한수지 54(3), 298-310, 2021
298
Copyright © 2021 The Korean Society of Fisheries and Aquatic Science pISSN:0374-8111, eISSN:2287-8815 Korean J Fish Aquat Sci 54(3),298-310,2021
Original Article
서 론
뱀장어
(Anguilla japonica)
는강하성어종으로한국,
일본,
대 만,
중국,
필리핀북부에분포하며(Tesch, 2003; Kuroki et al.,
2009),
동아시아에서상업적으로매우중요한양식대상어종이다
(Hsu et al., 2015).
뱀장어는괌인근해산에서산란및부화한후북적도해류와쿠로시오해류를따라이동하며
(Aoyama
et al., 2014),
자어기인yolk-sac larvae, pre-leptocephalus, lep- tocephalus, metamorphosis
단계를거쳐(Mochioka, 2003)
치 어인실뱀장어(Glass eel)
가되어강으로소상한다(Tsukamoto et al., 2003).
하지만해양환경및기후변화,
서식지파괴,
남 획등다양한원인으로뱀장어자원은급격히감소하고있으며(Tsukamoto et al., 2009; Chen et al., 2014),
이로인해소상하는실뱀장어포획에의존하고있는뱀장어양식산업은큰위기 에처해있다
(Yamada et al., 2019).
이에따라,
극동산뱀장어의 자원보호와양식산업의안정화를위한뱀장어인공종자대량 생산기술개발이시급히요구되고있다.
뱀장어인공종자생산 연구는1960
년대부터시작되어(Hibiya, 1970)
현재까지도활 발히진행되고있지만,
독특한생활사를가진뱀장어자어에적 합한사육시스템및먹이원을찾지못하여아직실험실수준의소규모생산에그치고있다
.
뱀장어자어는자연에서marine
snow (
해중설)
라고불리는표층에서떨어지는유기물(Otake et al., 1993; Miller et al., 2013)
및피낭동물인Larvacean
의집(Westerberg, 1990)
등을섭취하는것으로보고되고있으나,
해 당물질을사육환경에적용시키는것은매우어렵다.
따라서 로티퍼,
동물성플랑크톤,
홍합유생등다양한먹이원적용을인공생산 뱀장어(Anguilla japonica) 자어의 소화기관 발달 특성
신민규 1,2 ·김신권 1 ·이배익 1 ·최윤희 2 ·유용운 1 *
1국립수산과학원 양식연구과, 2부경대학교 해양바이오신소재학과
Histological Development of the Digestive System in Artificially Produced Anguilla japonica Larvae
Min Gyu Shin
1,2
, Shin-Kwon Kim1
, Bae-Ik Lee1
, Youn Hee Choi2
and Yongwoon Ryu1
*1
Aquaculture Research Division, National Institute of Fisheries Science, Busan 46083, Korea2
Department of Fisheries Biology, Pukyong National University, Busan 48513, KoreaFreshwater eel Anguilla japonica is a commercially important species in East Asia. However, given the continuously decreasing resources due to many problems as climate change, habitat destruction and overfishing, mass seed pro- duction is crucial. In particular, the eel larvae present a unique life history, spending prolonged periods in the Pacific Ocean. Therefore, finding suitable feed that lasts through the long larval stages is rather difficult. In the present study, we histologically observed the development of digestive organs for 50 days after hatching (DAH). The endogenous reserves were completely absorbed at 7 DAH, and the first exogenous feeding started thereafter. The teeth appeared at 4 DAH and dramatically changed from needle-like to broad-based types by 50 DAH. The intestine remained as straight gut until 50 DAH. The intestinal cilia appeared first at 6 DAH and their density continuously increased there- after. Interestingly, goblet cells appeared in the intestine, but not in the esophagus, where these cells are commonly observed in other teleosts. In addition, high amounts of eosinophil substances, assumed to be zymogen granules, ac- cumulated in the pancreas, suggesting a specific digestive strategy of this species. Overall, these observations largely update the previous reports and can be useful to improve A. japonica seed production.
