Sclerotiorin: a Novel Azaphilone with Demonstrated
Membrane Targeting and DNA Binding Activity against
Methicillin-Resistant
Staphylococcus aureus
Chakradhar Dasagrandhi1*, Anup Pandith2,3, and Khalid Imran1
1Department of Microbiology, Krupanidhi Degree College, Varthur (Hobli), Off Sarjapur Road, Carmelaram Post, Bangalore 560035, India 2Department for Management of Science and Technology Development, Ton Duc Thang University, Ho Chi Minh City, Vietnam
3Faculty of Applied Science, Ton Duc Thang University, Ho Chi Minh City, Vietnam
Received: February 13, 2020 / Revised: March 6, 2020 / Accepted: April 11, 2020
Introduction
Staphylococcus aureus (S. aureus), a versatile opportu-nistic pathogen has been implicated in many soft tis-sues, implant related diseases. Emergence of multi-drug resistance in methicillin-resistant Staphylococcus aureus (MRSA) is a serious public health concern. Further development of more virulent forms of S. aureus causes increased healthcare burden [1]. The
out-breaks of several superbugs in the recent past in several countries emphasized once again the need for the devel-opment of novel antimicrobial resources [2]. With the continuous withdrawal of pharmaceutical companies from the antibiotic market and discovery process, there is a great demand for novel antibiotic agents against S. aureus are necessary. Although several natural products of plant, and microbial origin have been reported against MRSA [3], relatively very few molecules are currently under clinical study. Hence there is tremendous oppor-tunity for novel natural products with diverse mecha-nism of action to expand the drug arsenal and to contravene the rise of multidrug resistant MRSA.
The emergence of multi-drug resistant, pathogenic methicillin-resistant Staphylococcus aureus (MRSA) is a threat to global health and has created a need for novel functional therapeutic agents. In this study, we evaluated the underlying mechanisms of the anti-MRSA effect of an azaphilone pigment, sclerotiorin (SCL) from Penicillium sclerotiorum. The antimicrobial activity of SCL was evaluated using agar disc dif-fusion, broth microdilution, time-kill assays and biophysical studies. SCL exhibits selective activity against Gram positive bacteria including MRSA (range, MIC = 128-1028 µg/ml) and exhibited rapid bacteri-cidal action against MRSA with a > 4 log reduction in colony forming units within three hours of adminis-tration. Biophysical studies, using fluorescent probes and laser or electron microscopy, demonstrated a SCL dose-dependent alternation in membrane potential (62.6± 5.0.4% inhibition) and integrity (> 95± 2.3%), and the release of UV260 absorbing materials within 60 min (up to 3.2 fold increase, p < 0.01) of exposure. Further, SCL localized to the cytoplasm and hydrolyzed plasmid DNA. While in vitro checker-board studies revealed that SCL potentiated the antimicrobial activity of topical antimicrobials such as polymixin, neomycin, and bacitracin (Fractional Inhibitory Concentration Index range, 0.26-0.37). Taken together these results suggest that SCL targets the membrane and DNA of MRSA to facilitate its anti-MRSA antimicrobial effect.
Keywords: Azaphilones; sclerotiorin, antimicrobial agents, MRSA, membrane action, synergy
*Corresponding author
Tel: +91-9740696123
E-mail: [email protected]
Fungal azaphilones are fungal pigments with wide health functional benefits. The biological activities of fungal azaphilones have been extensively reviewed [4]. Fungal azaphilones are known to possess inhibitory effect against nematodes, monoamine oxidases, choles-terol acetyl transferases, HIV replication, free radicals, gp120−CD4 binding, heat shock protein 90 (Hsp90), and protein-protein interactions [5−11]. Sclerotiorin (SCL), a azaphilone pigment produced by Penicillium species was reported to exhibit endothelin receptor binding [12]), anti lipoxygenase, aldose reductase [13], anti cholesterol [14], tumor suppressive [15], anti-HIV [16], antifungal [17], anti-mycobacterial [18] properties. The food com-patible SCL analogs have been reported earlier [19]. Although SCL was reported to be antibacterial, details on its mechanism of action was lacking.
SCL has been shown to exhibit some antimicrobial activities. However, a systematic study on the antimicro-bial action of SCL has not been documented.
The present study which was carried out in order to address the potential use of novel non-antibiotic natural product alternatives against MRSA infections, we evalu-ated the anti-microbial activity of SCL isolevalu-ated from Penicillium sclerotiorum and evaluated mechanism of action against methicillin-sensitive S. aureus (MSSA) and methicillin-resistant S. aureus (MRSA).
