pISSN 1226-4407, eISSN 2234-7631
서 론
비만의 유병률은 전세계적으로 지속적으로 증가하고 있
으며 대유행(epidemic)의 양상을 보이고 있다. 미국의 경우 현재 전체 인구의 60% 이상이 과체중 또는 비만으로 분류 되고 있으며 국내에서도 비만의 유병률은 계속적으로 상승
ꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏꠏ
Corresponding author: Kyung Mook Choi, Division of Endocrinology and Metabolism, Department of Internal Medicine,
Korea University Guro Hospital, Korea University College of Medicine, Guro-dong, Guro-gu, Seoul 152-703, Korea Tel: 82-2-2626-3043, Fax: 82-2-2626-1693, E-mail: [email protected], Mobile: 82-10-2074-3043
* This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/
by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
비만관련 질환 매개자로서의 아디포카인
Adipokines as a Mediator for Obesity-related Disorders
류혜진․최경묵*
고려대학교 의과대학 내과학교실 내분비내과
Hye Jin Yoo, Kyung Mook Choi*
Division of Endocrinology and Metabolism, Department of Internal Medicine, Korea University College of Medicine
요 약
과거 단순히 잉여 에너지의 저장고로만 알려져 있던 지방조 직은 최근 하나의 독립적이고 능동적인 내분비 기관으로 인식 되고 있으며, 지방세포에서 분비되는 단백질인 leptin, tumor necrosis factor-α (TNF-α), resistin, adiponectin 등과 같은 여 러 아디포카인들이 지방대사, 에너지 항상성 및 인슐린 감수성 을 조절하면서 비만 관련 대사 질환의 발생에 주요 영향을 미 치고 있다. 1994년 지방세포에서 렙틴이라는 호르몬이 발견되 면서 지방 조직의 내분비기관으로써의 접근이 시작되었는데, 지방세포는 렙틴과 같은 여러 종류의 아디포카인을 분비함으 로써 자율분비계의 신호전달망을 통한 복잡한 세포 기능 조절 에 관여하며, 또한 뇌하수체, 췌장, 간, 근육, 혈관내피, 면역계 와 같은 다양한 조직에도 영향을 미치는 것으로 알려져 있다.
비만 환자의 경우 이러한 아디포카인의 분비 조절이 비정상적 으로 편향되고, 특정 아디포카인의 분비가 증가 혹은 감소하게 된다. 따라서, 새로운 아디포카인을 발견하고 그 기능을 규명 하는 일은 비만과 연관되어 발생하는 대사 질환의 새로운 치 료 전략 모색을 가능케 할 것으로 기대한다.
중심단어: 아디포카인, 비만, 렙틴, 아디포넥틴
ABSTRACT
Adipose tissue which used to be simply known as a storage of surplus energy is now perceived as an independent and active endocrine organ, and various kinds of adipokine such as leptin (a protein secreted by fat cells), tumor necrosis factor-α (TNF-α), resistin, and adiponectin make major effects on obesity related metabolic diseases by controlling fat metabolism, energy homeostasis and insulin sensitivity. As the hormone called leptin was discovered at fat cells in 1994, it was started to study fat tissues as endocrine organs. Fat cells are engaged in complicated cell function coordination through signaling network of autonomic secretion system by secreting various kinds of adipokine, and are known to make effects on various organizations such as hypophyseal, pancreas, liver, muscle, vascular endothelium, and immune system. In obese patients, the secretion and coordination of such adipokines are biased abnormally and the secretion of specific adipokine becomes increased or decreased. Accordingly, to discover new adipokines and define their functions may enable us to find a new treatment strategy for metabolic disorders related to obesity.
