대한소화기학회지 2000;36:441 - 449
접수: 2000년 6월 8일, 승인: 2000년 8월 29일
연락처: 박유환, 501-759, 광주광역시 동구 서석동 375번지, 조선대학교 의과대학 내과학교실 Tel: (062) 220-3671, Fax: (062) 232-4943
※ 이 논문은 1999년도 조선대학교 학술연구비의 지원을 받아 연구되었음.
위 평활근에서 이온전류들에 대한 Im ipr a m in e 효과
조선대학교 의과대학 생리학교실, 정신과학교실*, 내과학교실†, 국립나주정신병원 , 서울대학교 의과대학 생리학교실§
전제열・염철호・김상훈*・박찬국†・정춘해†・박유환†・이병희 ・김기환§
E ffe c t s o f Im i p r a m i n e o n Io n i c Cu rr e n t s i n G a s t r i c S m o o t h Mu s c le o f G u i n e a -p i g
J a e Ye o u l J u n , M .D., Ch e o l H o Ye u m , P h .D ., S a n g H o o n Ki m , M .D .*, Ch an Gu k P ark, M.D.†, Ch oon Hae Ch u n g, M.D.†, Yoo Wh an P ark, M.D.†,
B y a n g H ae Le e , M.D. a nd Ki Wh a n Ki m , M.D.§
Departments of Physiology, Psychiatry* and Internal Medicine†, College of Medicine, Chosun University, Kwangju; Naju National Mental Hospital , Department of Physiology and Biophysics§,
Seoul National University College of Medicine, Seoul, Korea
Background/Aims: Imipramine is a widely used tricyclic antidepressant, but it has side effects of
gastrointestinal motility frequently. Gastrointestinal motility is related with ionic currents of smooth muscle cells. The aim of this study was to investigate whether or not imipramine could modulate ionic currents in gastric smooth muscle cells. Methods: Guinea-pig gastric smooth muscle cells were isolated from the antral circular layer using a collagenase, and then we recorded their membrane ionic currents using a whole-cell patch clamp technique. Results: Imipramine suppressed delayed rectifier K+ currents in a whole test voltage range. Chelerythrine, a protein kinase C inhibitor, did not affect on the inhibitory effect of imipramine on the delayed rectifier K+ currents. Under high extracellular K+ (90 mM), lemakalim produced inward currents at a -80 mV of holding potential, which were blocked by glibenclamide, a specific blocker of ATP-sensitive K+ channels. Imipramine inhibited the lemakalim-induced, ATP-sensitive K+ currents. Imipramine also inhibited the voltage- dependent Ca2+ currents in a concentration-dependent way in a whole test voltage range.Conclusions: These results suggest that imipramine could alter various membrane ionic currents in
gastric smooth muscle cells, which may cause side effects on gastrointestinal motility. (Kor JGastroenterol 2000;36:441 - 449)