Materials and Methods
Chemicals and reagents
Antibiotics (nisin, polymixin, neomycin, bacitracin), ethidium bromide (EtBr), propidium iodide (PI), 4′,6-diamidino-2-phenylindole (DAPI), and dimethyl sulfox-ide (DMSO) were purchased from Sigma-Aldrich (USA). Thin layer chromatography (TLC) plates (Silica gel 60 F254) were purchased from Merck (Germany). GenEluteTM Plasmid Mini Prep Kit was purchased from Sigma-Aldrich. SCL was purified (>95% purity, Fig. 1) from the culture broths of Penicillium sclerotiorum and identified by proton nuclear magnetic resonance as described previously [13].
Media, cultures, and growth conditions
Tryptic soy broth (TSB), nutrient broth (NB), and mueller hinton broth (MBH) were purchased from Becton-Dickinson (USA). S. aureus strains, ATCC 35139
(Methicillin resistant) and ATCC 29213 (methicillin-sensitive) were procured from American Type Culture Collection (USA). SA1199 and SA1199B are clinical strains obtained as gift by Dr. G.W. Kaatz (Wayne State University, USA). All other strain information is pro-vided in the Table 1. The cultures were stored at -80℃ in TSB stocks (50% v/v glycerol). The test bacteria grown either TSB or NB for overnight (12 h) were used for the experiments.
Agar disc diffusion assay
The procedure for evaluation of anti-microbial potency of SCL was performed as per the recommendations by Clinical Laboratory Standards Institute [20]. Briefly, test microorganisms (0.1 ml, 105 colony forming units (cfu)/ml approx.) were inoculated to MH agar plate (20 ml). SCL stock solutions were prepared in sterile DMSO (1 mg SCL in 0.1 ml DMSO) and 20 µl aliquot (100 µg) was loaded to sterile filter paper disk (6-mm dia). The filter paper discs were placed equidistantly on the pre seeded agar. The inoculated plates were incu-bated (37℃, 24 h) and the size of inhibitory zones around the test and control disks was determined.
Minimum inhibitory concentration (MIC)
A twofold serial dilution of SCL (range, 0.5−500 µg) or nisin (250−0.1 µg) were prepared in microtiter plates (MP) containing 0.1 ml of MHB and test bacterial Fig. 1. Sclerotiorin (SCL) structure. The 1NMR spectrum obtained in CDCl3 at 125 MHZ for the TLC pure fraction.The
pro-ton peaks of NMR spectrum were attributable to the groups (a-l) shown in the structure. Pure SCL crystal was shown in the inset.
(106cfu/ml approx.). The plates were incubated sta-tionary 35℃ for 24−48 h. The bacterial growth was determined by measuring the optical density (OD) of cul-ture at 610 nm using a plate reader (Tecan M200 Infinite Pro, Switzerland). MIC is defined as the lowest dilution of the test agent that inhibited >90% of the cell growth.
Time kill kinetics
Viable counts of S. aureus (ATCC 35139 and ATCC 29213) in presence of SCL were determined by time kill kinetic study according to the method of [21] with some modifications. Briefly, MHB (10 ml) adjusted with SCL (31.2, 62.5, and 125 µg/ml) or Nisin (64 µg/ml) and inocu-lated with S. aureus (2 × 105 cfu/ml approx.). An aliquot
of suspension (0.1 ml) was sampled at 0, 3, 6, 9 and 24 h, serial diluted, spread on MHA (0.1 ml), and incubated at 35℃ for 24−48 h. The viable colonies of S. aureus after 48 h were compared between control and test samples. Determination of membrane integrity
The integrity of S. aureus after treatment with SCL was determined using fluorescence assisted cell sorting (FACS) analysis by PI staining and EtBr uptake assays. The samples were prepared according to the method [22] with minor modifications. Briefly, the test bacteria (105 cfu/ml) in two sets of PBS (pH 7.4, 0.1 ml) were incubated with SCL (0−128 µg/ml) at 32℃ for 60 min. One set of test bacteria were incubated with PI (20 µg/ml) for 10 min at 32℃ and the samples were analyzed by flow cytometer (BD FACS Aria III, USA). Further, for real time monitoring of membrane integrity, the second set of SCL treated cell suspension (0.1 ml) was incubated with EtBr (5 µM) in a black bottom MP for 5 min and the EtBr fluorescence was recorded (ex; 360 nm, emi; 616 nm) using MP reader (Tecan Infinite pro, Model 814). Control experiments were conducted without the addi-tion of SCL or with the addiaddi-tion of nisin (128 µg). Determination of membrane potential
The membrane potential of MRSA was determined using Bac LightTM bacterial membrane potential kit (Life Technologies, USA) using 3,3′-diethyloxacarbocy-anine iodide (DioC2 (3)) as per manufacturer’s instruc-tions. Briefly ATCC 35139 cells (OD 0.1) suspended in PBS were labeled with 5 µM of DioC2 (3) at 28℃ for
10 min. The DioC2 (3) labeled cells in PBS were either untreated or treated with SCL (31.2−125 µg/ml) for 5 min. The fluorescence intensity of DioC2 (3) of untreated and SCL treated cells was measured using BD FACS flow cytometer (ex: 488) using argon laser. Nisin (62.5 µg/ml) served as positive control.