Keywords: Anguilla japonica , Leptocephalus, Ontogeny, Digestive organs, Histology
*Corresponding author: Tel: +82. 51. 720. 2181 Fax: +82. 51. 720. 2489 E-mail address: [email protected]
This is an Open Access article distributed under the terms of the Creative Commons Attribution Non-Commercial Licens (http://creativecommons.org/licenses/by-nc/3.0/) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
Received 4 March 2021; Revised 5 April 2021; Accepted 27 April 2021
저자 직위: 신민규(대학원생 및 연구원), 김신권(연구사), 이배익(연구사), 최 윤희(교수), 유용운(연구사)
https://doi.org/10.5657/KFAS.2021.0298
Korean J Fish Aquat Sci 54(3), 298-310, June 2021
뱀장어 자어의 소화기관 발달 특성
299
시도했었지만뱀장어자어의성장과생존에는적합하지않았 고
(Satoh, 1979; Yu et al., 1993; Tanaka et al., 1995),
이후곱 상어(Squalus acanthias)
알을기반한slurry
형태의인공사료 를개발하여실뱀장어까지사육하는데성공하였다(Tanaka et al., 2001; Tanaka et al., 2003).
하지만, Hsu et al. (2015)
에따 르면곱상어알기반의사료는뱀장어자어의소화및흡수능 력에적합하지않으며,
실제해당사료를사용하여생산한인 공생산자어는자연개체에비해느린성장과낮은생존율을나 타낸다(Tsukamoto, 2014).
따라서뱀장어자어에적합한먹이 원개선은종자대량생산을위해해결해야할큰문제중하나 로남아있다.
어류의자어는성장과생존을위해적합한먹이포획
,
섭취,
소 화,
흡수를위한구조적그리고기능적으로잘발달된소화기관 이필요하다(Kjørsvik et al., 2004).
하지만,
초기자어의소화 기관은그기능이제한적이기때문에(Rønnestad et al., 2013;
Yúfera et al., 2018)
완전히기능하기까지일련의발달변화가 필요하며(Govoni et al., 1986),
그시기는어종에따라차이가 있다(Treviño et al., 2011).
특히,
뱀장어자어는마린스노우를 먹으며일반양식어류에적용되는많은초기먹이원들이전혀 적용되지않기때문에,
일반적인어류자어와는다른독특한초 기소화특성을가지고있을것으로추측된다.
뱀장어자어의소화기관에대한연구는자연산의경우전장
11-48 mm
개체의장미세구조가보고되었으며(Otake, 1996),
인공산에서는부화후6-13
일초기자어의발달특성에대하여 보고된바있다(Kurokawa et al., 1995; Otake, 1996; Kuroka- wa and Pedersen, 2003; Ozaki et al., 2006; Yoshimatsu, 2011).
이들대부분의연구는초기자어를대상으로한단기간의관찰 이거나
,
가느다란(slender)
체형의얇고긴소화관으로인하여 일부기관만관찰되어왔기때문에소화기관전체의발달특성 을이해하기에부족함이있었다.
따라서,
독특한생태생리특성 을가지는뱀장어자어의소화발달특성을이해하고이를기반 으로적합한인공사료를개발하기위해서는긴자어기에걸친 소화기관의전반적인관찰이요구된다.
이연구에서는뱀장어 자어의yolk-sac larvae, pre-leptocephalus, leptocephalus
단계 를포함하는부화후0
일부터50
일까지자어의소화기관을조 직학적으로관찰하여뱀장어자어의초기생활사이해및인공 종자생산기술개발에필요한기초정보를축적하고자하였다.
재료 및 방법
친어 관리 및 부화자어 생산
뱀장어부화자어의생산을위하여암컷친어는전라남도부 안에서
2018
년9
월에채포한자연산1
개체(1,042 g/BW)
를사 용하였으며,
수컷은자연산실뱀장어를채포하여양식장에서3
년간키운1
개체(187 g/BW)
를이용하였다.