Key Words: Imipramine, Delayed rectifier K+ currents, ATP-sensitive K+ currents, Voltage-
dependent Ca2+ currents, Gastric smooth muscle cells442 대한소화기학회지 : 제 36 권 제 4 호 2000
서 론
Imipramine은 우울증과 정동장애에 널리 쓰여지 고 있는 삼환고리계의 항우울제 약물로서 항무스카 린성 효과, 직접성 근 이완, Ca2+ 길항작용 및 신경 말단에서 amine의 재흡수를 억제하는 작용 등 다양 한 약리학적 효과를 가지고 있다.1 Imipramine의 therapeutic 혈중농도는 0.8에서 6.3 μM/L이며 복용 시 오심, 구토와 설사 등 상대적으로 빈번한 위장관 운동성 부작용을 야기시킨다.2 무스카린 작용의 변 이나 신경말단에서 amine의 재흡수를 억제하여 장 관신경계의 기능이 변이되는 경우 위장관운동성의 변화가 초래될 수 있으며 결국 위장관운동성에 대 한 부작용을 나타낼 수 있다.3 그러나 혈관4이나 방 광 평활근,5 보고된 바와 같이 imipramine 또한 직접 적으로 위장관 평활근에 작용할 수 있다. Imi- pramine은 심장의 부정맥 치료제로도 분류되어 있 으며6 irritable bowel syndrome에서 보이는 운동성 장애에 치료제로도 제시되고 있다.7 심장의 부정맥 이나 위장관운동성은 세포막의 이온 전도도의 변화 와 관련이 있다. 세포막에 존재하는 이온통로들은 평활근의 막전압과 흥분성을 조절하고 있다. 내향성 이온전류의 활성화나 외향성 이온전류의 억제는 막 전압을 저분극(depolarization)시켜 세포의 흥분성을 증가시키는 반면, 내향성 이온전류의 억제 또는 외 향성 이온전류의 활성화는 막전압을 과분극(hyper- polarization)시켜 반대로 세포의 흥분성을 감소시킨 다. Imipramine은 신경8-12과 심장근13에서 막전압-의 존성 Na+ 통로, Ca2+ 및 K+통로를 조절한다고 알려 져 있다. 그러나 위장관 평활근의 이온전류들에 대 한 imipramine의 효과와 조절에 대해서는 잘 알려져 있지 않다. 따라서 본 연구는 imipramine이 위 평활 근에 존재하는 Ca2+통로와 K+통로에 대한 조절 여 부를 알아보기 위하여 단일세포에서 세포막전압고 정법(whole-cell patch clamp technique)을 이용하여 이온전류를 기록하였다.
대상 및 방법
1. 세포 분리
체중 300g 내외의 기니피그를 암수 구별 없이 실 험동물로 사용하였다. 경동맥을 절단하여 실혈시킨 뒤 개복하여 위를 적출하였다. 실온에서 100% O2로 평형을 이루고 있는 phosphate-완충 Tyrode 용액 (NaCl 147, KCl 4, CaCl2 2, MgCl2 1.05, NaH2PO4 0.42, Na2HPO3 1.81, glucose 5.5 mM, pH 7.35)으로 채워진 준비용기 속에서 유문동 부위를 떼어낸 후, 소만부위를 따라 위를 절개하였다. 현미경 밑에서 점막층을 박리하여 윤상근이 표면에 노출되게끔 깨 끗이 분리하였다. 윤상근만을 종주근으로부터 조심 스럽게 분리하여 작은 절편을 만들었다(2×3 mm).
절편들을 0.1% collagenase, 0.1% trypsin inhibitor, 0.2% bovine serum albumin이 들어있는 Ca2+이 없는 생리식염수(Physiological Salt Solution, PSS)에 옮긴 다음 37℃에서 20-25분간 항온 소화시켰다. 소화시 킨 후 low Cl-, high K+ 용액14으로 교체한 다음 끝이 무딘 유리피펫을 사용하여 조심스럽게 진탕시켜 단 일 세포를 분리하고 4℃에 냉장 보관하였다. 모든 실험은 단일세포를 분리한 후 30분 후에 시행하였 으며 실온에서 시행하였다.
2. 막전류의 기록
분리된 세포들을 도립현미경(inverted microscope) 위에 설치되어 있는 작은 실험용기(300 μl)로 옮긴 후 분당 2-3 ml 속도로 세포 외 용액을 관류시켰다.
유리전극의 저항은 3-5 MΩ의 것을 사용하였다. 막 전압고정법은 Hamil 등15과 유사한 방법을 사용하였 다. Patch clamp 증폭기(Axopatch 1-D, Axon Ins- truments, USA)를 통하여 나오는 신호는 digital oscilloscope (Hitachi, Japan), 생리기록기(Recorder 220, Gould, USA)를 통해서 관찰하였고, 고정전압 과 자극전압의 조정 및 전류의 기록은 pClamp software v.6.0 (Axon Instruments, USA)와 IBM- compatible computer를 사용하였다.