Release of UV260 nm absorbing materials
The release of UV260 nm absorbing material by S. aureus in presence of SCL was quantified according to the method previously described [23] with slight modifi-cations. Briefly, S. aureus cells (OD610 nm 0.1) were incubated with SCL (62.5, 125 and 250 µg/ml). At 0, 30, 60, 120, 240 min after incubation, a 0.1 ml aliquot from each treatment was filtered (0.2 µm) and the D of the fil-trate was determined 260 nm. Background OD imparted by SCL at respective concentration was sub-tracted from the final values.
Scanning electron microscopy (SEM)
SEM analysis was carried out according to the method of [24]. Briefly, S. aureus (105 cfu/ml) in PBS was treated
with SCL (128 µg/ml) at 32℃ 60 min. After brief centrif-ugation for 5 min (8000 ×g), the cell pellet was reconsti-tuted with PBS and an aliquot (0.1 ml) was placed on cellulose acetate filter (0.2 µm) and fixed with glutaral-dehyde (4% v/v) for 3 h, rinsed with distilled water, and dehydrated with 50, 70, 80, 90, 95, and 100% (v/v) graded ethanol. The samples were sputter coated with gold and observed under SEM at 10 KV (JOEL, JSM-6390LV, Japan).
Cell penetrating ability of SCL
S. aureus (105 cfu/ml) was incubated with sub MIC of
SCL (15.6 and 31.2 µg/ml) at 32℃ for 2 h. Those without SCL served as negative controls. The bacteria were washed with PBS, harvested by centrifugation (3000 ×g/ 5 min) and the reconstituted pellet in PBS was incu-bated with lysostaphin (10 U) for 10 min. The cell sus-pension was filtered (0.2 µm) and the fluorescence of lysate was determined (ex: 312 nm, emi: 418 nm). DNA binding assay
The interaction of SCL with plasmid DNA was deter-mined by agarose gel electrophoresis. In brief, 200 ng of plasmid DNA isolated from ATCC 35139 using Gen
elute plasmid isolation kit (Sigma-Aldrich) was incu-bated with SCL (0−256 µg/ml) in binding buffer (Tris Hcl, pH 8.4) at 28℃ for 30 min. The DNA migration was analyzed by EtBr staining on 1% agarose by gel electro-phoresis. The degree of DNA condensation was deter-mined using EtBr displacement assay. Plasmid DNA (10 µg/ml) was incubated with EtBr for 1 h at 28℃. The EtBr-DNA complex was incubated with sub MIC of SCL (15.6, 31.2 µg/ml) and the fluorescent intensity of the complex was determined (exi: 520 nm; emi: 590 nm). Interaction of SCL with topical antibiotics
The interaction of SCL with antibiotics was deter-mined by checkerboard method [25]. Twofold serial dilu-tions of antibiotics (bacitracin, polymixin, neomycin) and SCL (0−64 µg/ml) were prepared in 0.1 ml of MBH in MP containing the test bacteria (2 × 105 cfu/ml). The plates were incubated at 37℃ for 24−48 h and OD610 of the cell suspension was determined using MP reader. The frac-tional inhibitory concentration index (FICI) was deter-mined by the following formula:
where FIC A = MIC of SCL in combination with anti-biotic/MIC of SCL alone, FIC B = MIC of antibiotic in combination with SCL/MIC of antibiotic alone). The interaction can be synergy (FICI < 0.5), Indifference (FICI = 4) or antagonism (FICI > 4).
Statistical analysis
A two tailed student’s t test using Graph Pad Prism (GraphPad Software, Version 5.00, USA) was performed.
Results
SCL exhibits anti-staphylococcal activity
It is evident that SCL exhibited narrow spectrum activity with MIC range of 128−1028 µg/ml against Gram positive bacteria including S. aureus (Table 1). SCL exhibited dose dependent inhibitory effect achiev-ing complete suppression (> 90%) of bacterial growth at 125 µg/ml up to 24 h (Fig. S1). Further from Table S1 it is evident that S. aureus strains (including clinical isolates of MRSA) exhibited susceptibility (MIC range, 125−250 µg/ml; MBC range, 125−500 µg/ml) to SCL.
