CONTENTS
Ⅰ. INTRODUCTION
Ⅱ. MATERIALS AND METHODS
Ⅲ. RESULTS
Ⅳ. DISCUSSION
Ⅴ. CONCLUSIONS REFERENCES KOREAN ABSTRACT EXPLANATION OF FIGURES
Ⅰ. INTRODUCTION
Xerostomia is rarely seen in isolation, but with time, the changes of quantity and quality of saliva can be devastating to oral health and may severely affect the general well-being and lifestyle of the patient. Indeed, the oral tissues become susceptible to infection, and the ability for oral function may be disturbed. In addition, a number of pathologic conditions can be developed, such as halitosis, burning mouth syndrome, gustatory changes and other oral inflammatory diseases.
1,2)Several systemic diseases, medication, and
psychologic processes may affect salivary flow and cause subjective oral dryness
3-6). Locker
7)reported that taking prescribed medication and experiencing a stressful life change had a significant effect on subjective oral dryness. And Bergdahl et al.
8)also confirmed that psychological factors, especially depression, are associated with hyposalivation and subjective oral dryness, and may play a crucial role in the etiology of the conditions.
There is a reported evidence that repeated stress, even of temporary duration, is able to influence directly or indirectly the morphofunctional state of the salivary glands
9), suggesting a functional linkage. It has been also shown that apoptosis (programmed cell death) of the submandibular gland may occur under the psychological stress
10). A linkage between stress and apoptosis has been well demonstrated in postmenopaused women in whom a significant alteration in the salivary compositions is attributed to symptomatic activation in response to psychologic stress
11). In order to have a better understanding of the pathologic mechanism in submandibular gland disease, and emphasizing the significant role of
An Expression of Caspase-3 on the Restraint Stress in the Rat Submandibular Gland
Won-Young Sohn, D.M.D., Sung-Hee Jung, D.M.D., Jae-O Cho, D.M.D.,M.S.D.,Ph.D.
Yang-Hyun Chun, D.M.D.,M.S.D.,Ph.D., Jung-Pyo Hong, D.M.D.,M.S.D., Ph.D.
Department of Oral Diagnosis & Oral Medicine, School of Dentistry, Kyung Hee University
* This study was supported by a grant of the Korea Health 21 R&D Project, Ministry of Health & Welfare, Republic of Korea, HMP-00-CH-10-0009.
stress in the disease, the present study was performed to investigate the expression of caspase-3, final effector in apoptotic death
12), with respect to apoptosis in cells of the submandibular gland under psychological stress by restraint.
Ⅱ. MATERIALS AND METHODS
1. Experimental animals and tissue preparation
Eighteen Sprague-Dawley rats (8-week-old) were purchased from Dae-Han Experimental Animal Research Center, Seoul, Korea. They were maintained at 20-23℃ and fed ad libitum on a normal laboratory diet. The rats were divided into 2 groups: restraint stress (15 rats) and control groups (3 rats). The rats of restraint stress group were placed in the plastic cages (150 x 50 mm) during the experiment. The stress groups were then sacrificed by neck-dislocation at 0, 1, 3, 5 and 7 day of the experiment and the submandibular glands were excised immediately. The excised tissues were frozen in dry ice in cryomolds, and stored at -70 ℃ until use.
2. Preparation of frozen sections
Serial frozen sections were made 4-8 ㎛ thickness, placed on positive-charged and RNA- free microscope slides, and stored at -70℃ until use. Frozen sections were allowed to come to room temperature (30 minutes) and fixed in solution of cold alcohol(99.9%) and acetone (1:1) for 10 minutes, kept refrigerated. The specimens were rinsed in three changes of 0.01M Tris buffered saline (TBS ; pH is 7.4±0.05. Tris : 0.05 M, NaCl : 0.15 m, Lab Vision Corp., U.S.A.) solution.
3. Immunohistochemistry
Anti-caspase-3(H-277 ; Santa Cruz Biotechno- logy, U.S.A.) used for immunohistochemistry is a rabbit polyclonal antibody raised against a
recombinant protein corresponding to amino acids 1-277 representing the full-length precursor form of caspase-3 of human origin. The anti-caspase-3 is reactive with the p11 and p20 subunits and the precursor of caspase-3(also designated CPP32) of mouse, rat and human origin.
Immunohistochemistry analysis was performed on the frozen section by the method as described previously.