Key words: Adipokines, Obesity, Leptin, Adiponectin
하고 있다. 비만은 인슐린 저항성, 제2형 당뇨병, 고혈압, 이 상지혈증 및 죽상동맥경화증과 같은 여러 가지 대사질환과 병인론적으로 밀접한 연관을 맺고 있다. 1994년 렙틴의 발 견 이후 지방조직은 대표적인 내분비기관으로 자리잡게 되 었으며 지방조직에서 유래한 여러 가지 아디포카인들이 전 신적인 에너지 균형, 포도당 항상성, 지방 대사 및 염증에 핵심적인 역할을 하고 있음이 확인되었다. 본 원고에서는 비만 관련 염증 질환의 매개자 역할을 수행하는 대표적인 아디포카인인 leptin, adiponectin과 새로운 아디포카인인 retinol binding protein 4, adipocyte fatty acid binding protein, lipocalin-2에 관한 실험 및 임상 연구들을 간략하 게 정리하여 보고자 한다.
본 론 1. 렙틴(Leptin)
렙틴은 비만쥐 모델의 ob 유전자의 단백질 산물로 처음 클로닝되었으며 혈중 렙틴은 지방조직에서 주로 생산되고 분비된다.1) 렙틴의 지방조직 발현과 혈중 농도는 비만이 나 타나면서 증가한다. 렙틴의 발현은 식사에 의해 자극되어 증가하며 공복 중에는 혈중 농도가 급속히 감소한다. 인슐 린은 렙틴 발현의 강력한 자극제이며 식후 렙틴 농도 증가 의 주된 원인이 된다. 인슐린 이외에도 당질코르티코이드나 에스트로겐의 장기간 자극은 렙틴의 생산과 분비를 촉진한다.
렙틴은 시상하부에서 식욕유발 펩티드(orexigenic peptide;
neuropeptide Y [NPY] and agouti-related peptide [AgRP]) 의 발현을 감소시키고 식욕부진유발 펩티드(anorexigenic peptide; pro-opiomelanocortin [POMC] and cocaine and amphetamine-regulated transcript [CART])의 발현을 증가 시켜 식욕과 체중 증가를 억제하며 근육과 간에서 세포 내 지방을 감소시킨다.2) 렙틴 농도의 감소는 에너지 섭취를 촉 진하고 많은 에너지를 소모하는 생식(reproduction), 갑상선 에 의한 열발생(thermogenesis) 및 면역 반응 등을 억제한 다. 고지방식이를 한 설치류에서 장기적으로 렙틴을 투여하 면 말초조직의 지방이 감소하고 인슐린 저항성이 개선되는 데 렙틴은 골격근에서 AMPK를 직접적으로 자극하며 지방 산 산화와 지방분해를 촉진하는 것으로 알려져 있다. 하지 만 비만한 환자에서 혈중 렙틴은 오히려 증가하는 것이 관 찰되며 이는 렙틴 작용에 대한 저항성을 시사한다. 설치류 에서 고지방식이를 할 경우 혈중 렙틴 농도가 증가하고 혈 뇌장벽(blood-brain barrier)를 통한 렙틴 수송이 저해되며 간과 골격근 같은 말초조직에서 렙틴의 대사 작용에 대한 저항성이 나타나는 것이 확인되었다.3) 최근 연구에서는 설 치류에서 10주간의 고지방 식이를 하였을 때 렙틴 수용체 단백질량이 감소되는 것이 보고되었다. SOCS3 (suppressor of cytokine signaling 3)는 가장 잘 알려진 렙틴 신호전달
억제제인데 비만한 환자의 골격근에서 SOCS3 mRNA 발현 이 유의하게 증가되고 사람의 근육세포에서 SOCS3를 과발 현시킬 경우 렙틴에 의한 AMPK 자극 효과가 소실되는 것 이 발표된 바 있다.4)
2. 아디포넥틴(Adiponectin)