확보된친어들은약
20°C
의해수에천천히순치시켰다.
암수친어의성성숙유도를위하여
Kim et al. (2007)
의방법을따라암컷은6
주동 안매주연어뇌하수체추출물(Salmon pituitary extract, SPE;
Yasaka, Tokyo, Japan)
을20 mg/fish,
수컷은12
주동안매주 인간태반성성선자극호르몬(human chorionic gonadotropin, hCG; Daesung Microbiological, Uiwang, Korea)
을체중1 g
당
1 IU
씩복강에주사하여성성숙을유도하였다.
암컷체중이초기체중의
20%
이상증가하였을때생식공에 실리콘관 을삽입하여난을채취하였으며난경이900 μm
이상으로증 가하였을때17α-hydroxyprogesterone (OHP; Sigma-Aldrich Co., St. Louis, MO, USA) 10 mg/fish
와17, 20β-dihydroxy- 4-pregnen-3-one (DHP; Sigma-Aldrich Co., St. Louis, MO,
USA) 10 mg/fish
을복강에주사하여최종성숙및배란을유도하였다
.
수컷의경우복부압박으로채정하여수정에이용하 였다.
최종적으로수정란생산을위하여약20°C
해수의배란유도수조에성숙한암컷과수컷을각
1
마리씩합사한뒤23°C
로서서히수온을높여자연산란및수정을유도하였다
.
그결과2018
년12
월27
일(
오전9
시경)
에집란조에모인수정란을확인할수있었고
,
부상된수정란은약23°C
의레이스형부화수조에이동시켜부화시까지관리하여약
48
시간후에90%
이상부화하였다
. 자어 사육
부화된자어는부화후
6
일까지500 L
원통형수조에서먹이공급없이
25 μm
와10 μm
의카트리지필터로500
마리수준 으로수용하였으며,
부화후7
일째의15
시부터첫먹이공급을 실시하였다.
먹이는곱상어알을기초로하여대두박,
곤쟁이,
어분,
비타민등으로슬러리형사료로제작하여공급하였으며(Kim et al., 2014),
하루5
회(09:00, 11:00, 13:00, 15:00, 17:00)
각수조당6 mL
의사료를공급하였다.
부화후7
일부터50
일 까지주수량과사료점도는약1.0 L/min, 100 mPa·s
이었다. 샘플링 및 조직학적 분석
부화자어의소화기관발달을조직학적으로관찰하기위하여 부화직후부터부화후
7
일까지는매일,
그리고부화후10
일, 15
일, 30
일, 50
일째에20
마리이상샘플링하였다.
채집된개 체는마취제(MS-222, Ethyl 3-aminobenzoate methanesulfo- nate; Sigma-Aldrich Co., St. Louis, MO, USA) 200 ppm
으 로마취한후실체현미경(SteREO Discovery. V20; Carl Zeiss, Oberkochen, Germany)
에 연결된 디지털카메라(AxioCam MRc5; Carl Zeiss, Oberkochen, Germany)
로촬영한뒤즉시Bouin’s
용액에24
시간고정하였다.
각샘플일에20
마리씩촬 영된자어는Image analysis software (ZEN 2012 blue edition;
Carl Zeiss, Oberkochen, Germany)
를 이용하여 전장(total
length, TL)
을측정하였고,
고정된자어는파라핀포매법에따 라파라핀블록을제작한후마이크로톰(RM2255; Leica, Wet-
zlar, Germany)
을이용하여4-6 μm
두께로소화기관과평행하 게횡단으로 연속절편하였다.
이후슬라이드글라스에절편을신민규
ㆍ
김신권ㆍ
이배익ㆍ
최윤희ㆍ
유용운300
부착한뒤
Crossmon’s Trichrome
염색하여관찰하였다.
일부시료는식도와장의배상세포발달을확인하기위해
ab-PAS
염색을실시하였다
.