After fixation, the sections were washed in TBS three times and incubated with 3% H
2O
2for 10 minutes at room temperature. And then the slides washed with distilled water (D.W.) three times. For the protein blocking, 150 μl normal serum in 10 ml TBS for 20 minutes at room temperature. And then the sections were incubated with the rabbit anti-caspase-3 in humidity at room temperature that was diluted 1:50 with concentrated-reagent diluents (PBS with carrier protein and 0.09%
sodium azide). After 2hr, the specimen were rinsed with TBS solution and then were incubated with biotinylated goat anti-rabbit immunoglobulin for 40 minutes at room temperature. The slides were then washed in TBS and incubated with avidin- biotin-peroxidase complex (ABC kit ; VECTASTAIN
®, Vector Lab., Burlingame, CA, USA) for 40 minutes. Thereafter, the tissues were rinsed with TBS solution and imunohistochemical reactions were detected by color development using 3-amino-9-ethylcarbazole solution (AEC kit ; Vector Lab., Burlingame, CA, USA). After washing in D.W., the slides were counterstained with hematoxylin and mounted in VECTASHIELD
®Mounting Medium for retaining tissues. Thereafter, immunohistochemical reactions were examined under the microscope and photographed.
Ⅲ. RESULTS
To localize the expression of caspase-3 in the
submandibular glands, the glands from each group
were fixed and processed for immunohisto-
chemistry using the specific antibody against
caspase-3.
As time goes by, amount of caspase-3 was decreased and apoptosis was increased in the rat submandibular gland under the restraint stress. In other words, it can be found that cell destruction (apoptosis) was followed by the expression of caspase-3.
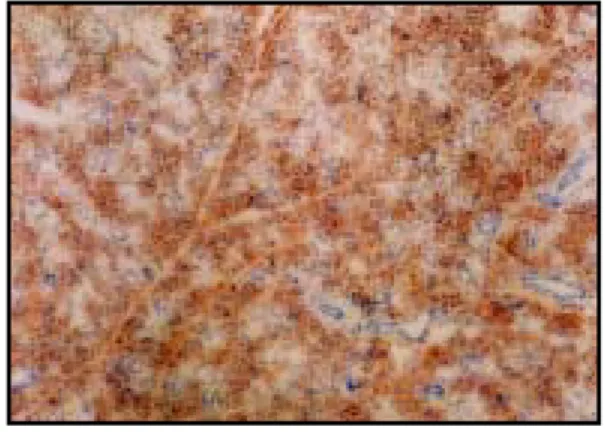
In the normal control group, caspase-3 immunoreactivity was not detected at any time during the experiment (Fig.1, 7, 13).
In the restraint stress group, the weak expression of caspase-3 initiated in intracellular space and normal cellular features observed immediately after application of the stress (Fig.2, 8, 14)
In the restraint stress group, immunoreaction of caspase-3 appeared to be strong positive reaction on the 1 day after the application of the stress (Fig.3, 9, 15), and then gradually decreased (Fig.4-6, 10-12, 16-18).
In the restraint stress group, there are apoptosis of glandular cells on the 5th day after the application of the stress, and the cells almost destructed on the 7th day after the restraint stress.
Ⅳ. DISCUSSION
This study confirmed that psychological stress is associated with dysfunction in salivary gland. An expression of caspase-3, known as a key enzyme in apoptosis indicated apoptosis occurred in submandibular gland under the restraint stress. As time goes by, amount of caspase-3 was decreased and apoptosis was increased. So, like our previous investigations
10,13-15), the present study provides that psychological stresses play a crucial role in the etiology of xerostomia and hyposalivation.
When etiological factors were considered in earlier investigations of hyposalivation and subjective oral dryness in patients, much emphasis has been placed on medication, and psychological factors have been largely disregarded
8). Physical illness, however, is frequently associated with depression
16), and analysis of empirical data has shown that depressive disorders are among the most common diseases, comparable with
hypertension and diabetes
17). Subjective oral dryness
18)and reduced salivary flow
3)have been found in many individuals with depression, and clinical improvement from the depression brought about by electroconvulsive therapy, but not drug treatment, has been found to increase salivary flow
19). And there are another reports that alarm-like reactions, depression and anxiety inhibit, while pleasurable, relaxed sensations promote to increasing salivation.
20,21)Black
22)addressed that no disease is exempt from psychological influences, and, as a corollary, all diseases have psychological repercussions.
It has been suggested that psychological stress could play a key role in triggering suppression of the immune system and alter the susceptibility of animal and man to infectious agents, influencing the onset, course and outcome of certain infectious pathologies
23). Recently, quite a few studies have shown that stresses are closely associated with diseases, especially with oral diseases.