아디포넥틴은 지방조직에서 처음 클로닝된 30-kDa의 단 백질이다.5) 아디포넥틴은 여러 가지 형태로 존재하는데 기 본적인 형태는 trimer이고 2개의 trimer가 비공유적으로 결 합된 hexamer (medium molecular weight form)와 6 timer 로 구성된 18 mers (high molecular weight form)로도 존재 한다. 특히 High molecular weight 형태의 아디포넥틴은 생 물학적으로 활성을 가지며 인슐린 감수성을 촉진하는데 핵 심적인 역할을 하는 것으로 보이는데 thiazolidinedione에 의해 상향조절되는 주된 아디포넥틴이다. 아디포넥틴은 지 방세포에서 주로 발현하는데 C/EBPα와 PPARγ가 아디포넥 틴 전사를 조절하는 주요 전사인자(transcription factor)이 다. 최근 연구에서 C/EBPα와 Foxo1은 상호작용을 통하여 전사 복합체(transcription complex)를 형성하며 2개의 Foxo1 결합 부위를 통하여 아디포넥틴의 전사를 상향조절 (upregulation)하는 것으로 보고되었다. 아디포넥틴 수용체 1과 2 (AdipoR1과 AdipoR2)가 아디포넥틴의 생물학적 작 용과 신호전달을 담당하는 것으로 알려져 있는데 AdipoR1 은 전신적으로 발현하지만 골격근에서 가장 많이 발현되고 AdipoR2는 간에서 주로 발현된다.6) AMPK는 아디포넥틴 에 의해 활성화되어 아디포넥틴 신호전달체계의 중요한 역 할을 담당하는 중요한 매개체이다. 아디포넥틴은 골격근에 서 AMPK의 인산화를 통하여 지방산 산화를 촉진하며 결 과적으로 acetyl CoA carboxylase (ACC)를 자극한다. 또한 장기적인 아디포넥틴의 투여는 미토콘드리아의 생합성 및 지방산 산화에 관련된 효소를 증가시킨다. Yamauchi 등은 AdipoR1/R2 KO 생쥐에서 인슐린 저항성이 나타나며 아데 노바이러스를 통하여 간에서 AdipoR1 또는 AdipoR2를 과 발현시켰을 때 AMPK 활성도 및 PPARα 발현 증가를 통하 여 포도당 대사가 개선되는 것을 보고하였다.6) 한편 아디포 넥틴 결핍 생쥐는 인슐린 저항성과 내당능장애를 나타낸다.
In vivo 및 in vitro 실험에서 아디포넥틴은 인슐린 감수성 을 개선하고 지방산 산화를 증가시키며 포도당 섭취를 증가 시키고 간 포도당 생산을 억제하는 것으로 알려져 있다.7) 최근에는 인슐린 감수성 개선 작용 외에 in vivo 실험에서 아디포넥틴이 췌장의 인슐린 분비를 자극하여 포도당 대사 를 개선하는 작용이 보고되었다.8) 또한 아디포넥틴은 중추 신경계에 작용하여 식이 섭취와 에너지 소모를 조절하는 작 용도 가지는 것으로 보인다.9) 현재까지 비만, 제2형 당뇨병 및 지방간 등의 다양한 대사 질환과 아디포넥틴과의 연관성 을 단면적, 전향적으로 살펴본 많은 임상 연구가 진행되었
다. 본 교실에서는 60세 이상의 노인 372명을 3년간 추적 관찰하였을 때, 기저 아디포넥틴 농도가 하위 삼분위에 속 한 군이 가장 높은 삼분위에 속한 군에 비하여 연령과 고지 혈증을 보정한 후에도 대사 증후군 발생 위험도가 2.7배 가 량(P = 0.031; 95% confidence interval [CI], 0.94~6.70) 증가함을 보고하였다.10)