However, Gram-negative and yeast strains were found to be resistant to SCL toxicity. In time kill studies using FICI FIC A
FIC B ---=
Table 1. Anti-bacterial activity of Sclerotiorin against test pathogens.
*Bacterial strain ZOIa
(mm)
MICb (μg/ml) Gram-Positive pathogens
Bacillus subtilis (ATCC 6051) 12 125
Listeria monocytogens (ATCC 19111) 10 1000
Enterobacter clocae (ATCC 13048) 09 1000
Staphylococcus aureus Methicillin-sensitive SA1911 & 1911 B 11 250 ATCC 29213 10 125 Methicillin-resistant ATCC 35139 12 125 01ST001 12 125 Gram-negative bacteria
Escherichia coli (ATCC 8739) 07 ND
Salmonella typhi (KCTC 2515) nil ND
Yeast pathogens
Candida albicans (KCTC 11282) nil ND
Cyrptococcus neoformans(KCCM 50564) nil ND
*ATCC: American Type Culture Collection center, KCTC: Korean
Collection for Type Culture (Daejeon, South Korea), KCCM: Korean Culture of Center of Microorganism (Sudaemun, Seoul). aZOI: Zone
of inhibition, bMIC: minimum inhibitory concentration.
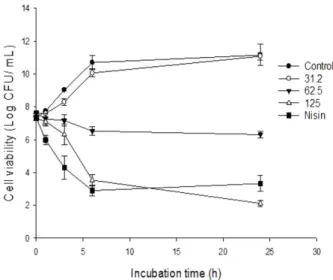
Fig. 2. Time kill kinetic analysis of MRSA (ATCC 35139) in the presence of increasing concentration of sclerotiorin (31.2-125 μg/ml). Mean± SD (n = 4) value was represented.
S. aureus (Fig. 2), the untreated ells exhibited exponen-tial growth reaching >10 log cfu in 24 h. Cells treated with SCL (125 µg/ml) reduced viable counts MRSA by
4 log cfu/ml. Whereas SCL at 64 µg/ml (0.5 ×MIC) caused a 3 log reduction within 6 h but failed to control till 24 h. Similar results were also observed in MSSA Fig. 3. Confocal laser scanning microscopic (CLSM) images of MRSA either untreated (A), treated with DMSO (B), and scle-rotiorin (SCL) at 31.2-125 μg/ml (C-E) or nisin at 62.5 μg/ml (F). S. aureus (ATCC 35139) cells were exposed to SCL or DMSO
(negative control) were subsequently washed with saline, stained with DAPI/PI and subjected to CLSM. Cells with red color indicate dead cells. Each experiment was repeated twice with 5 random filed (Scale: 20 μm).
Fig. 4. Sclerotiorin (SCL) induced cell membrane integrity damage in MRSA. Flowcytometry dot plots (A) and percentage of
MRSA cells (ATCC35139) showing enhanced propidium iodide fluorescence (B) in presence of SCL (32-128 μg/ml). Nisin (128 μg) served as positive control. Mean± SD (n = 4) value was represented. Student’s t test: ***p < 0.001, **p < 0.01.
(data not shown).
SCL causes membrane integrity damage
The confocal laser scanning microscopy (CLSM) study using DAPI (Blue) and PI (red) suggested that untreated cells stained blue, whereas SCL treated cells exhibited red fluorescence (Fig. 3). Further FACS analysis (Fig. 4A, B), showed that only 8.7± 0.3% cells exhibited PI flu-orescence. However, a dose dependant increase in PI
uptake was evident in presence of SCL (64−256 µg/ml) resulting in maximum cell damage by MIC of SCL (p < 0.001, 99.6 ± 0.2%) and this effect was comparable to nisin treatment (p < 0.001, 99.4 ± 0.2%). In time dependent assay, cells treated with SCL (31.2 µg/ml) exhibited PI fluorescence which was not significantly dif-ferent from controls (Fig. 5). However cell treated with SCL (62.5 µg/ml) exhibited increased PI fluorescence (2.3± 1.1 fold) until 120 min. Further increase in SCL (125 µg/ml) rapidly increased PI fluorescence (p < 0.01, 3.1± 1.3 fold) within 60 min and then slightly decreased (Fig. 3B).