A few studies have suggested that stress is strongly associated with oral diseases. Chun and Hong
24)indicated that stress causes various forms of diseases in the region including oral psychosomatic diseases in which emotional stress appears to play a major role, oral diseases in which psychologic factors appear to play a role, oral infections where emotional stress is a significant predisposing factor, oral lesions induced by neurotic habits inflicting trauma, neurotic oral symptoms, and oral pain induced by emotional stress.
Causes including stress may reduce salivary flow and increase subjective oral dryness by mimicking autonomic nervous system actions, or by acting directly on the cellular processes necessary for salivation
5). Recently, apoptosis attract public attention for the representitive the cellular precesses.
Apoptosis or programmed cell death (PCD) is a
genetically regulated, cellular suicide mechanism
that plays a crucial role in development and in the
defense of homeostasis
25). Cells respond to a variety
of disparate signals by committing suicide through
a series of dramatic but remarkably uniform events.
Morphologically, cells undergoing apoptosis demonstrate nuclear/cytoplasmic condensation and membrane protrusions. These initial changes are followed by fragmentation of the nuclear contents and subsequent encapsulation of these fragments into "apoptotic bodies" that are quickly and unobtrusively consumed by adjacent cells, thereby leaving little trace of the apoptotic cell's prior existence
26). Biochemically, apoptotic cells are characterized by reduction in the mitochondrial transmembrane potential, intracellular acidification, production of reactive oxygen species, externalization of phosphatidylserine residues in membrane bilayers, selective proteolysis of a subset of cellular proteins, and degradation of DNA into internucleosomal fragments
27-32).
Although apoptosis of salivary gland cells has been demonstrated in several pathological conditions, the role of apoptosis in the postnatal development of the salivary glands is largely unknown. Hecht et al.
33)studied the development of the rat submandibular gland (SMG) during its transition from the perinatal stage to the mature adult stage. They suggested that the loss of Type I cells and reduction of SMG immunoreactivity during development of the intercalated ducts of the adult rat SMG is due, at least in part, to apoptosis.
Scott et al.
34)also suggested that pathological atrophy is largely similar to physiological atrophy, providing a mechanism for acinar cell survival under adverse conditions, with the possibility of eventual recovery. Their results confirmed that totally obstructed glands undergo a rapid, progressive severe atrophy amounting to absolute losses of over 85% of acinar tissue by two weeks.
It has been reported that morphology and function of the salivary glands are vulnerable to stress: the inhibition of salivary gland function is due to central influences from higher centers acting on the salivary centers and thereby suppressing reflex activity
35); the submandibular gland is more sensitive to cold than the sublingual gland36);
repeated stress, even of temporary duration, is able
to influence directly or indirectly the morphofunctional state of the submandibular gland
9); acute emotional painful stress enhances free-radical lipid oxidation in the salivary gland tissues and reduces the activities of antioxidant enzymes in it
37). In addition, the submandibular gland is reportedly more age-related, in the pattern of fluid secretion with an intense gustatory stimulus than parotid gland
38).
Recently, there are several studies about decreased salivary flow resulted from correlation between psychological stress and apoptosis in human submandibular gland with respect to clusterin
10,13-15)and caspases
39).
These apoptosis include the activation of an intrinsic cell death apparatus that has been exquisitely conserved during evolution. At the core of this death apparatus is a novel family of proteases related to the Caenorhabditis elegans cell death gene product CED-3, the so-called caspases (cysteine proteases with aspartate-specificity), that are universal effectors of apoptotic cell death
25). With the cloning of ced-3 (Caenorhabditis elegans cell-death gene) came the observation that it encoded a protease homologous to the mammalian ICE (interleukin-1beta-converting enzyme), a novel cysteine protease with aspartate specificity required for proteolytic conversion of pro-interleukin-1 into its mature form
40-42). Together with the concurrent finding that apoptosis could be induced in mammalian cells by transient expression of ICE or ced-3
43), these studies provided the first inkling that a conserved family of proteases might be a critical component of the cell death machinery. At the present time, at least 10 additional mammalian homologs of ced-3 have been published. Given the considerable confusion generated by the rapid growth of this family, the trivial names of these cysteine proteases with aspartate specificity (caspases) have been replaced with a uniform nomenclature: caspases-1 through -11
44,45).
Caspases can be activated by one of two
potentially interacting and reversible pathways: a
mitochondrial and a death receptor route. By unclear mechanisms, apoptotic stimuli such as DNA damaging agents trigger the mitochondrial release of cytochrome c into the cytosol, an event that is antagonized by Bcl-2 and its antiapoptotic relative Bcl-xL. In the cytoplasm, cytochrome c binds to the CED-4 (Caenorhabditis elegans cell death protein) homolog Apaf-1 (precursor protein for activating caspases), which interacts with both death effectors (pro-caspase-9) and death antagonists (Bcl-xL) to form the apoptosome.