이후슬라이드커버로봉입한뒤광학현미경(DM6000B; Leica, Wetzlar, Germany)
에연결된디지털카메 라(DFC450 C; Leica, Wetzlar, Germany)
를이용하여소화경 로의구강인두,
식도,
장과소화를돕는분비기관인간,
담낭,
췌장의발달변화를중점적으로관찰하였다.
결 과
내부영양물질 흡수
A. japonica
자어의부화직후전장은3.31-3.79 mm (
평균3.61±0.14, n=20)
였으며,
부화후50
일에16.41-19.49 mm (
평균17.86±0.82, n=20)
까지성장하였다.
부화후0-50
일자 어의전장과부화일수의관계식은y=0.255x+4.996 (r²=0.981)
이었다
(Fig. 1).
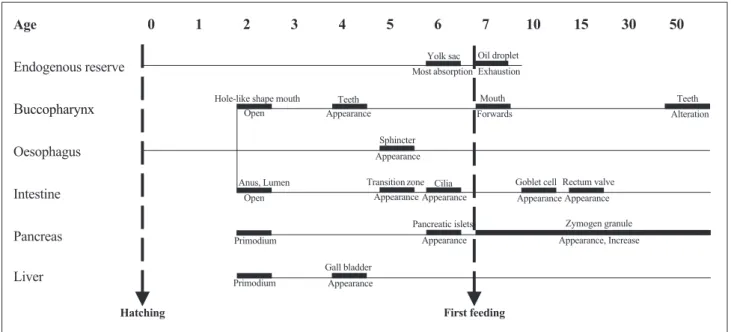
뱀장어자어는발달함에따라내부영양물질과소화기관에큰변화가나타났다
(Fig. 2).
부화시자어의난황(yolk sac, YS)
은전체체면적의2/3
이상차지하였으며,
유구(oil droplet, OD)
는몸의앞부분에한개존재하였다(Fig. 2A).
내부영양물질은앞부분부터흡수되기시작하였고부화후
1
일 에는앞쪽난황이흡수되어난황합포체층(yolk syncytial layer, YSL)
만남은채유구는뒤로이동하였다(Fig. 2B).
부화후6
일 에는대부분의난황이흡수되었으며장밑부분을따라소량의 난황과난황합포체층이관찰되었다(Fig. 2G).
부화후7
일에는 유구가대부분흡수된채난황합포체층만심장(heart, H)
뒷부 분에소량남아있었다(Fig. 2H).
부화후10
일에는난황합포체 층이모두흡수되어관찰되지않았다(Fig. 2I).
구강인두 발달
뱀장어자어는부화직후입이열려있지않았으나
(Fig. 3A),
부화후
2
일에인두강이넓어지며관모양의작은입이열린것 이확인되었다(Fig. 3B).
부화후3
일에는아래턱이분화되었으 며위턱과아래턱끝이융기되었다(Fig. 3C).
부화후4
일에는 양턱끝의융기된부분에이빨이관찰되기시작하였으며(Fig.
3D),
부화후5
일에는윗니가커지고더욱날카로워졌으며입천장에는연골조직이관찰되었다
(Fig. 3E).
첫섭식을시작하 는부화후7
일에는위턱보다아래턱이빠르게발달하여아래 턱이길어졌으며입이앞을향하게되었다(Fig. 3G).
부화후10
일에는아래쪽을향하던윗니가전방을향하였고
(Fig. 3G),
부화후
30
일에는위턱뼈가발달하고oral valve
가관찰되기시 작하였다(Fig. 3H).
부화후50
일에는이빨의크기가줄어듦과 동시에표피가발달하여외부형태관찰시이빨의일부만관찰 되었다(Fig. 2L, 3I).
식도 발달
부화 직후 자어의 식도
(oesophagus, OE)
는 단층입방상피(simple cuboidal epithelium)
로 이어진 얇은 관이었다(Fig.
4A).