전제열 외 7인. 위 평활근에서 이온 전류들에 대한 Imipramine의 효과 443
3. 실험용액
세포 외 관류용액은 생리적 식염수(NaCl 134.8, KCl 6.2, CaCl2 2.3, glucose 12.2, HEPES 0.4, tris 1.8 mM, pH 7.4)를 사용하였다. Ca2+ 전류만을 기록 할 경우 전극 내 용액을 cesium-aspartate 110, Mg- ATP 5, di-tris creatinine phosphate 2.5, disodium creatinine phosphate 2.5, MgCl2 1, HEPES 0.1, TEA-Cl (tetraethylammonium chloride) 20, EGTA (ethyleneglycol-bis-(β-aminoethylether) N,N,N' ,N' - tetra-acetic acid) 5 mM, pH 7.4인 용액을 사용하였 으며, 외향성 K+ 전류를 관찰하기 위해서 전극 내 K-aspartate 110, Mg-ATP 5, di-tris creatine phosphate 5, KCl 20, MgCl2 1, EGTA 0.1 (or 10), HEPES 5 mM pH 7.4인 용액을 사용하였다. Lemakalim으로 활성화되는 전류를 기록하기 위해서 세포 내 전극 은 NaCl 10, KCl 102, CaCl2 1, GTP 1, HEPES 10, EGTA 10, ATP 0.1, MgCl2 1, KOH 38 mM, pH 7.2 인 용액을 사용하였다.
본 연구는 실험 성적 분석이 집단 간의 정성적 비교이기 때문에 4-13번 반복 실험하여 같은 경향을 확인한 후 그 중 대표적인 결과를 본 논문에 게재하 였다.
4. 실험에 사용된 약물
Imipramine (Sigma), glibenclamide (Sigma), ni- fedipine (Sigma), TEA (Sigma), lemakalim (SKB), chelerythrine (RBI).
결 과
1. De la y ed r ec t if ier K + cu r r e n t s 에 대한 im ipr a m in e의 효과
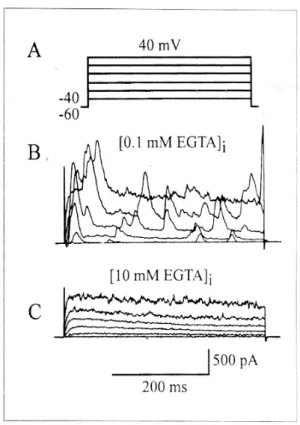
Fig. 1은 위 유문동 윤상근세포에서 기록한 외향 성 K+ 전류를 보여주고 있다. 유지전압을 -60 mV에 서 고정한 상태에서 저분극 자극을 주어 전류를 기 록하였다. 전극 내 Ca2+ 완충제인 0.1 mM EGTA가 포함되어 있는 용액을 사용하여 20 mV씩 증가시키 면서 40 mV까지 단계적으로 400 ms 동안의 저분극 자극(Fig. 1A)을 주게 되면 일과성 및 자발적 진동
을 보이는 K+ 전류(transient and spontaneous oscillatory current)가 기록된다(Fig. 1B)(n=4). 이러 한 외향성 K+ 전류는 전극 내 10 mM EGTA를 첨가 하여 기록하면 일과성 및 자발적 진동을 보이는 K+
전류는 사라지고 지속적인 외향성 K+ 전류(delayed rectifier K+ current)만이 기록된다(Fig. 1C). 이는 일 과성 및 자발적 진동을 보이는 K+ 전류가 세포 내 Ca2+에 의존성임을 나타내고 있다. Fig. 2A는 delayed rectifier K+ 전류에 대한 TEA 효과를 나타낸 것이 다. TEA는 농도에 의존해서 delayed rectifier K+ 전 류를 억제하였다. TEA 2 mM과 10 mM 투여 전과 투여 후 각 전압에 대한 최대전류의 크기를 전류-전 압곡선에 표시하였다. 10 mM TEA 투여시 20 mV 에서 대조군의 전류 크기의 76.8% (n=5)까지 억제
Fig. 1. Whole-cell outward K+ currents recorded in gastric smooth muscle cells. A, Pulse protocol. B, Transient and spontaneous oscillatory K+ currents recorded with 0.1 mM of EGTA in the pipette. C, These transient and spontaneous oscillatory K+ currents were abolished at 10 mM of EGTA in the pipette and only delayed rectifier K+
currents were recorded.