Binding of cytochrome c and ATP to Apaf-1 somehow promotes its ability to catalyze the proteolytic activation of pro-caspase-9, whereas Bcl-xL presumably inhibits this function. Active caspase-9, in turn, proteolytically activates the downstream caspase terminators such as caspase-3 that kill cells by cleaving key intracellular proteins
25).
There is several evidence that caspase-3 may act as a distal effector of the apoptotic machinery.
Caspase-3 has been shown to cleave the death substrate poly(ADP-ribose) polymerase (PARP), in addition to being inhibitable by the cowpox serpin, CrmA46). Activated caspase-3, comprised of p17 and p12 subunits, was purified from cell extracts using a tetrapeptide aldehyde inhibitor corresponding to amino acids at the PARP cleavage site.
47)Depletion of caspase-3 from these extracts abrogated their apoptotic potential in vitro.
47)This apoptotic activity could be restored by adding back purified caspase-3 to the depleted extracts.
47)Porter and Janicke
48)studied the specific requirements of caspase-3 in apoptosis. They suggested that caspase-3 is essential for normal brain development and is important or essential in other apoptotic scenarios in a remarkable tissue-, cell type- or death stimulus-specific manner.
Caspase-3 is also required for some typical hallmarks of apoptosis, and is indispensible for apoptotic chromatin condensation and DNA fragmentaion in all cell types examined. Thus, caspase-3 is essential for certain processes associated with the dismantling of the cell and the
formation of apoptotic bodies, but it may also function before or at the stage when commitment to loss of cell viability is made.
Essmann et al.
49)also suggested that the differential regulation of the homologue GDP dissociation inhibitors Rho-GDI 1 and D4-GDI during drug-induced apoptosis by proteolysis mediated by caspase-3. Owing to their crucial role as modulators of Rho GTPases, that might in turn have a significant impact on the mechanisms that induce the cytoskelotal and morphological changes in apoptotic cells.
Marissen et al
50)reported that the cleavage of eukaryotic translation initiation factor 4G (elF4G) by caspase-3 was described as a possible event contributing to translation inhibition that could regulate translation during apoptosis. Hartmann et al.
12)reported that caspase-3 act as a vulnerability factor and final effector in apoptotic death of dopaminergic neurons in Parkinson's disease.
The present study has shown that when salivary glands are affected by a variety of stimuli, the cells of the salivary glands may adapt to them by eliciting various cellular responses such as structural and biochemical transformations. But if the stimulus is lying outside an acceptable range of normality, it may cause such changes at the clinical and subclinical levels as alterations in the production, flow rate, the response to stimuli, the components and the immune function of saliva.
In summary, stress may cause apoptosis depending upon degree, duration of the stress applied and host's adaptability. Caspase-3 can be expressed as a distal effector of the apoptotic machinery when the stress far exceed the range. A excessive stress may induce apoptosis in the cell.
Since stress-induced apoptosis alters the normal
salivary physiology, the present study was
proposed that pathologic change of the salivary
glands can be initiated by stress and then
exacerbated by apoptosis. However, further studies
are necessary how the stress act as triggering
factor for inducing caspase-3 to activate through
what mechanism. And it is necessary to study
about the change of the neural and hormonal system under psychological stress.
Ⅴ. CONCLUSIONS
Dry mouth (xerostomia) is a symptom, but most people who suffered from dry mouth feel like a disease. So, xerostomia acts as a stressful life event, however, the patients who suffered from dry mouth are mostly under the stress.
Despite extensive studies on the relationship between stress and the oral health have been conducted, the pathology of dry mouth, halitosis and burning mouth syndrome remains to be elucidated. In order to have a better understanding of the pathology in orofacial diseases, the present study was undertaken to observe the stress- induced alterations in the submandibular gland and the expression of caspase-3 which is believed to be a enzyme responsible for controlling apoptosis.
Eighteen Sprague-Dawley rats (8-week-old) were purchased from Dae-Han Experimental Animal Research Center, Seoul, Korea. They were maintained at 20-23℃ and fed ad libitum on a normal laboratory diet. The rats were divided into 2 groups: restraint stress (15 rats) and control groups (3 rats). The rats of restraint stress group were placed in the plastic cages (150 x 50mm) during the experiment. The stress groups were then sacrificed by neck-dislocation at 0, 1, 3, 5 and 7 day of the experiment and the submandibular glands were collected immediately. The excised tissues were frozen and stored at -70℃ until use.