Localization of SCL in MRSA
S. aureus cells treated with sub MIC of SCL (15.6−31.2 µg/ml) exhibited green fluorescence (Fig. 6). Further we quantified the intracellular abundance of SCL by mea-suring its fluorescence by fluorescent spectrophotome-ter. From Fig. 6B, it is evident that the fluorescent intensity of cell lysate of S. aureus treated with SCL (15.6−31.2 µg/ml) was significantly high (240 ± 5.2 AU, 1.6 fold, p < 0.05) compared to the control (161± 2.4). SCL induces membrane damage in S. aureus
From flow cytometric dot plots (Fig. 7A), it is evident that SCL (62.5−125 µg/ml) resulted in increased propor-tion of red fluorescence of MRSA cells. Further when Fig. 5. Time dependent uptake of ethidium bromide
facili-tated by sclerotiorin (SCL) in MRSA. EtBr uptake by ATCC
35139 cells in the presence of SCL was quantified by fluores-cent spectrophotometer. Nisin served as positive control). Mean± SD (n = 4) value was represented. Student’s t test: ***p < 0.001, **p < 0.01
Fig. 6. Sclerotiorin (SCL) uptake by individual MRSA cells. Bright field (BF, left), fluorescent (UV, middle) and merge (right) images
of ATCC 35139 (A) either untreated (-SCL) or treated with 31.2 μg/ml SCL (+SCL). Scale bar: 20 μm. Fluorescent intensity of cell lysates of ATCC 35139 cells (B) incubated with SCL (16-32 μg/ml) incubated for 40 min. (n = 6, **p < 0.01).
compared to controls (Fig. 7B) there was a statistically significant reduction (p < 0.01) in the mean fluorescence intensity value of cells incubated with SCL which was comparable to the positive control (Nisin, 62.5 µg). Further from Fig. 8A, it is evident that the UV260
absorbing values of untreated cells slightly increased from 0−240 min. sub MIC of SCL (31.2−62.5 µg/ml) exhibited no considerable increase (0.6 fold) increase in UV260. In presence of SCL (125−250 µg/ml), a rapid increase (p < 0.01, 1.7−3.2 fold) was evident with in Fig. 7. Membrane potential alterations induced by sclerotiorin (SCL) in MRSA. Comparison of flow cytometric profiles of
untreated (control), positive control (nisin), and SCL treated ATCC 35139 cells (A). Mean fluorescence intensity of DioC2 (3) labeled
S. aureus cells either untreated or treated with 31.2-125 μg/ml of SCL (B). (n = 3, **p < 0.01, ***p < 0.001).
Fig. 8. Membrane permeability induced by sclerotiorin (SCL) in MRSA. Leakage of UV260 nm absorbing materials from S. aureus
incubated with SCL (A). Scanning electron microscopy (SEM) analysis of cell morphology of ATCC 35139 cells either untreated (B) or treated with SCL at 128 μg/ml (C)
60 min and remained stable thereafter. In addition, the SEM studies indicated that the untreated S. aureus cells exhibited smooth cell surface (Fig. 8B). In contrast, SCL treated cells exhibited surface blebs with altered mem-brane integrity (Fig. 8C).
SCL interact with MRSA plasmid DNA
At SCL/DNA ratio (0.01 w/w), a fraction of the plasmid DNA is less mobile than plasmid DNA alone. Above SCL/DNA ratio of 0.04, retardation of the plasmid DNA mobility is completely inhibited (Fig. 9A). Additionally, SCL dose dependently (31.2−125 µg/ml) displaced EtBr-DNA complex and resulted in decrease in EtBr fluores-cence in a time dependent manner (Fig. 9B).
SCL potentiates topical antimicrobial efficacy
From Table 2, it is evident that sub MIC (31.2 µg/ml) of SCL exhibited synergy with bacitracin and neomycin (FICI range, 0.26−0.37) and an additive effect with poly-mixin against MRSA and MSSA (FICI = 0.5). However
SCL and antibiotic combinations were ineffective against Gram negative bacteria.
Discussion
SCL was reported to anti-staphylococcal [26]. However the mechanism of anti-staphylococcal action of SCL has not been reported and hence we investigated the anti-staphylococcal mechanism of action of SCL was investi-gated. In the present study, we found that SCL was Gram positive specific especially against S. aureus. The MIC (125−250 µg/ml) of SCL of this study is comparable to the MIC of some azaphilones (MIC range, 15−250 µg/ ml) previously reported against S. aureus [27, 28, 16]. The bactericidal activity exhibited by SCL of the present study (> 4 log cfu reduction) is in contrast with the bacte-riostatic property reported earlier for SCL [27]. In our study SCL exhibited narrow spectrum activity against Gram positive bacteria. The apparent lack of activity of SCL against Gram-negative bacteria may be due to Fig. 9. Interaction of sclerotiorin (SCL) with plasmid DNA. Electrophoretic mobility of plasmid DNA of ATCC 35139 either untreated (Lane 1) or treated with 15.6-128 μg/ml of SCL (A). Displacement of EtBr from the EtBr-plasmid DNA complex by SCL
(B). Mean± SD (n = 4) value was represented.