444 The Korean Journal of Gastroenterology : Vol. 36, No. 4, 2000
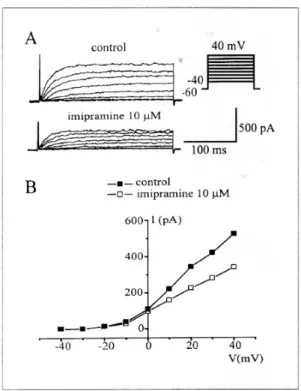
되었다(Fig. 2B). Fig. 3는 delayed rectifier K+ 전류 에 대한 imipramine의 효과와 전류-전압곡선을 나타 내고 있다. 세포외액에 imipramine (10 μM) 투여시 20 mV에서 대조군 전류 크기의 63.7% (n=6)까지 억제되었다. 여러 세포에서 protein kinase C가 delayed rectifier K+전류를 조절한다고 알려져 있 다.16,17 따라서 imipramine의 억제 효과가 protein kinase C를 매개하여 이루어지는가를 보기 위하여 protein kinase C의 선택적 억제제인 chelerythrine을 전처치하여 imipramine의 억제 효과를 관찰하였다.
Imipramine의 효과를 관찰하기 전 chelerythrine을 20분 동안 전처치하였다. Chelerythrine (1 μM) 그 자체는 K+ 전류의 크기에는 아무런 영향을 보이지 않았다(data not shown). Imipramine은 chelerythrine 의 존재하에서도 여전히 delayed rectifier K+ 전류를 억제하였다(Fig. 4A). Chelerythrine 존재하에서 20 mV에서 기록된 imipramine의 억제 효과는 대조군 전류 크기의 68.4±6% (n=4)까지 억제되어 chelery- thrine이 존재하지 않는 상태에서 기록된 값과 통계 학상 유의한 차이를 보이지 않았다(Fig. 4B).
2. A T P - 민감성 K + 전류에 대한 im ipr a - m in e 의 효과
Delayed rectifier K+ 전류와 Ca2+-의존성 K+ 전류 를 배제하기 위하여 유지전압은 -80 mV로 고정하 고 전극 내 10 mM EGTA를 사용하여 세포 내 Ca2+
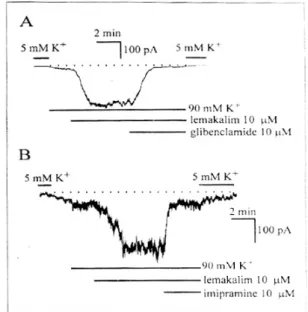
을 완충하였다. 세포 외 90 mM K+ 용액으로 관류 시켜 K+ 전류 전도도를 증가시킴과 아울러 K+의 평 형전압 값 아래에서 내향성 방향으로 전류가 기록 되게 하였다. 세포 외 5 mM K+ 용액에서 90 mM K+ 용액으로 관류하게 되면 내향성의 전류가 발생 된다. 이 결과에서 lemakalim (10 μM)을 투여하게 되면 더 큰 내향성 전류(-224±34 pA, n=13)가 기록 되었다. 내향성 전류는 ATP-민감성 K+ 통로를 선택 적으로 차단하는 glibenclamide (10 μM)을 투여한 경우 차단되었다(Fig. 5A). Imipramine (10 μM)은 lemakalim으로 활성화된 전류를 95±3.4% (n=5)까 지 억제하였다(Fig. 5B).
Fig. 2. Effects of TEA on the delayed rectifier K+ currents recorded with 10 mM EGTA in the pipette. A, TEA (2 and 10 mM) suppressed delayed rectifier K+ currents in a dose-dependent manner. B, The current-voltage relation- ships are compared between to the peak currents in control and in the presence of TEA. n=5.
Fig. 3. Effects of imipramine on the delayed rectifier K+ currents. A, Imipramine (10 μM) inhibited delayed rectifier K+ currents in a whole test voltage range. B, The current-voltage relationships are compared between to the peak currents in control and in the presence of imipramine. n=6.
Jun, et al. Effects of Imipramine on Ionic Currents in Gastric Smooth Muscle of Guinea-pig 445
3. 막전압- 의존성 Ca 2+ 전류에 대한 im i- pr a m in e 의 효과
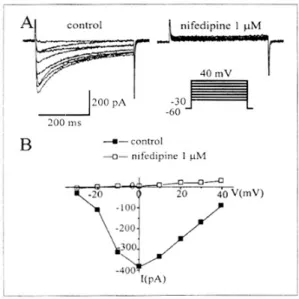
K+ 전류를 차단하는 물질인 cesium과 TEA가 들 어 있는 전극 내 용액을 사용하여 이온전류를 기록 하였다. 유지전압 -60 mV에서 단계적으로 10 mV씩 저분극 자극을 주게 되면 -30 mV에서부터 활성화 되어 0 mV에서 최대크기를 보였다가 그 이후 전압 에서 다시 크기가 감소하는 막전압-의존성 내향 전 류가 기록되었다. 이러한 전류는 Ca2+ 통로 차단제 인 nifedipine 10 μM에 의해서 완전히 차단되며(Fig.