The expression of caspase-3 in the submandibular gland tissues was observed by immunohisto- chemistry method. The results were as follows:
1. In the normal control group, caspase-3 immunoreactivity was not detected at any time during the experiment .
2. In the restraint stress group, strong positive immunoreaction of caspase-3 was noticed on 1 day after the application of the stress, and then gradually decreased.
3. In the restraint stress group, there are apoptosis of glandular cells 5 day after the application of the stress, and the cells almost destructed after 7 day after the restraint stress.
The overall results indicate that caspase-3 occurs as a result of cell response to psychological stress for apoptosis but then gradually disappear. It is likely that if stress far exceeds the acceptable range for adaptation, apoptosis may occur in the submandibular gland and cause pathologic changes of the gland. Through this experiment, caspase-3 was confirmed as a very specific tracing protein during stress condition, so the patients under excessive stress who have symptoms resulted from dysfunction of salivary gland, may be diagnosed with this protein, caspase-3. Consequently, we propose that stress can be a direct causal factor in the patient groups who show salivary disturbances under severe emotional stress.
REFERENCES
1. Field, E.A., Longman, L.P., Bucknall, R., Kaye, S.B., Higham, S.M., and Edgar, W.M. : The establishment of a xerostomia clinic: a prospective study. Br. J. Oral Maxillofac. Surg., 35:96-103, 1997.
2. Sreebny, L.M. : Recognition and treatment of salivary induced conditions. Int. Den. J., 39:197-204, 1989.
3. Mathew, R.J., Weinman, M., and Claghorn, J.L. : Xerostomia and sialorrhea in depression. Am. J.
Psychiatry., 136:1476-1477, 1979.
4. Sreebny, L.M. and Valdini, A. : Xerostomia. Part II;
relationship to other oral symptoms, drugs, and diseases. Oral Surg. Oral Med. Oral Pathol., 68:291-338, 1989.
5. Sreebny, L.M. and Schwartz, S.S. : A reference guide to drugs and dry mouth. 2nd ed. Gerodontology, 14:33-47, 1997.
6. Navazesh, M., Christensen, C.M. : A comparison of whole-mouth resting and stimulated salivary measurement procedures. J. Dent. Res., 61:1158-1162, 1996.
7. Locker, D. : Subjective reports of oral dryness in an older adult population. Community Dent. Oral Epidemiol., 21:165-168, 1993.
8. Bergdahl, M. and Bergdahl, J. : Low unstimulated
salivary flow and subjective oral dryness: association with medication, anxiety, depression, and stress. J.
Dent. Res., 79(9):1652-1658, 2000.
9. Pellegrini, A., Grieco, M., Materazzi, G., Gesi, M., and Ricciardi, M.P. : Stress-induced morphohistochemical and functional changes in rat adrenal cortex, testis and major salivary glands. Histochem. J., 30:695-701, 1998.
10. Park, H.K., Chun, Y.H., Lee, J.Y., Cho, H.G. and Hong, J.P. : Expression of Clusterin in the Salivary Gland under Restraint Stress. Kor. J. Stress Res., 6(2):33-44, 1998.
11. Ben, A.H., Gottlieb, I., Ish-Shalom, S. et al : Oral complaints related to menopause. Maturitas., 24:185-189, 1996.
12. Hartmann, A., Hunot, S., Michel, P.P. et al : Caspase-3: A vulnerability factor and final effector in apoptotic death of dopaminergic neurons in Parkinson's disease. Proc. Natl. Acad. Sci. USA., 14;97(6):2875-80, 2000.
13. Chun, Y.H. and Hong, J.P. : The molecular biological study on change of clusterin (SGP-2) in the salivary glands of streptozotocin-induced diabetic rats under stress. Kor. J. Stress Res., 5:13-32, 1997.
14. Kim, S.H., Hong, J.P. : Clusterin (SGP-2) in the salivary gland of insulin injected rats under stress. J.
Kor. Acad. Oral Med., 23:309-326, 1998.
15. Chung, W.B., Cho, H.G., Hong, J.P. : The expression of clusterin (SGP-2) to the stress on the salivary glands of rats. J. Kor. Acad. Oral Med., 22:395-408, 1997.
16. Cavanaugh, S., Clark, D.C., and Gibbons, R.D. : Diagnosing depression in the hospitalized medically ill. Psychosomatics., 24:809-815, 1983.
17. Judd, L.L. : Mood disorders in the general population represent an important and worldwide public health problem. Int. Clin. Psycopharmacol., 10:5-10, 1995.