3. Retinol binding protein 4 (RBP4) 지방조직에 특이적으로 GLUT4 발현이 감소될 경우 전신적인 인슐린 저항성이 발생하는 점에 착안해서 Khan 등은 지방조직에서 GLUT4 발현이 특이적으로 증가된 transgenic mice (adip-GLUT4-Tg)와 GLUT4 발현이 감소 된 transgenic mice (adip-GLUT4-/-)를 제작하였다. 특히 adip-GLUT4-/- 쥐들은 근육의 GLUT4 발현이 정상으로 유 지되는 데에도 불구하고 인슐린 저항성을 나타내었으며 지 방조직에서 혈중으로 분비된 물질이 근육의 인슐린 저항성 을 유발하는 것으로 추정되었다. 이에 따라 Yang 등은 이들 transgenic mice의 지방조직을 이용하여 oligonucleotide microarray를 실시하여 adip-GLUT4-/- mice에서 발현이 증 가하는 유전자들을 발견하였는데 이 중 하나가 retinol binding protein 4 (RBP4)로 이전에는 혈중에서 retinol의 운반을 담당하는 단백질로만 알려져 있었다.11) 예상한 바와 같이 adip-GLUT4-/- mice의 지방조직에서의 RBP4 mRNA 가 2.3배 증가한 것이 확인되었을 뿐 아니라 혈청 RBP4도 대조군에 비해 2.5배 증가된 것이 발견되었다. 뿐만 아니라 고지방식이를 한 쥐와 유전적으로 비만한 ob/ob 쥐에서도 마찬가지로 혈청 RBP4의 증가가 확인되었다. 또한 PPAR-γ 작용제인 rosiglitazone을 투여하였을 경우 adip-GLUT4-/- mice에서 증가되었던 RBP4 mRNA의 발현과 혈청 RBP4 의 증가가 모두 정상화되는 것이 관찰되었다. 이와 더불어 recombinant human RBP4 (300 μg/d at 8-10 h intervals) 를 정상쥐에게 투여하였을 때에도 인슐린 저항성과 내당능 장애가 나타났다. 이러한 결과들은 RBP4가 제2형 당뇨병의 발병과 연관된 아디포카인이라는 점을 보여 준다. 이와 반 대로 RBP4의 발현이 소실된 transgenic mice의 경우에는 인슐린 저항성이 개선되는 효과가 나타났다. 이러한 효과들 을 일으키는 기전으로 RBP4가 골격근에서 포도당 흡수를 저해하고 간에서 포도당 생산을 촉진하는 것이 확인되었다.
이 후 이 들은 추가 연구를 통해서 혈중 RBP4가 비만환자 와 비만한 당뇨병환자에서 대조군에 비해 유의하게 증가하 며 euglycemic clamp 연구에서 glucose disposal rate와 역 비례하는 것을 관찰하였다. 또한 하루 60분씩 일주일에 3회, 4주간 운동을 실시한 결과 인슐린 저항성이 개선된 그룹에 서는 혈청 RBP4 농도가 감소되었으나 인슐린 저항성이 변 화가 없는 그룹에서는 RBP4의 농도 역시 변화가 없었다고 보고하였다.12) 국내에서도 RBP4에 대한 연구가 활발한 편
이며 Cho 등은 혈장 RBP4 농도가 한국인 내당능장애 환자 또는 제2형 당뇨병 환자들에서 유의하게 증가됨을 확인한 바 있었다.13) 또한 Choi 등은 이전에 임신성 당뇨병을 가졌 던 환자들에서 내당능 장애의 정도가 RBP4 농도 증가와 연 관됨을 보고하였으며14), Lim 등은 한국인 여성에서 운동으 로 인한 인슐린 저항성 개선 효과가 RBP4 감소와 관련되는 것을 관찰하였다.15) 본 교실의 Seo 등도 혈청 RBP4 농도가 non-alcoholic fatty liver disease (NAFLD) 및 간 효소 수 치와 연관됨을 보고한 바 있다.16)
4. Adipocyte Fatty-acid Binding Protein (A-FABP)