부화후5
일에상부식도조임근(upper esophageal sphinc- ter)
이점막층(mucosa, M),
점막하층(submucosa, SM),
근육외 층(muscularis externa, ME),
바깥막(adventitia, A) 4
층구조로 발달된것을확인할수있었으며,
식도전체에서얇은근육외층 이관찰되었다(Fig. 4B).
부화후7
일에상부식도조임근은부 화후5
일에비해더넓게분포하였으며상피층은대부분단층 편평세포(simple squamous epithelium)
로구성되어있었지만 일부비각질중층편평상피(nonkeratinized stratified squamous epithelium)
가관찰되기시작하였다(Fig. 4C).
부화후30
일에 는상부식도조임근의범위가더넓어지고두꺼워졌고,
식도점 막상피중조임근의점막상피만비각질중층편평상피로변한 것을확인할수있었다(Fig. 4D).
장 발달
부화직후자어의장
(intestine, I)
은식도와이어진큰내강과 아직열리지않은관으로존재하였다(Fig. 5A).
부화후1
일까 지장내강과항문(anus, AN)
은열리지않았으나(Fig. 5B),
부 화후2
일에장내강과항문이열린것을확인할수있었다(Fig.
5C).
부화후5
일의장조직은단층원주상피세포(simple co- lumnar epithelium)
로구성되어있었으며미세융모(microvilli)
가표면에발달되어있었다(Fig. 5D).
부화후6
일에는후장의점막 상피에섬모
(cilia, C)
가 처음으로관찰되기 시작하였으며
,
부화후7
일에는장내강이부화6
일에비해넓어졌고중 장(middle intestine, MI)
에서도섬모가관찰되었으나주로후 장(posterior intestine, PI)
에 분포하고 있었다(Fig. 5E).
부화 후15
일에는부화후7
일에비해장내강이더욱넓어졌고중 장과후장상피에서보다많은섬모가관찰되었다.
또한,
중장 과후장사이에직장판막(rectum valve, RV)
이발달하였고직 장에는점막주름(mucosal fold, MF)
이발달되어있었다(Fig.
Fig. 1. Growth in total length of Anguilla japonica larvae from hatching to 50 days after hatching.
y=0.255x+4.996 r
²=0981 0
5 10 15 20
0 10 20 30 40 50
Total length (mm)
Days after hatching
뱀장어 자어의 소화기관 발달 특성
301
5F).
부화후30
일에는식도와전장(anterior intestine, AI)
의 경계(transition zone)
에점막주름이발달하였으며직선형장(straight gut)
이유지되었다(Fig. 5G).
배상세포 분포
부화후
5
일부터자어의식도와장내강상피에서ab-PAS
에양성반응을보이는산성점액단백질
(acidic mucosubstance)
이 관찰되었지만점액을형성하고분비하는배상세포(goblet cell, GC)
는첫섭식시까지관찰되지않았다.
부화후10
일에식도 점막상피의산성점액단백질은비교적증가하였으나(Fig. 6A),
배상세포는부화 후50
일까지확인되지 않았다(Fig. 6B).
한 편장에서는부화후10
일에처음으로배상세포가관찰되었고(Fig. 6C),
부화후50
일까지그수가증가한것을확인할수있 었다(Fig. 6D).
간과 췌장 발달
뱀장어자어의간
(liver, L)
과췌장(pancreas, P)
은부화후2
일에유구위소화관근처에처음출현하였다(Fig. 7A).
이후 자어가성장하며식도가길어짐에따라점차몸의중간부분 으로이동하였고(Fig. 2),
부화후4
일에는간과췌장주변에서 담낭(gall bladder, GB)
이출현하였다(Fig. 7B).
부화후5
일에 는간과췌장이조직학적으로구분되기시작하였으며,
다면체 의간세포에서는지질축적의흔적으로보이는공포가관찰되 었고소엽구분없이실질이길게배열되어있었다.
또한,
담관 Fig. 2. Comparison with morphological (upper) and histological (lower) development of Anguilla japonica larvae. Abbreviations: DAH, days after hatching; OE, oesophagus; OD, oil droplet; YS, yolk sac; YSL, yolk syncytial layer; L, liver; P, pancreas; I, intestine; AN, anus;GB, gall bladder. Scale bars indicate 500 μm.