18. Bergdahl, M., Bergdahl, J., and Johansson, I. : Depressive symptoms in individuals with idiopathic subjective dry mouth. J. Oral Pathol. Med., 26:448-450, 1997.
19. Bolwig, T.G. and Rafaelsen, O.J. : Salivation in affective disorders. Pshychol. Med., 2:232-238, 1972.
20. Dobrosielski-Vergona, K. : Biology of the salivary glands. Boca Raton, 1993, CRC Press, Inc.
21. Lynch, M.A., Brightman, V.J., and Greenber, M.S. : Burket's oral medicine. 9th Ed., philadelphia, 1994, Lippincott Co., pp. 399-400.
22. Black, P.H. : Psychoneuroimmunology: brain and
immunity. Sci. Am. Sci. Med., 2:16-25, 1995.
23. Biondi, M. and Zannino, L.G. : Psychological stress, neuroimmunomodulation, and susceptibility to infectious diseases in animals and man. A review.
Psychother. Psychosom., 66:3-26, 1997.
24. Chun, Y.H. and Hong, J.P. : Stress and oral diseases.
Kor Stress Res., 3(1):57-72, 1995.
25. Cryns and Yuan : Protease to die for. Genes Dev.
12:1551-1570, 1998.
26. Kerr, J.F.R., Wyllie, A.H., and Currie, A.R. : Apoptosis: a basic biological phenomenon with wide-ranging implications in tissue kinetics. Br. J.
Cancer 26:239-257, 1972.
27. Wyllie, A.H., Morris, R.G., Smith, A.L. and Dunlop, D.
: Chromatin cleavage in apoptosis: Association with condensed chromatin morphology and dependence on macromolecular synthesis. J. Pathol., 142:66-77, 1984.
28. Hockenbery, D.M., Oltvai, Z.N., Yin, X.M., Milliman CL and Korsmeyer SJ : Bcl-2 functions in an antioxidant pathway to prevent apoptosis. Cell 75:241-251, 1993.
29. Lazebnik, Y.A., Kaufmann, S.H., Desnoyers, S., Poirier, G.G., and Earnshaw, W.C. : Cleavage of poly(ADP-ribose) polymerase by a proteinase with properties like ICE. Nature, 371:346-347, 1994.
30. Martin, S., Reutelingsperger, C., McGahon, A. et al : Early redistribution of plasma membrane phosphatidylserine is a general feature of apoptosis regardless of the initiating stimulus: Inhibition by overexpression of Bcl-2 and Abl. J. Exp. Med., 182:1545-1556, 1995.
31. Gottlieb, R.A., Nordberg, J., Skowronski, E., and Babior, B.M. : Apoptosis induced in Jurkat cells by several agents is preceded by intracellular acidification. Proc. Natl. Acad. Sci., 93:654-658, 1996.
32. Zamzami, N., Susin, S.A., Marchetti, P. et al : Mitochondrial control of nuclear apoptosis. J. Exp.
Med., 183:1533-1544, 1996.
33. Hecht, R., Connelly, M., Marchetti, L., Ball, W.D., and Hand, A.R. : Cell death during development of intercalated ducts in the rat submandibular gland.
Anat. Rec., 258:349-358, 2000.
34. Scott, J., Liu, P., and Smith, P.M. : Morphological and functional characteristics of acinar atrophy and recovery in the duct-ligated parotid gland of the rat.
J. Dent. Res., 78:1711-1719, 1999.
35. Garrett, J.R. : The proper role of nerves in salivary secretion. J. Dent. Res., 66:387-97, 1987.
36. Fedelich, M.A., Rins de David, M.L. : Stress and the
salivary glands. Rev. Fac. Odontol. Univ. Nac.
(Cordoba), 17:55-69, 1989.
37. Tarasenko, L.M., Devatkina, T.A., Tsebrzhinskii, O.I., Grebennikova, V.F., and Mel'nikova, S.V. : Reaction of the salivary glands to acute stress. Fiziol. Zh., 36:104-6, 1990.
38. Sokolenko, V.N. and Silenko, IuI. : The free-radical involvement of the salivary gland in stress.
Stomatologia (Mosk), 74:17-9, 1995.
39. Chung, W.B., Jung, S.H., Chun, Y.H., Lee, J.Y. and Hong, J.P. ; Caspase-3 expression in the submandibular gland of rat under restraint stress, J.
of Korean Academy of Oral Medicine 25(3):265-278, 2000.
40. Yuan, J., Shaham, S., Ledoux, S., Ellis, H.M., and Horvitz, H.R. : The C. elegans cell death gene ced-3 encodes a protein similar to mammalian interleukin-1 beta-converting enzyme. Cell, 75:641-652, 1993.