Adipocyte fatty acid binding protein (A-FABP)은 포화, 불포화 지방산과 결합한 후 다양한 염증 반응 경로와 대사 경로로 지방산의 분포를 조율하여 주는 세포질 내 단백질로 알려져 왔다.17) Xu 등에 의해 세포질 내에 고정되어 있는 것으로만 알려져 있던 a-FABP가 혈청으로 분비되며, 혈청 a-FABP 농도가 대사 증후군의 구성 요소들과 유의한 상관 관계가 있음이 밝혀지면서18), 대사 증후군에 있어서의 a-FABP의 역할이 새롭게 조명되기 시작하였다. 1996년 Hotamisligil 등은 a-FABP 에 변이를 일으킨 생쥐(aP2-/- mice)에 고지방 식이를 섭취시킬 경우 대조군 생쥐 (aP2+/+mice)에 비하여 혈장 내 인슐린 농도 및 중성지방 수 치가 유의하게 낮았으며 인슐린 저항성과 관계 있는 TNF-α 의 지방 세포에서의 발현이 감소되어 있음을 확인하였다.19) 실제로 Uysal 등은 경구 당부하 검사를 통하여 aP2-/- ob/ob mice가 aP2+/+ ob/ob mice에 비하여 인슐린 감수성이 향상 됨을 증명하였다.20) 또한 apolipoprotein E의 형질 변화를 일으켜 죽상 동맥 경화증이 발생하도록 한 쥐에 a-FABP의 형질 변이를 유발하였을 경우 죽상 동맥 경화증의 발생이 유의하게 감소함을 확인함으로써 a-FABP가 인슐린 저항성 뿐 아니라 심혈관 질환의 발생에도 의미 있는 역할을 수행 할 것이라 추정케 하였다.21) 흥미로운 점은 a-FABP가 지방 세포뿐 아니라 대식세포에도 분포하면서 죽상 동맥 경화증 의 병인에 지방 세포와 더불어 주요한 영향을 미치고 있다는 점이다. Makowski 등에 의하면 a-FABP를 발현 억제시킨 대식세포의 경우, 콜레스테롤을 방출시키는 PPAR-LXRα -ABCA1 반응 경로가 항진되어 대식세포 내 콜레스테롤의 침착을 저해하는 동시에 IκB, NF-αB 등 주요 염증 반응을 매개하는 효소의 활성은 감소시켜 TNF-α, interleukin-6, monocyte chemoattractant protine-1과 같은 여러 염증 싸이 토카인(inflammatory cytokines)의 생성을 억제함으로써 죽 상 동맥 경화증의 발생을 감소시킨다고 설명하고 있다.22) 이렇듯 a-FABP는 인슐린 저항성과 심혈관 질환의 병인에 주요한 역할을 수행하며, 지방 세포와 대식 세포간의 염증 반응을 매개하는 중요한 분자적 연결 고리로 생각된다.
2006년도에 최초로 홍콩의 Xu 등에 의하여 혈장내의 a-FABP의 농도와 대사증후군의 여러 임상 지표와의 연관 성이 있다고 보고한 연구18) 이후로 아시아인이 아닌 코카시 아인에서도 동일한 적용이 가능하다는 연구 발표가 있었으 며23), 10년 추적 관찰 시 기초 FABP4 값이 당뇨 발생 예측 에 유의한 독립적 인자라는 연구도 있었다.24) 실제로 죽상 동맥 경화증의 지표인 경동맥 내중막 두께와 혈장 내 a-FABP값이 유의한 연관성도 보고되었으며25) 비만 아동에 게 체중감량을 시켰을 경우 a-FABP의 혈장 내 농도가 유의 하게 감소됨을 확인하는 등26) 비만과 연관된 염증 질환의 임상 지표로서의 a-FABP4의 유용성에 대하여 추가 연구들 이 보고되고 있다.
한편, Furuhashi 등은 A-FABP의 유전자인 aP2를 특이 적으로 강력하게 억제할 수 있는 경구용 약제(small- molecule inhibitor)를 개발하여 이를 투여한 결과, 고지방식 이를 한 ApoE-/- mice에서 동맥경화를 억제하는 효과와 여 러 가지 비만과 당뇨 모델 쥐에서 인슐린 저항성을 개선하 고 혈당을 감소시키는 효과를 나타냈다고 보고한 바 있어27), 향후 약제로의 이용 가능성도 기대 된다.
