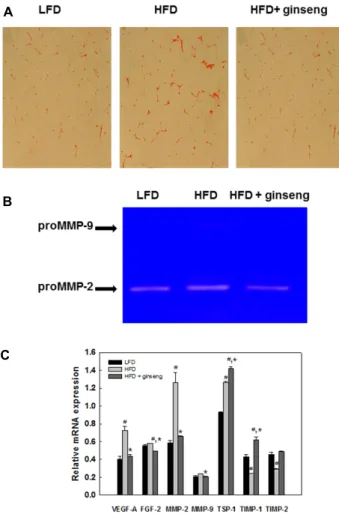
The Role of Angiogenesis in Obesity
15
0
0
전체 글
(2)
(3)
(4)
(5)
(6)
(7)
(8)
(9)
(10)
(11)
(12)
(13)
(14)
(15)
수치

+5


관련 문서
Purpose: TThis study examined the effect of repetitive magnetic stimulation (RMS) on the viability and proliferative response of human adipose tissue‐derived stromal cells
Obesity and Breast Cancer: The Role of Crown-Like Structures in Breast Adipose Tissue in Tumor Progression, Prognosis, and
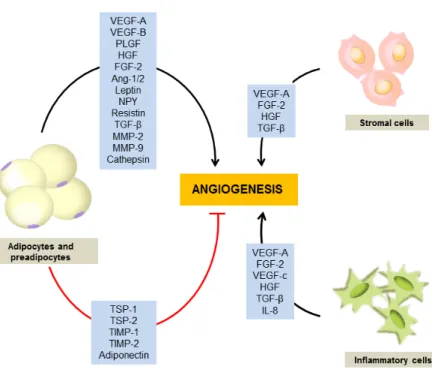
Adipose tissue which used to be simply known as a storage of surplus energy is now perceived as an independent and active endocrine organ, and various kinds
Increased adipose tissue plays an important role in the development of low-grade inflammation, which is characterized by cytokine production and stimulation of inflammatory cytokine
Key words : Human adipose tissue-derived mesenchymal stem cells (hADSCs), osteogenic differentiation, nuclear factor of activated T cells (NFAT5)..
Fig. Tube formation of adipose tissue-derived CD146 positive endothelial cells in DMEM, DMEM with osteogenic inductive agents, EGM-2, and EGM-2 with osteogenic inductive
In the present study, we collected adipose tissue-derived stem cells from adult patients with liver disease, placed them in artificial scaffolds, and implanted these scaffolds in
Normal and healthy adipose tissue harbors various types of anti- inflammatory immune cells such as M2-like macrophages (M2 MAC), eosinophils, helper T type 2 (T H 2) cells, regulatory