6)(n=4), 세포 외 Ca2+을 제거하면 내향성 전류가 사 라지게 되어 다른 평활근세포에서 보고한 막전압-
의존성 Ca2+ 전류임을 알 수 있었다. Fig. 7은 Ca2+
전류에 대한 imipramine의 효과와 전류-전압곡선을 나타내고 있다. Imipramine은 농도-의존적으로 막전 압 의존성 Ca2+ 전류를 차단하였다(n=4).
고 찰
위장관 평활근의 자발적 수축은 막전압이 주기적 으로 변하는 전기적 현상인 서파에 의해 발생되며18 세포막 전압은 세포막에 존재하는 이온통로들의 활 동도에 의해서 결정된다. 평활근세포에서 특정이온 의 활성화는 막전압의 저분극과 과분극을 유발하고, 이는 평활근 수축과 이완에 매우 중요한 역할을 하 고 있다. K+ 통로의 활성화는 막전압을 과분극시켜 세포의 흥분성을 저하시킨다. 위 평활근에 존재하는 K+ 통로는 주로 delayed rectifier K+ 통로, Ca2+-의 존성 K+ 통로19-21와 ATP-민감성 K+ 통로이다.22,23 Fig. 4. Effect of chelerythrine, a selective protein kinase
C inhibitor, on the inhibitory action of imipramine on the delayed rectifier K+ currents. A, Currents were recorded in control and in the presence of imipramine (10 μM) and chelerythrine (1 μM). Chelerythrine did not affect on the inhibitory effect of imipramine. B, The current-voltage relationships are compared between to the peak currents in control and in the presence of imipramine and chelery- thrine. n=4.
Fig. 5. A, Typical trace illustrating the effect of glibenclamide (10 μM) on the lemakalim-activated K+
current. The current was recorded at -80 mV of holding potential with 0.1 mM ATP and 140 mM K+ in the pipette and 90 mM K+ in the bath solution. Dotted line indicates the zero current level. B, Effects of imipramine on the lemakalim-activated K+ current. Imipramine (10 μM) inhibited lemakalim-activated K+ current (n=5). Dotted line indicates the zero current level. n=5.
446 대한소화기학회지 : 제 36 권 제 4 호 2000
본 실험에서 imipramine은 TEA에 억제되는 delayed rectifier K+ 전류를 억제하였다. 이 전류는 여러 세
포에서 이차전령계인 protein kinase C에 의해서 조 절받고 있다고 알려져 있다.16,17 Ferrugia 등24은 또 다른 항우울제인 fluoxetine이 개의 jejunal 평활근세 포에서 protein kinase C를 통하여 delayed rectifier K+ 전류를 억제한다고 보고하였다. 그러나 본 실험 에서는 protein kinase C 억제제를 전처치한 후에도 여전히 imipramine은 delayed rectifier K+ 전류를 억 제하여 protein kinase C와는 무관한 작용 양상을 나 타냈다. Ferrugia 등과의 실험 결과의 차이는 실험에 사용한 동물이나 조직의 차이로 인한 결과일 것으 로 생각된다. 왜냐하면 동물이나 조직에 따라 위장 관의 전기적 활동도인 서파의 빈도나 크기가 다르 기 때문에 여기에 관여하는 이온전류의 양상이나 조절 등이 다르리라 생각된다. 신경9,11이나 심장근 6,13에서도 imipramine이 protein kinase C와는 무관 하게 delayed rectifier K+ 전류를 억제한다고 알려져 있다.
