Phylogenetic Study of Genus Haliotis in Korea by Internal Transcribed Spacer Sequence (ITS)
Man Kyu Huh*, Jung Ho Kim
1and Du-Ho Moon
1Department of Molecular Biology, Dongeui University, 995 Eomgwangno, Busanjin-gu, Busan 614-714, Korea
1
전체 글
1
수치


관련 문서
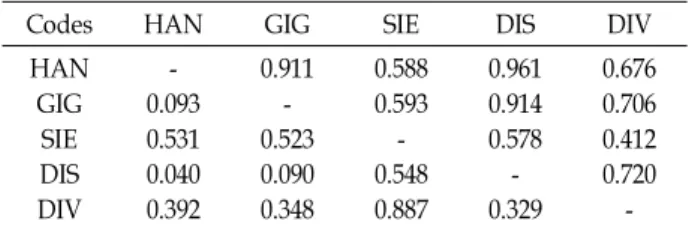
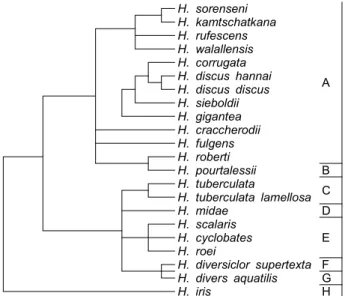
We determined the nucleotide sequences of the mitochondrial DNA (mtDNA) control region using cloning and sequencing, and obtained the complete sequence from the cattle bones
Full genome cloning and nucleotide sequence analysis of hepatitis C virus from sera of chronic hepatitis patients in Korea. Detection of hepatitis C virus RNA
Therefore, in this study, genetic tests were performed to investigate and identify the infections of Orientia tsutsugamushi, Ricekttsia genus, Anaplasma phagocytophilum ,
The Dusky Lory ( Pseudeos fuscata ) is a monotypic species of parrot in the Psittaculidae family, and the only species of the genus Pseudeos... platyrhynchos
Zao Wou-Ki generated the best H1 result for the entire Asian continent in Hong Kong; 53% of Zhang Daqian’s turnover was hammered there, and lots of new
It considers the energy use of the different components that are involved in the distribution and viewing of video content: data centres and content delivery networks
After first field tests, we expect electric passenger drones or eVTOL aircraft (short for electric vertical take-off and landing) to start providing commercial mobility
1 John Owen, Justification by Faith Alone, in The Works of John Owen, ed. John Bolt, trans. Scott Clark, "Do This and Live: Christ's Active Obedience as the