Table 2. Synergy of Sclerotiorin (SCL) with topical antibiotics on Gram positive and Gram negative bacteria. aStrain
bMIC (μg/ml) MIC (μg/ml) MIC (μg/ml)
SCL(a) PMX(b) (a) + (b) cFICI BAC(c) (a) + (c) FICI NEO(d) (a) + (d) FICI
ATCC35139 125 128 32+128 1.25 256 32+16 0.31 1028 32+64 0.26
ATCC29213 125 256 32+128 0.5 128 32+8 0.31 4 32+0.5 0.37
ATCC8739 >125 256 32+256 1.25 512 32+512 1.25 8 32+4 0.75 KCTC2515 >125 128 32+128 1.25 512 32+512 1.25 2 32+2 1.25
a
ATCC 35139 and 29213 are susceptible for the synergistic combinations of Sclerotiorin (SCL) with bacitracin (BAC) and neomycin (NEO).
bMinimum Inhibitory concentration, cFICI; Fractional Inhibitory Concentration Index. Potential synergistic interaction are indicated in bold
absence of specific target sites for SCL. The lipopolysac-charides of E. coli were reported to prevent the entry of hydrophobic drugs [29].
Our study using CLSM and flow cytometry suggested that SCL could dose dependently alter the cell brane integrity. PI can’t enter the intact cell mem-branes. In the present study SCL facilitated the entry of small molecules such as PI and EtBr by loss of mem-brane integrity mechanisms. This loss in memmem-brane integrity may further result in temporary pore forma-tion in the cell membrane leading to altered membrane potential, membrane damage and release of large mole-cules such as DNA as evident from depolarization, SEM and UV260 nm releasing experiments. SCL exhibits auto fluorescence (exi, 358 nm; emi 512 nm). Using CLSM and fluorescent spectrophotometer, we found that SCL fluorescence was internalized in the cell cytoplasm of S. aureus. In support of our observation, epicocconone with intrinsic fluorescence was reported to be cell permeable and stain the cytoplasmic components [30, 31]. This made us to look for extra cellular targets for SCL and we confirmed the affinity of SCL to bind plasmid DNA and compete for EtBr in EtBr-DNA complex. Sub MIC of SCL (15.6−31.2 µg/ml) also potentiated the efficacy of bacitracin and neomycin and these results in agreement with the synergistic action of SCL (20 µM) with rifampin against mycobacterium [18].
S. aureus acquires multidrug resistance due to thick cell wall, antibiotic modifying enzymes, efflux pumps, and plasmid encoded genes. From our experiments, we deduce certain understanding of mechanism of action of SCL against MRSA. The negative charges of SCL help in preliminary interaction with the cell membrane and establish an electrostatic interaction with positively charged cell components. SCL with its polyene side chain attached to isochromane ring increase the hydro-phobicity of SCL (log P = 1.8) and increase its tendency to partition in to the cell membrane and drives the pene-tration of SCL. An electrochemical gradient know as proton motive force (PMF) across the cell membrane is maintained by the bacteria [32]. Although SCL is known to interact with transmembrane proteins (PknG) via hydrogen interactions [14, 33], our biophysical studies indicated that rapid perturbation of membrane integrity allows slow release of biomolecules resulting in permea-bilization of cell membrane and loss of viability. Our
observations are in line with the earlier findings of [34] that the agents that selectively collapse proton motive force (PMF) can kill bacteria. The intracellular localiza-tion of SCL further can access plasmid DNA and inhibit replication and transcription of plasmid DNA. The potential of membrane targeting compounds to alter PMF, metabolic activity, and intracellular function has been reported [34, 35]. Further, the membrane integrity loss caused by SCL at sub MIC concentration was thought be responsible for neomycin and bacitracin uptake which augmented the toxicity against S. aureus.
The membrane targeting and DNA targeting action of SCL is encouraging. Owing to the health benefits of SCL as antioxidant, anti-inflammatory and antimicrobial, it can be used in topical preparations both as preservative and antimicrobial. However a thorough investigation on the development of structural analogs to and to test its efficacy In-vivo is required inorder to develop SCL in to an effective therapeutic agent.