5. Lipocalin-2
Lipocalin family에 속하는 단백질들은 lipocalin folds 라고 불리는 공통적인 3차원 구조를 공유한다. 그 중 lipocalin-2는 neutrophil gelatinase-associated lipocalin (NGAL)라고도 알려진 물질로 특정 박테리아의 주요한 성 장 인자로 작용하는 철의 이송에 결정적인 역할을 하는 siderophores와 결합해 버림으로써 박테리아의 번식을 억제 하는 것을 기전으로 생체 내 면역 반응에 관여하는 것으로 알려져 있었다.28) 2007년 Wang 등이 lipocalin-2가 신장 내
피 세포뿐 아니라 대사 증후군을 조절하는 주요 기관인 지 방과 간에서도 풍부히 발현된다는 사실에 입각하여 대사 증 후군의 환자들의 혈청과 db/db mouse 및 고지방 식이 비만 쥐의 지방 조직에서 lipocalin-2의 발현이 대조군에 비해 유 의하게 증가되어 있고, 이러한 증가가 PPAR-γ Agonist 에 의해 감소되는 것을 확인함으로써 대사 증후군 병인에 관여 하는 지방유래 단백질로 lipocalin-2를 재조명하기 시작하였 다.29) Yan 등은 lipocalin-2가 3T-3L1 cell line에서 TNF와 dexamethasone 처리시 발현이 유의하게 증가되고 PPAR-γ Agonist를 처리하면 발현이 오히려 감소함을 확인하였으며, lipocalin-2를 주입하였을 때 H4IIe 간세포에서 포도당의 생 성이 증가함을 보여줌으로써 lipocalin-2가 인슐린 저항성을 유발하는 새로운 아디포카인일 가능성을 제시하였다.30) Lipocalin-2에 대한 Roudkenar 등의 연구에 의하면 간모세 포종(human hepatoblastoma)로부터 얻어진 HepG2 세포에 방사선 조사를 통하여 활성 산소(reactive oxygen species, ROS)를 유발하였을 때 세포 내 lipocalin-2의 발현이 증가 하였으며31), lipocalin-2의 혈중 값이 관상 동맥 스텐트 삽입 술 후 조영제 유발 신 손상이 발생할 가능성에 대한 조기 표지자임을 밝힌 연구도 있어31), 현재로서 lipocalin-2는 산 화 스트레스에 대한 생체 내 표지자 혹은 조기 염증 반응을 매개하는 주요 유인자(chemoattractant)로 여겨지고 있다.
본 교실에서도 한국인에서 lipocalin-2 농도가 성별과 체질 량지수를 보정한 이후에도 체중, 공복 인슐린, 인슐린 저항 성 지표 등과 양의 연관성을 보이고 HDL 콜레스테롤과 음 의 연관성을 보이는 점을 확인하였고, 더 나아가 관상동맥 질환을 가진 환자에서 대조군에 비해 유의하게 혈중 lipocalin-2 농도가 유의하게 증가함을 보고한 바 있다.32)
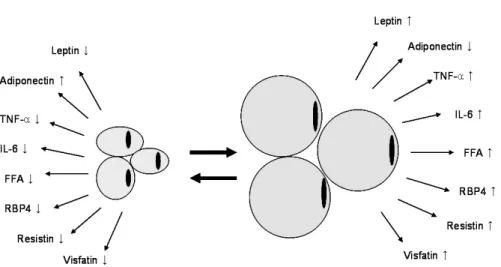
Fig. 1. The changes of adipokines in obesity.
결 론
지방조직은 단순한 과잉 에너지의 저장소를 넘어서 지방 조직에서 유래한 아디포카인들을 통하여 전신적인 대사에 핵심적인 역할을 담당하고 있다. 향후 아디포카인에 대한 보다 심층적인 연구를 통하여 비만과 연관된 인슐린 저항성 과 제2형 당뇨병의 증가에 대한 중요한 기전과 새로운 치료 방법을 발견할 수 있을 것으로 기대된다.