41. Thornberry, N.A., Bull, H.G., Calaycay, J.R. et al : A novel heterodimeric cysteine protease is required for interleukin-1 beta processing in monocytes. Nature, 356:768-774, 1992.
42. Cerretti, D.P., Kozlosky, C.J., Mosley, B. et al : Molecular cloning of the interleukin-1 beta converting enzyme. Science, 256:97-100, 1992.
43. Miura, M., Zhu, H., Rotello, R., Hartwieg, E.A., and Yuan, J. : Induction of apoptosis in fibroblasts by IL-1 beta-converting enzyme, a mammalian homolog of the C. elegans cell death gene ced-3. Cell, 75:653-660, 1993.
44. Alnemri, E.S., Livingston, D.J., Nicholson, D.W. et al : Human ICE/CED-3 protease nomenclature. Cell, 87:171, 1996.
45. Wang, S., Miuyra, M., Jung, Y.k. et al : Murine caspase-11, an ICE-interacting protease, is essential for the activation of ICE. Cell,92:501-509, 1998.
46. Teware, M., Quan, L.T., O'Rourke, K. et al : Yama/CPP32 beta, a mammalian homolog of CED-3, is a CrmA-inhibitable protease that cleaves the death substrate poly (ADP-ribose) polymerase. Cell, 81:801-809, 1995.
47. Nicholson, D.W., Ali, A., Thornberry, N.A. et al : Identification and inhibition of the ICE/CED-3 protease necessary for mammalian apoptosis. Nature, 376:37-43, 1995.
48. Porter, A.G. and Janicke, R.U. : Emerging roles of caspase-3 in apoptosis. Cell Death Differ, 6:99-104, 1999.
49. Essmann, F., Wieder, T., Otto, A. et al : GDP dissociation inhibitor D4-GDI(Rho-GDI 2), but not thes homologous rho-GDI 1, is cleaved by caspase-3 during drug-induced apoptosis. Biochem. J., 15:777-783, 2000.
50. Marissen, W.E., Guo, Y., Thomas, A.A., Matts, R.L., and Lloyd, R.E. : Identification of caspase 3-mediated cleavage and functional alteration of eukaryotic initiation factor 2alpha in apoptosis. J. Biol. Chem., 31:9314-9323, 2000.
Corresponding Author : Jung-Pyo Hong, Professor, Department of Oral Diagnosis & Oral Medicine, School of Dentistry, Kyung Hee University, 1 Hoegi-Dong, Dongdaemun-Ku, Seoul 130-701, Korea
국문초록
구속에 따른 스트레스가 타액선 조직의 Caspase-3 발현에 미치는 영향에 관한 면역조직화학적 연구
경희대학교 치과대학 구강내과학 전공
손 원 영․정 성 희․조 재 오․전 양 현․홍 정 표
구취, 구강작열감, 그리고 구강건조증은 스트레스와 많은 연관성을 가지고 있다. 이러한 스트레스에 의하여 인 체 조직은 apoptosis등의 다양한 변화가 초래될 수 있다. Apoptosis는 세포의 DNA가 분절되어 발생되는 것으로 서 caspase-3 가 결정적인 역할을 한다고 생각된다. 이에 본인은 백서에 구속 스트레스를 가하여 caspase-3 를 관찰함으로써 타액선 조직의 변화기전을 구명하고자 본 실험을 시행하였다.
웅성백서 (Sprague-Dawley, 8주, 238-250g) 18마리를 실험 전 기간에 걸쳐 구속스트레스를 가하고, 각각 실험 후 즉일, 1일, 3일, 5일, 7일에 희생시켰다. 그 후 실험동물의 악하선을 절취하여 동결절편을 제작한 후, caspase-3 에 대하여 면역조직화학법을 시행하여 관찰하였다.
1. 정상대조군에서 caspase-3는 발현되지 않았다.
2. 구속스트레스 부여 1일군에서 caspase-3가 강반응을 보였고, 이후 점차 감소되어 7일군에서는 거의 소실되었 다.
3. 구속스트레스 부여 5일후부터 세포의 파괴가 관찰되었으며, 7일 후에는 현저하였다.
본 실험을 통하여 caspase-3의 강한 발현과 세포의 파괴는 기존의 구속스트레스 후 clusterin의 발현과 조직학
적인 apoptosis가 발견되었던 것에 대한 과정 및 기전을 확인할 수 있었다. 따라서 정서적 스트레스가 과도한 환
자군에서 타액선 기능저하로 인한 장애가 관찰될 경우에는 스트레스가 직접적인 원인요소로 작용할 수 있다는
가능성을 제시할 수 있다.