OD YS YSL OD YS OD
OE OE OE I
YSL YS
I
YSL YS AN I
OD YS OE
L and P OE L P YS I
OD GB AN
OD YS OE
L P
I AN YSL OE L GB I
P AN
L and P
OE L
P GB I
I L GB OE
P
AN OE L GB I
P
GB I OE L
P GB
(D)
(A) (B) (C)
(E) (F)
(G) (H) (I)
(J) (K) (L)
OD YS
OE OE
YSL OD YS YSL OD
OE I
YS
YS I AN YSL
OE OD
I
YS ODOE YS I
AN
YSL L I AN OE L I
OE L I AN
OD OE
L AN
GB
GB I YS OE
L I
OE OE L I
0 DAH 1 DAH 2 DAH
3 DAH
6 DAH
4 DAH 5 DAH
7 DAH 10 DAH
30 DAH 50 DAH
15 DAH
(A) (B) (C)
(D) (E) (F)
(G) (H) (I)
OE
H
H
OE OE
H BC
T BC OE
H H
T
BC BC OE
T
50 μm 50 μm 50 μm
100 μm
100 μm 100 μm 200 μm
BC BC
BC T T
T
OV OV
T EP
EP
50 μm 50 μm
0 DAH 2 DAH 3 DAH
4 DAH 5 DAH 7 DAH
30 DAH 50 DAH
10 DAH
BC
50 μm 50 μm
50 μm
OE H
OE
OE
H
YSL
SM M ME A OD
H
(A) (B)
(C)
100 μm
OE H
(D)
BC
0 DAH 5 DAH
7 DAH 30 DAH
100 μm 100 μm 100 μm
100 μm 100 μm
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
50 μm 50 μm 50 μm
50 μm
50 μm
100 μm
50 μm 50 μm
50 μm 50 μm
H YSL
OE I
YS OD
I YS YSL
AN AN
YS YSL I UB
YSL P
YS
AI MI
YS
PI
YSL AN
UB
P
ZG
AI MI PI
AN C UB
C
AI MI
C
C P
ZG ZG
MF AI OE
ZG
P MI
RV
C
AN
AN MF
C C
PI UB
UB C
C PI
MF L
P P
GB OE (A)
C C C
(B) (C)
(D)
(E)
(F)
(G)
MF
C C
C C C
0 DAH 1 DAH 2 DAH
5 DAH
7 DAH
15 DAH
30 DAH
Anterior
Anterior
Anterior
Anterior
Middle
Middle
Middle
Middle
Posterior
Posterior
Posterior
Posterior
Oesophagus Intestine
(B)
50 μm 50 μm 50 μm
50 μm 50 μm
50 μm
50 μm 50 μm
50 μm 50 μm 50 μm
50 μm
(A)
(C)
(D)
OE OE OE
OE OE
OE L
L
P P
P ZG
ZG P GB ZG
ZG GC
GC
GC
GC
GC GC
GC
GC GC
GC GC
GC GC GC GC
GC GC
10 DAH
10 DAH 50 DAH
50 DAH
I OD
OE
YS
L and P OE AI
L and P GB
L GB
OE BD AI GB EP ZG
P
50 μm
50 μm 50 μm
50 μm
(A) (B)
(C) (D)
(E) (F)
AI IL
L EP
ZG
2 DAH 4 DAH
5 DAH 7 DAH
15 DAH 50 DAH
신민규
ㆍ
김신권ㆍ
이배익ㆍ
최윤희ㆍ
유용운302
(bile duct, BD)
은장의앞부분에연결되어있는것을확인할수 있었다(Fig. 7C).
부화후7
일이후에는자어의성장과함께간 의크기,
간세포의수그리고담낭의크기가증가하였으나,
부화 후10
일이후에는간세포내지질축척이관찰되지않았다(Fig.
