J. S. : Volume-sensitive transport systems and volume homeostasis in vertebrate red blood cells.
J. Exp. Biol. 200, 343 (1997).
18) Lauf, P. K. and Adragna, N. C. : Twenty-five years of K-Cl cotransport: from stimulation by a thiol reaction to cloning of the full-length KCCs. Adv. Exp. Med. Biol. 559, 11 (2004).
19) Adragna, N. C., Ferrell, C. M., Zhang, J., Di Fulvio, M., Temprana, C. F., Sharma, A., Fyffe, R. E., Cool, D. R. and Lauf, P. K. : Signal transduction mechanisms of K+-Cl- cotransport regulation and relationship to disease. Acta Physiol. (Oxf). 187, 125 (2006).
20) Kim, J. A., Kang, Y. S. and Lee, Y. S. : Involvement of K+-Cl-- cotransport in the apoptosis induced by N-ethylmaleimide in HepG2 human hepatoblastoma cells. Eur. J. Pharmacol. 418, 1 (2001).
21) Leslie, C. C. : Regulation of arachidonic acid availability for eicosanoid production. Biochem. Cell. Biol. 82, 1 (2004).
22) Harizi, H., Corcuff, J. B. and Gualde, N. : Arachidonic-acid- derived eicosanoids: roles in biology and immunopathology.
Trends Mol. Med. 14, 461 (2008).
23) Imokawa, G. and Motegi, I. : Skin organ culture model for examining epidermal melanization. J. Invest. Dermatol. 100, 47 (1993).
24) Bakalova, R., Matsura, T. and Kanno, I. : The cyclooxygenase inhibitors indomethacin and rofecoxib reduce regional cerebral blood flow evoked by somatosensory stimulation in rats. Exp.
Biol. Med. 227, 465 (2002).
25) Tang, D. G., Chen, Y. Q. and Honn, K. V. : Arachidonate
lipoxygenases as essential regulators of cell survival and apoptosis. Proc. Natl. Acad. Sci. U.S.A. 93, 5241 (1996).
26) Kehl, S. J. : Eicosatetraynoic acid (ETYA), a non-metabolizable analogue of arachidonic acid, blocks the fast-inactivating potassium current of rat pituitary melanotrophs. Can. J.
Physiol. Pharmacol. 79, 338 (2001).
27) Han, H. Y., Lee, J. R., Xu, W. A., Hahn, M. J., Yang, J. M. and Park, Y. D. : Effect of Cl- on tyrosinase: complex inhibition kinetics and biochemical implication. J. Biomol. Struct. Dyn. 25, 165 (2007).
28) Matthew, E., Laskin, J. D., Zimmerman, E. A., Weinstein, I. B., Hsu, K. C. and Engelhardt, D. L. : Benzodiazepines have high-
affinity binding sites and induce melanogenesis in B16/C3 melanoma cells. Proc. Natl. Acad. Sci. U.S.A. 78, 3935 (1981).
29) Kim, N. H., Cheong, K. A., Lee, T. R. and Lee, A. Y. : PDZK1 upregulation in estrogen-related hyperpigmentation in melasma. J. Invest. Dermatol. 132, 2622 (2012).
30) Sun, Y. T., Shieh, C. C., Delpire, E. and Shen, M. R. : K+-Cl- cotransport mediates the bactericidal activity of neutrophils by regulating NADPH oxidase activation. J. Physiol. 590, 3231 (2012).
31) Funasaka, Y., Komoto, M. and Ichihashi, M. : Depigmenting effect of α-tocopheryl ferulate on normal human melanocytes.
Pigment Cell Res. 13(Suppl. 8), 170 (2000).