EXPLANATION OF FIGURES
Fig. 1. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the control group (X40).
normal cell, nucleus, acinar cell and ductal cell in submandibular gland.
Fig. 2. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress immediatly after the application of the stress (X40). Weak expression of caspase-3 (brown spots) and normal cellular feature.
Fig. 3. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 1 day after the application of the stress (X40). Strong expression of caspase-3 and normal cellular feature.
Fig. 4. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 3 days after the application of the stress (X40). Strong and clear expression of caspase-3
Fig. 5. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 5 days after the application of the stress (X40). Weakened expressions of caspase-3 and unclear boundary between acinar and ducatal cells.
Fig. 6. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 7 days after the application of the stress (X40). Caspase-3 is almost disappeared and there are throughout apoptosis in gland cells.
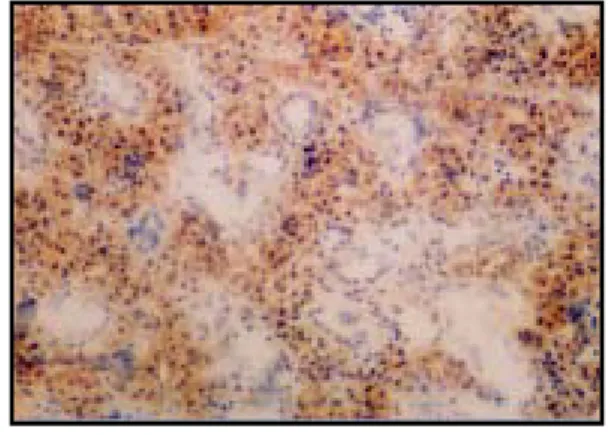
Fig. 7. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat of the control group (X100).
Fig. 8. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress immediatly after the application of the stress (X100). Weak expression of caspase-3 (brown spots).
Fig. 9. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 1 day after the application of the stress (X100). Strong expression of caspase-3 in intracellular space.
Fig. 10. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 3 days after the application of the stress (X100). Still strong and clear expression of caspase-3.
Fig. 11. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 5 days after the application of the stress (X100). Weak expression of caspase-3 and cell destructions (apoptosis) can be observed.
Fig. 12. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 7 days after the application of the stress (X100). Cell destructions (apoptosis) can be detected in throughout salivary glands.
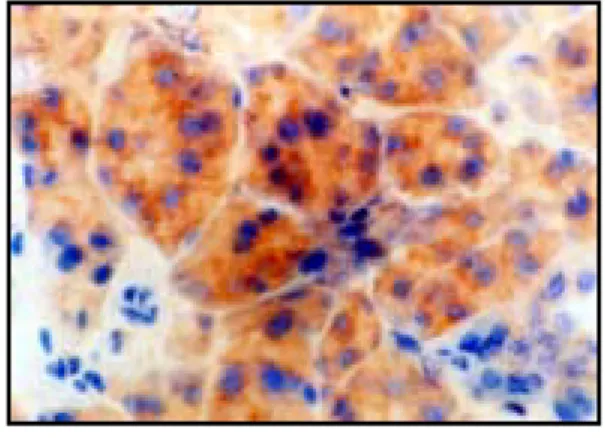
Fig. 13. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under of the control group (X400). Normal cytoplasm and nulceus.
Fig. 14. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress immediatly after the application of the stress (X400). Weak expression of caspase-3 in intracellular space (brown spots)
Fig. 15. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 1 day after the application of the stress (X400). Clear, brown and roundish expression of caspase-3 in intracellular space. normal cytoplasm and nucleus.
Fig. 16. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 3 days after the application of the stress (X400). Still clear and strong expression of caspase-3 in intracellular space of salivary gland. Normal cellular features.
Fig. 17. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 5 days after the application of the stress (X400). Weak and unclear expression of caspase-3, Cell destruction (apoptosis) like chromatin condensation and cell membrane rupture can be detected.
Fig. 18. Immunohistochemistry microscopy of caspase-3 in the submandibular gland of the rat under restraint stress 7 days after the application of the stress (X400). Throughout cell destruction and unclear boundary of cytplasm. Disrupted nuclear and cellular features morphologically.
Fig. 1. Fig. 2.
Fig. 3. Fig. 4.
Fig. 5. Fig. 6.
Fig. 7. Fig. 8.
Fig. 9. Fig. 10.
Fig. 11. Fig. 12.
Fig. 13. Fig. 14.
Fig. 15. Fig. 16.
Fig. 17. Fig. 18.