ATP-민감성 K+ 통로는 처음 심장근25에서 보고된 이후 췌장의 β-세포,26 혈관 평활근27,28 담낭 평활근 29 방광 평활근30에서도 존재하는 것으로 알려져 있 다. 특히 대장 평활근에서는 기초 상태에서도 활성 화되어 안정막 전압을 유지하는 데 대단히 중요한 역할을 담당하고 있다.31 이 통로의 특징은 세포 내 ATP 농도가 낮아지면 활성화되는 것으로 통로를 차단하면 막전압이 저분극되어 세포의 흥분성이 증 가된다. 이 통로는 K+ 통로 개방제인 lemakalim, cromakalim, pinacidil 그리고 diazoxide 등에 의해서 활성화되며 sulfonylurea 계통의 glibenclamide나 tolbutamide 등에 의해서 선택적으로 차단된다.32 본 연구자 등도22,23 이미 기니피그 위 평활근세포에서 이 통로가 존재한다는 것을 보고하였다. 본 실험에 서 imipramine은 ATP-민감성 K+ 전류를 억제하였 다. Imipramine이 통로를 cloning하여 Xenopus oocyte에 발현시킨 후 실험한 결과에서 ATP-민감성 K+ 전류를 직접 억제한다고 보고하고 있다.33 이러 한 결과는 imipramine이 이 통로를 억제하여 막전압 을 변화시킬 수 있음을 암시한다.
평활근 수축에 있어 일정한 농도의 세포 내 Ca2+
의 증가는 필수적이다. 세포 내 Ca2+의 동원은 크게 두 가지로 세포 외에서 막전압-의존성 Ca2+ 통로를 Fig. 6. Voltage-dependent Ca2+ currents in gastric smooth
muscle cells. A, Inward control currents at various test potentials from a -60 mV of holding potential by incre- ment of 10 mV steps. Inward currents were completely blocked in the application of nifedipine (1 μM). B, The current-voltage relationships are compared between to the peak currents in control and in the presence of nifedipine.
n=4
Fig. 7. Effects of imipramine on the voltage-dependent Ca2+ currents. A, Imipramine (10 μand 100 μM) sup- pressed Ca2+ currents in a dose-dependent manner. B, The current-voltage relationships are compared between to the peak currents in control and in the presence of imi- pramine. n=4.
전제열 외 7인. 위 평활근에서 이온 전류들에 대한 Imipramine의 효과 447
통해서 들어오는 것이고 또 하나는 세포 내 Ca2+ 저 장고인 endoplasmic reticulum에서 분비되는 것이 다.34 혈관 평활근에서는 2가지 종류의 막전압-의존 성 Ca2+ 통로가 존재한다고 알려져 있으나35 위 평 활근세포에서는 dihydropyridine 계통의 약물인 nifedipine에 의해서 차단되는 L-type의 Ca2+ 통로만 이 존재하는 것으로 알려져 있다.19,20 본 실험에서 imipramine은 농도-의존적으로 역치전압 및 최대전 류를 나타내는 막전압에는 아무런 변화 없이 시험 전압 전 범위에서 Ca2+ 전류를 억제하였다. 이상의 결과들은 imipramine이 여러 종류의 이온통로들을 조절하여 위 평활근의 수축성을 변화시킬 수 있음 을 시사한다.
결론적으로 imipramine은 위 평활근세포에서 delayed rectifier K+ 전류, ATP-민감성 K+ 전류 및 막전압-의존성 Ca2+ 전류를 억제하였다. 따라서 이 러한 작용이 위장관운동성에 부작용을 일으키는 기 전이 될 수 있으리라 생각된다.
요 약
목적: Imipramine은 널리 쓰이는 항우울제이지만 상대적으로 빈번한 위장관운동성 부작용을 초래한 다. 위장관운동성은 평활근세포의 이온전류와 관계 하는 바 본 실험은 imipramine의 위 평활근세포에 존재하는 여러 이온전류들에 대한 작용을 알아보고 자 하였다. 대상 및 방법: 기니피그 위 윤상근 조직 에서 collagenase를 사용하여 단일세포를 분리한 후 세포막전압고정법(whole cell patch clamp technique) 을 시행하였다. Delayed rectifier K+ 전류를 기록하 기 위하여 K+-aspartate용액을 사용하였고, ATP-민 감성 K+ 전류를 기록하기 위하여 유지전압을 -80 mV에 고정하였으며 세포 외 용액은 90 mM K+ 용 액을 사용하였다. 막전압-의존성 Ca2+ 전류를 기록 하기 위하여 전극 내 20 mM TEA가 들어있는 용액 을 사용하였다. 결과: Imipramine은 시험전압 전 범 위에서 delayed rectifier K+ 전류를 억제하였다.