7D-7F).
췌장에서는부화후6
일에처음으로내분비기관인췌장섬
(islets of Langerhans)
이구분되기시작하였으며,
부화후7
일부터외분비췌장에서효소원과립(zymogen granules, ZG)
이관찰되었다(Fig. 7D).
이후성장함에따라췌장의크기,
세 포수그리고효소원과립의분비가증가하였으며췌장내다량 의효소원과립으로추정되는호산성물질의축적이관찰되었다(Fig. 7D-7F).
고 찰
이번연구에서 관찰한뱀장어자어의부화후
50
일 동안의 일간평균성장률은0.255 mm/day
로나타났다. Tanaka et al.
(2001)
가앞서보고한인공산50
일까지의성장률0.234 mm/
day
와유사한결과를나타냈지만자연산자어에서추정된성장 률0.5 mm/day
의절반수준이었다(Ishikawa et al., 2001; Oka- mura et al., 2014).
이는상술한Hsu et al. (2015)
의보고와같 이현재의슬러리형인공사료가뱀장어자어의장내소화및흡수에적합하지않기때문에자연산에비해성장이느린것으로 추측되며
,
따라서적합한사료의개발은인공종자의성장과생 존율을높이기위해시급히해결해야할문제이다.
자어의혼합섭식기간은첫외부먹이의섭식부터내부영양이 흡수완료되기까지의기간을의미하며
(Fhyn, 1989),
이기간에 는내부영양과외부영양을동시에소비하며성장과생존에필 요한에너지를생산한다.
혼합섭식기간이짧은것으로알려진 어종들은돌돔(Oplegnathus fasciatus) 0
일(Shan et al., 2008), gilthead sea bream (Sparus aurata) 0
일(Yúfera et al., 1993),
넙치(Paralichthys olivaceus) 0-1
일(Dou et al., 2005),
줄삼치(Sarda orientalis) 2
일(Kaji et al., 2002)
로보고된바있으며,
대 구(Gadus macrocephalus) 9
일(Shin et al., 2019),
명태(Gadus chalcogrammus) 4-6
일(Yokota et al., 2016),
까나리(Ammo- dytes personatus) 10
일(Yamashita and Aoyama, 1985),
자주 복(Takifugu rubripes) 8
일(Jiang et al., 2002)
등은혼합섭식기 간이긴것으로알려져있다.
이번연구에서부화자어의초기 생존및발달에중요한내부영양물질인난황과유구는각각부 화후5
일과7
일에완전히흡수되었고,
이후7
일령부화자어는 바로먹이섭취를시작해혼합섭식기간이거의존재하지않았 다.
이는Kurokawa et al. (1995)
가인공산뱀장어자어에서관 찰한결과와동일하였으나, Kurokawa and Pedersen (2003)
는 Fig. 3. Longitudinal histological sections of the buccopharynx in Anguilla japonica larvae. Abbreviations: DAH, days after hatching; OE, oesophagus; H, heart; BC, buccal cavity; T, teeth; OV, oral valve; EP, epithelium.OD YS YSL OD YS OD
OE OE OE I
YSL YS
I
YSL YS AN I
OD YS OE
L and P OE L P YS I
OD GB AN
OD YS OE
L P
I AN YSL OE L GB I
P AN
L and P
OE L
P GB I
I L GB OE
P
AN OE L GB I
P
GB I L OE
P GB
(D)
(A) (B) (C)
(E) (F)
(G) (H) (I)
(J) (K) (L)
OD YS
OE OE
YSL OD YS YSL OD
OE I
YS
YS
AN I YSL
OE OD
I
YS ODOE YS I
AN
YSL L I AN OE L I
OE L I AN
OD OE
L AN
GB
GB I YS OE
L I
OE OE L I
0 DAH 1 DAH 2 DAH