Acknowledgment
CD would like to thank the Management, Krupanidhi Group of Insti-tutions (KGI) and Krupanidhi-Research Incubation Center (K-RIC, KGI) for constant support and encouragement.
Conflict of Interest
The authors have no financial conflicts of interest to declare.
References
1. Stewart CM, Cole MB, Legan JD, Slade L, Vandeven MH, Schaffner DW. 2002. Staphylococcus aureus growth boundaries: moving mechanistic predictive mode is based on solute-specific effects.
Appl. Environ. Microbiol. 68: 1864-1871.
2. Tommasi R, Brown DG, Walkup GK, Manchester JI, Miller AA. 2015. ESKAPEing the labyrinth of antibacterial discovery. Nat.
Rev. Drug. Disc. 14: 529-542.
3. Friedman M. 2015. Antibiotic-resistant bacteria: prevalence in food and inactivation by food compatible compounds and plant extracts. J. Agric. Food. Chem. 63: 3805-3822.
4. Osmanova N, Schultz W, Ayoub N. 2010. Azaphilones a class of fungal metabolites with diverse biological activities. Phytochem.
Rev. 9: 315-342.
5. Dong J, Zhou Y, Li R, Zhou W, Li L, Zhu Y, et al. 2006. New nemati-cidal azaphilones from the aquatic fungus Pseudohalonectria
adversaria YMF1.01019. FEMS. Microbiol. Lett. 264: 65-69.
azaphilones, leteusins C, D, and E, from an ascomycete, Talaromyces
luteus. Chem. Phar. Bull. 44: 284-287.
7. Yang DJ, Tomoda H, Tabata N, Masuma R, Omura S. 1996. New isochromophilones VII and VIII produced by Penicillium sp. FO-4164. J. Antibiot. 49: 223-229.
8. Yu BZ, Zhang GH, Du ZZ, Zheng YT, Xu JC, Luo XD. 2008 Pho-moeuphorbins A-D, azaphilones from the fungus Phomopsis
euphorbiae. Phytochemistry 69: 2523-2526.
9. Dos Santos P, Ferraz C, Ribeiro PP, Miranda FM, da Silva F, de Souza JT, et al. 2019. Antioxidant and antibacterial activities of the chlorine pigment sclerotiorin from Penicillium mallochii and its chemotaxonomic significance. Biochem. Syst. Ecol. 86: 103915. 10. Musso L, Dallavalle S, Merlini L, Bava A, Nasini G, Penco S, et al. 2010. Natural and semisynthetic azaphilones as a new scaffold for Hsp90 inhibitors. Bioorg. Med. Chem. 18: 6031-6043.
11. Nam JY, Kim HK, Kwon JY, Han MY, Son KH, Lee UC, et al. 2000. 8-O-Methylsclerotiorinamine, antagonist of the Grb2-SH2 domain, isolated from Penicillium multicolor. J. Nat. Prod. 63: 1303-1305. 12. Pairet L, Wrigley SK, Chetland I, Reynolds EE, Hayes MA, Holloway
J, et al. 1995. Azaphilones with endothelin receptor binding activity produced by Penicillium sclerotiorum: taxonomy, fermen-tation, isolation, structure elucidation and biological activity. J.
Antibiot. 48: 913-923.
13. Chidananda C, Jagan Mohan Rao L, Sattur AP. 2006. Sclerotiorin, from Penicillium frequantans, a potent inhibitor of aldose reduc-tase. Biotechnol. Lett. 28: 1633-1636.
14. Tomoda H, Matsushima C, Tabata N, Namatame I, Tanaka H, Bam-berger MJ, et al. 1999. Structure-specific inhibition of cholesteryl ester transfer protein by azaphilones. J. Antibiot. 52: 160-170. 15. Giridharan P, Verekar SA, Khanna A, Mishra PD, Deshmukh SK.
2012. Anticancer activity of sclerotiorin, isolated from endo-phytic fungus cephalotheca faveolata Yaguchi, Nishim, & Udagawa. Ind. J. Exp. Biol. 50: 464-468.
16. Arunpanichlert J, Rukachaisirikul V, Sukpondma Y, Phongpaichit S, Tewtrakul S, Rungjindamai S, et al. 2010. Azaphilone and iso-coumarin derivatives from the endophytic fungus Penicillium
Sclerotiorum PSU A-13. Chem. Pharm. Bull. 58: 1033-1036.
17. Lin L, Mulholland N, Huang SW, Beattie D, Irwin D, Gu YC, et al. 2012. Design, synthesis, and fungicidal activity of novel scleroti-orin derivatives. Chem. Biol. Drug. Des. 80: 682-692.