Protein kinase C 억제제인 chelerythrine은 imipra- mine의 억제작용을 차단하지 못하였다. 유지전압 -80 mV에서 K+-통로 개방제인 lemakalim은 내향성
전류를 활성화시켰으며 ATP-민감성 K+ 통로 차단 제인 glibenclamide에 의해서 억제되었다. Imipra- mine은 ATP-민감성 K+ 전류를 억제하였다. Imipra- mine은 또한 막전압-의존성 Ca2+ 전류를 농도-의존 적으로 시험전압 전 범위에서 억제하였다. 결론: 이 상의 실험 결과들은 imipramine이 위 평활근세포에 서 여러 이온통로들을 조절할 수 있음을 제시하며 이러한 작용들이 위장관운동성에 부작용을 일으킬 수 있는 기전이 될 수 있으리라 생각된다.
색인단어: Imipramine, Delayed rectifier K+ 전류, ATP-민감성 K+ 전류, 막전압-의존성 Ca2+ 전류, 위 평활근세포
참 고 문 헌
1. Grover R, Ruggieri MR, Levin RM. Functional effects of imipramine on the rabbit urinary bladder:
an in-vitro study. Pharmacology 1988;37:148-153.
2. Gilman AG, Rall TW, Nies AS, Taylor P. The pharmacological basis of therapeutics. 8th ed. New York: Pergamon Press, 1991.
3. 이상인. 위의 운동성질환. 대한소화기학회지 1991;
23:325-330
4. Auguet M, Clostre F, Defeudies FV. Effects of antidepressants on receptor-activated and Ca2+- activated contractions of rabbit isolated aorta. Gen Pharmacol 1986;17:607-610.
5. Pietra C, Poggesi E, Angelico P, Guarneri L, Testa R. Effects of some antidepressants on the volume- induced reflex contractions of the rat urinary bladder: lack of correlation with muscarinic recep- tors affinity. Pharmacol Res 1990;22:421-432.
6. Valenzuela C, Chapula J, Delpon, E, Elizalde A, Perez O, Tamarago J. Imipramine blocks rapidly activating and delays slowly activating K+ current activation in guinea pig ventricular myocytes. Circ Res 1994;74:687-699.
7. Gorard DA, Libby GW, Farthing MJ. Effect of tricyclic antidepressant on small intestinal motility in helath and diarrhea-predominant irritable bowel
448 The Korean Journal of Gastroenterology : Vol. 36, No. 4, 2000
syndrome. Dig Dis Sci 1995;40:86-95.
8. Pancrazio JJ, Kamatchi, GL, Roscoe AK, Lynch C.
Inhibition of neuronal Na+ channels by antide- pressant drugs. J Pharmacol Exp Ther 1998;284:
208-214.
9. Ogata N, Yoshii M, Narahashi T. Psychotic drugs block voltage-gated ion channels in neuroblastoma cells. Brain Res 1989;476:140-144.
10. Choi JJ, Huang GJ, Shafik E, Wu WH, Mcardle JJ.
Imipramines selective suppression of an L-type calcium channel in neurons of murine dorsal root ganglia involves G proteins. J Pharmacol Exp Ther 1992;263:49-53.
11. Wooltorton JRA, Mathie A. Block of potassium currents in rat isolated sympathetic neurons by tricyclic antidepressants and structurally related compounds. Br J Pharmacol 1993;110:1126-1132.
12. Lee K, Mckenna F, Rowe IC, Ashford ML. The effects of neuroleptic and tricyclic compounds on BKCa channel activity in rat isolated cortical neurons. Br J Pharmacol 1997;121:1810-1816.
13. Isenberg G, Tamargo J. Effect of imipramine on calcium and potassium currents in isolated bovine ventricular myocytes. Eur J Pharmacol 1985;108:
121-131.