참 고 문 헌
1. Zhang Y, Proenca R, Maffei M, Barone M, Leopold L, Friedman JM. Positional cloning of the mouse obese gene and its human homologue. Nature 1994;372:425-32.
2. Schwartz MW, Woods SC, Porte DJ, Seeley RJ, Baskin DG. Central nervous system control of food intake. Nature 2000;404:661-71.
3. Caro JF, Kolaczynski JW, Nyce MR, Ohannesian JP, Opentanova I, Goldman WH, et al. Decreased cerebrospinal-fluid/serum leptin ratio in obesity: a possible mechanism for leptin resistance. Lancet 1996;348:159-61.
4. Steinberg GR, McAinch AJ, Chen MB, O'Brien PE, Dixon JB, Cameron-Smith D, et al. The suppressor of cytokine signaling 3 inhibits leptin activation of AMP-kinase in cultured skeletal muscle of obese humans. J Clin Endocrinol Metab 2006;91:3592-7.
5. Maeda K, Okubo K, Shimomura I, Funahashi T, Matsuzawa Y, Matsubara K. cDNA cloning and expression of a novel adipose specific collagen-like factor, apM1 (AdiPose Most abundant Gene transcript 1). Biochem Biophys Res Commun 1996;221:286-9.
6. Yamauchi T, Kamon J, Ito Y, Tsuchida A, Yokomizo T, Kita S, et al. Cloning of adiponectin receptors that mediate antidiabetic metabolic effects. Nature 2003;423:762-9.
7. Yamauchi T, Kamon J, Minokoshi Y, Ito Y, Waki H, Uchida S, et al. Adiponectin stimulates glucose utilization and fatty-acid oxidation by activating AMP-activated protein kinase. Nat Med 2002;8:
1288-95.
8. Lee YH, Magkos F, Mantzoros CS, Kang ES. Effects of leptin and adiponectin on pancreatic beta-cell function. Metabolism 2011;60:1664-72.
9. Stofkova A. Leptin and adiponectin: from energy and
metabolic dysbalance to inflammation and autoimmunity. Endocr Regul 2009;43:157-68.
10. Choi KM, Lee J, Lee KW, Seo JA, Oh JH, Kim SG, et al. Serum adiponectin concentrations predict the developments of type 2 diabetes and the metabolic syndrome in elderly Koreans. Clin Endocrinol (Oxf) 2004;61:75-80.
11. Yang Q, Graham TE, Mody N, Preitner F, Peroni OD, Zabolotny JM, et al. Serum retinol binding protein 4 contributes to insulin resistance in obesity and type 2 diabetes. Nature 2005;436:356-62.
12. Graham TE, Yang Q, Bluher M, Hammarstedt A, Ciaraldi TP, Henry RR, et al. Retinol-binding protein 4 and insulin resistance in lean, obese, and diabetic subjects. N Engl J Med 2006;354:2552-63.
13. Cho YM, Youn BS, Lee H, Lee N, Min SS, Kwak SH, et al. Plasma retinol-binding protein-4 concentrations are elevated in human subjects with impaired glucose tolerance and type 2 diabetes. Diabetes Care 2006;29:2457-61.
14. Choi SH, Kwak SH, Youn BS, Lim S, Park YJ, Lee H, et al. High plasma retinol binding protein-4 and low plasma adiponectin concentrations are associated with severity of glucose intolerance in women with previous gestational diabetes mellitus. J Clin Endocrinol Metab 2008;93:3142-8.
15. Lim S, Choi SH, Jeong IK, Kim JH, Moon MK, Park KS, et al. Insulin-sensitizing effects of exercise on adiponectin and retinol-binding protein-4 concentrations in young and middle-aged women. J Clin Endocrinol Metab 2008;93:2263-8.
16. Seo JA, Kim NH, Park SY, Kim HY, Ryu OH, Lee KW, et al. Serum retinol-binding protein 4 levels are elevated in non-alcoholic fatty liver disease. Clin Endocrinol (Oxf) 2008;68:555-60.