18. Chen D, Ma SS, He L, Yuan P, She Z, Lu Y. 2017. Sclerotiorin inhibits protein Kinase G from Mycobacterium tuberculosis and impairs mycobacterial growth in macrophages. Tuberculosis 103: 37-43. 19. Gomes DC, Takahashi AP. 2016. Sequential fungal fermentation
biotransformation process to produce a red pigment from Scle-rotiorin. Food Chem. 210: 355-361.
20. Clinical and Laboratory Standards Institute. 2016. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically; approved standards. CLSI document M45, 3rd Ed.
Clinical and Laboratory and Standards Institute, Wayne, PA. 21. Pag U, Oedenkoven M, Papo N, Oren Z, Shai Y, Sahl HG. 2004. In
vitro activity and mode of action of diasteriometic antimicrobial
peptides against bacterial clinical isolates. J. Antimicrob. Chemother.
53: 230-239.
22. Uppu DSSM, Manjunath GB, Yarlagadda V, Kaviyil JE, Ravikumar R, Paramanandham K, et al. 2015. Membrane active macromole-cules resensitize NDM-1 gram negative clinical isolates to tetra-cycline antibiotics. PLoS One 10: e0119422.
23. Carson CF, Mee BJ, Rile TV. 2002. Mechanism of action of Melaleuca
alternifolia (Tea tree oil) on Staphylococcus aureus determined by
time kill assay, lysis, leakage, and salt tolerance assays, and elec-tron microscopy. Antimicrob. Agents. Chemother. 46: 1914-1920. 24. Tyagi P, Singh M, Kumari H, Kumari A, Mukhopadhyay K. 2015.
Bactericidal activity of curcumin I is associated with damaging of bacterial membrane. PLoS One 10: e0121313.
25. Nair DR, Monteiro JM, Memmi G, Thanassi J, Pucci M, Schwartz-man J, et al. 2015. Characterization of a novel small molecule that potentiates β-lactam activity against positive and Gram-negative pathogens. Antimicrob. Agents. Chemother. 59: 1876-1885.
26. Zhou SL, Wang M, Zhao HG, Huang YH, Lin YY, Tan GH, et al. 2016. Penicilazophilone C, a new antineoplastic and antibacterial azaphilone from the marine fungus Penicillium Sclerotiorum.
Arch. Pharm Res. 39: 1621-1627.
27. Lucas EMF, Monteiro de Castro MC, Takahashi JA. 2007. Antimi-crobial properties of Sclerotiorin, isochromophilone VI, and pen-colide, metabolites, from a brazilian cerrado isolate of Penicillium
sclerotiorum Van Beyma. Braz. J. Microbiol. 38: 785-789.
28. Gao SS, Li XM, Zhang Y, Li CS, Cui CM, Wang BG. 2011. Coma-zaphilones A-F, azaphilone derivatives from the marine sedi-ment-derived fungus Penicillium commune QSD 17. J. Nat. Prod.
74: 256-261.
29. King JD, Kocincova D, Westman EL, Lam JS. 2009. Review: lipo-polysaccharide biosynthesis in Pseudomonas aeruginosa. Innate
Immun. 15: 261-312.
30. Bell PJL, Karuso P. 2003. Epicocconone, a novel fluorescent com-pound from the fungus Epicoccum nigrum. J. Am. Chem. Soc. 25: 9304-9305.
31. Choi HY, Veal DA, Karuso P. 2006. Epicocconone, a new cell-permeable long stoke’s shift fluorescent stain for live cell imag-ing and multipleximag-ing. J. Fluoresc. 16: 475-482.
32. Mitchell P. 1966. Chemiosmotic coupling in oxidative and photo-synthetic phosphorylation. Biol. Rev. Camb. Philos. Soc. 41: 445-502. 33. Cheng MJ, Wu MD, Yanai H, Su YS, Chen IS, Yuan GF, et al. 2012.
Secondary metabolites from the endophytic fungus
Biscogni-auxia formosana and their antimycobacterial activity. Phytochem. Lett. 5: 467-472.
34. Farha MA, Verschoor CP, Bowdish D, Brown ED. 2013. Collapsing the proton motive force to identify synergistic combinations against Staphylococcus aureus. Cell. Chem. Biol. 20: 1168-1178. 35. del Castillo FJ, Del Castillo I, Moreno F. 2001. Construction and
characterization of mutations at codon 751 of the Escherichia coli gyrB gene that confer resistance to the antimicrobial peptide microcin B17 and alter the activity of DNA gyrase. J. Bacteriol.