14. Isenberg G, Klockner U. Calcium tolerant ventri- cular myocytes prepared by pre-incubation in a KB-medium. Pflügers Arch 1982;395:6-18.
15. Hamil OP, Marty A, Neher E, Sakmann B, Sigworth FJ. Improved patch-clamp techniques for high-resolution current recording from cells and cell-free membrane patches. Pflügers Arch 1981;
395:6-18.
16. Standen NB, Quayle JM. K+ channel modulation in arterial smooth muscle. Acta Physiol Scand 1998;164:549-557.
17. Odile CC, Walsh MP, Cole WC. Angiotensin II activation of protein kinase C decreases delayed rectifier K+ current in rabbit vascular myocytes. J Physiol 1996;493:689-700.
18. Ohba M, Sakamoto Y, Tomita T. The slow wave in the circular muscle of the guinea-pig stomach. J
Physiol 1992;451:387-417.
19. Rhee PL, Lee SJ, Kim SJ, So IS, Hwang SI, Kim KW. Effects of dopamine on the Ca2+ dependent K+
currents in isolated single gastric myocytes of the guinea-pig. Kor J Physiol 1993;27:139-150.
20. Mitra R, Morad M. Ca2+ and Ca2+-activated K+
current in mammalian gastric smooth muscle cells.
Science 1985;229:269-272.
21. Katzka DA, Morad M. Properties of calcium channels in guinea-pig gastric myocytes. J Physiol 1991;413:175-197.
22. Jun JY, Yeum CH, Yun PJ, Chang IY, Kim SJ, Kim KW. ATP-sensitive K+ current and its modulation of substance P in gastric myocytes isolated from guinea pig. Eur J Pharmacol 1998;
358:77-83.
23. 전제열, 염철호, 문성호 등. 위평활근에서 아세틸콜린 의 저분극기전에 관한 연구. 대한소화기학회지 1998;
31:421-431.
24. Farrugia G. Modulation of ionic currents in isolated canine and human jejunal circular smooth muscle cells by fluoxetine. Gastroenterology 1996;110:
1438-1445.
25. Noma A. ATP-regulated K+ channels in cardiac muscle. Nature 1983;305:147-148.
26. Ashcroft SJ, Ashcroft FM. Properties and functions of ATP-sensitive K channels. Cell Signal 1990;2:
197-214.
27. Quayle JM, Bonev AD, Brayden JE, Nelson MT.
Calcitonin gene-related peptide activated ATP- sensitive K+ currents in rabbit arterial smooth muscle via protein kinase A. J Physiol 1994;
475:9-13.
28. Kleppisch T, Nelson MT. Adenosine activates ATP-sensitive potassium channels in arterial myo- cytes via A2 receptors and cAMP-dependent protein kinase. Proc Natl Acad Sci USA 1995;92:12441- 12445.
29. Zhang L, Bonev AD, Nelson MT, Mawe GM.
Activation of ATP-sensitive potassium currents in guinea-pig gall-bladder smooth muscle by the neuropeptide CGRP. J Physiol 1994;478:483-491.
Jun, et al. Effects of Imipramine on Ionic Currents in Gastric Smooth Muscle of Guinea-pig 449
30. Bonev AD, Nelson MT. ATP-sensitive potassium channels in smooth muscle cells from guinea pig urinary bladder. Am J Physiol 1993;264:C1190- C1200.
31. Koh SD, Bradley KK, Rae MG, Keef KD, Horo- witz B, Sanders KM. Basal activation of ATP- sensitive potassium channels in murine colonic smooth muscle cells. Biophysics J 1998;75:1793- 1800.
32. Edwards G, Weston AH. The pharmacology of ATP-sensitive potassium channels. Annu Rev Phar-
macol Toxicol 1993;33:597-637.
33. Sakuta H. Inhibition by antidepressants of gliben- clamide-sensitive K+ currents in follicle- enclosed Xenopus oocytes. Can J Physiol Pharmacol 1994;
72:1586-1588.
34. Genikevitch VYa, Isenberg G. Caffeine-induced release and reuptake of Ca2+ by Ca2+ stores in myocyte from guinea-pig urinary bladder. J Physiol 1992;458:99-117.
35. Bean BP. Classes of calcium channels in vertebrate cells. Ann Rev Physiol 1990;51:367-384.