17. Makowski L, Hotamisligil GS. Fatty acid binding proteins--the evolutionary crossroads of inflammatory and metabolic responses. J Nutr 2004;134:2464S-8S.
18. Xu A, Wang Y, Xu JY, Stejskal D, Tam S, Zhang J, et al. Adipocyte fatty acid-binding protein is a plasma biomarker closely associated with obesity and metabolic syndrome. Clin Chem 2006;52:405-13.
19. Hotamisligil GS, Johnson RS, Distel RJ, Ellis R, Papaioannou VE, Spiegelman BM. Uncoupling of obesity from insulin resistance through a targeted mutation in aP2, the adipocyte fatty acid binding
protein. Science 1996;274:1377-9.
20. Uysal KT, Scheja L, Wiesbrock SM, Bonner-Weir S, Hotamisligil GS. Improved glucose and lipid metabolism in genetically obese mice lacking aP2.
Endocrinology 2000;141:3388-96.
21. Makowski L, Boord JB, Maeda K, Babaev VR, Uysal KT, Morgan MA, et al. Lack of macrophage fatty-acid-binding protein aP2 protects mice deficient in apolipoprotein E against atherosclerosis. Nat Med 2001;7:699-705.
22. Makowski L, Brittingham KC, Reynolds JM, Suttles J, Hotamisligil GS. The fatty acid-binding protein, aP2, coordinates macrophage cholesterol trafficking and inflammatory activity. Macrophage expression of aP2 impacts peroxisome proliferator-activated receptor gamma and IkappaB kinase activities. J Biol Chem 2005;280:12888-95.
23. Stejskal D, Karpisek M. Adipocyte fatty acid binding protein in a Caucasian population: a new marker of metabolic syndrome? Eur J Clin Invest 2006;36:621-5.
24. Tso AW, Xu A, Sham PC, Wat NM, Wang Y, Fong CH, et al. Serum adipocyte fatty acid binding protein as a new biomarker predicting the development of type 2 diabetes: a 10-year prospective study in a Chinese cohort. Diabetes Care 2007;30:2667-72.
25. Yeung DC, Xu A, Cheung CW, Wat NM, Yau MH, Fong CH, et al. Serum adipocyte fatty acid-binding protein levels were independently associated with carotid atherosclerosis. Arterioscler Thromb Vasc Biol 2007;27:1796-802.
26. Reinehr T, Stoffel-Wagner B, Roth CL. Adipocyte fatty acid-binding protein in obese children before and after weight loss. Metabolism 2007;56:1735-41.
27. Furuhashi M, Tuncman G, Gorgun CZ, Makowski L, Atsumi G, Vaillancourt E, et al. Treatment of diabetes and atherosclerosis by inhibiting fatty-acid-binding protein aP2. Nature 2007;447:959-65.
28. Flo TH, Smith KD, Sato S, Rodriguez DJ, Holmes MA, Strong RK, et al. Lipocalin 2 mediates an innate immune response to bacterial infection by sequestrating iron. Nature 2004;432:917-21.
29. Wang Y, Lam KS, Kraegen EW, Sweeney G, Zhang J, Tso AW, et al. Lipocalin-2 is an inflammatory marker closely associated with obesity, insulin resistance, and hyperglycemia in humans. Clin Chem 2007;53:34-41.
30. Yan QW, Yang Q, Mody N, Graham TE, Hsu CH, Xu Z, et al. The adipokine lipocalin 2 is regulated by obesity and promotes insulin resistance. Diabetes 2007;56:2533-40.
31. Roudkenar MH, Kuwahara Y, Baba T, Roushandeh AM, Ebishima S, Abe S, et al. Oxidative stress induced lipocalin 2 gene expression: addressing its expression under the harmful conditions. J Radiat Res 2007;48:39-44.
32. Choi KM, Lee JS, Kim EJ, Baik SH, Seo HS, Choi DS, et al. Implication of lipocalin-2 and visfatin levels in patients with coronary heart disease. Eur J Endocrinol 2008;158:203-7.