3 DAH
6 DAH
4 DAH 5 DAH
7 DAH 10 DAH
30 DAH 50 DAH
15 DAH
(A) (B) (C)
(D) (E) (F)
(G) (H) (I)
OE
H
H
OE OE
H BC
T BC OE
H H
T
BC BC OE
T
50 μm 50 μm 50 μm
100 μm
100 μm 100 μm 200 μm
BC BC
BC T T
T
OV OV
T EP
EP
50 μm 50 μm
0 DAH 2 DAH 3 DAH
4 DAH 5 DAH 7 DAH
30 DAH 50 DAH
10 DAH
BC
50 μm 50 μm
50 μm
OE H
OE
OE
H
YSL
SM M ME A OD
H
(A) (B)
(C)
100 μm
OE H
(D)
BC
0 DAH 5 DAH
7 DAH 30 DAH
100 μm 100 μm 100 μm
100 μm 100 μm
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
50 μm 50 μm 50 μm
50 μm
50 μm
100 μm
50 μm 50 μm
50 μm 50 μm
H YSL
OE I
YS OD
I YS YSL
AN AN
YS YSL I UB
YSL P
YS
AI MI
YS
PI
YSL AN
UB
P
ZG
AI MI PI
AN C UB
C
AI MI
C
C P
ZG ZG
MF AI OE
ZG
P MI
RV
C
AN
AN MF
C C
PI UB
UB C
C PI
MF L
P P
GB OE (A)
C C C
(B) (C)
(D)
(E)
(F)
(G)
MF
C C
C C C
0 DAH 1 DAH 2 DAH
5 DAH
7 DAH
15 DAH
30 DAH
Anterior
Anterior
Anterior
Anterior
Middle
Middle
Middle
Middle
Posterior
Posterior
Posterior
Posterior
Oesophagus Intestine
(B)
50 μm 50 μm 50 μm
50 μm 50 μm
50 μm
50 μm 50 μm
50 μm 50 μm 50 μm
50 μm
(A)
(C)
(D)
OE OE OE
OE OE
OE L
L
P P
P ZG
ZG P GB ZG
ZG GC
GC
GC
GC
GC GC
GC
GC GC
GC GC
GC GC GC GC
GC GC
10 DAH
10 DAH 50 DAH
50 DAH
I OD
OE
YS
L and P OE AI
L and P GB
L GB
OE BD AI GB EP ZG
P
L AI L
AI ZG
ZG GB
GB
50 μm 50 μm
50 μm 50 μm
50 μm
100 μm
(A) (B)
(C) (D)
(E) (F)
AI IL
L
IL EP
EP
EP
EP ZG
ZG ZG
2 DAH 4 DAH
5 DAH 7 DAH
15 DAH 50 DAH
뱀장어 자어의 소화기관 발달 특성
303
부화후
8
일에첫섭식을시작하며난황과유구는각각부화후7
일과12
일에완전히흡수된다고보고하였다.
이러한결과를 바탕으로,
인공생산된뱀장어는사육환경및자어의영양상태 에따라다소차이가있을수있으나혼합섭식기간이없거나짧 은그룹에속하는것으로생각된다.
어류자어에있어짧은혼합 섭식기간은기아발생에취약하며첫섭식실패시성장과생존 율감소로이어진다(Yokota et al., 2016; Shin et al., 2019).
특 히,
뱀장어자어사육시아직적절한사육시스템과먹이공급 방법이확립되어있지않아많은수의개체가기아로폐사하는 문제점이있어(Okamura et al., 2019),
적정시기의첫먹이공 급및기아발생방지는초기성장이느리고생존율이낮은인공 산뱀장어의사육기술개선에매우중요한요인으로판단된다.
이번연구에서뱀장어자어는부화후2
일에입과항문이열 렸고첫섭식시기에맞춰다수의바늘모양이빨이출현하였으 며윗앞니가전방약45°
의갈고리모양으로변했다.
이는크고 부드러운입자를잡는데적합한형태라는것이보고된바있으 며(Westerberg, 1990),
태평양에서채집된뱀장어자어의유전 자동정결과다양한미생물과동물성플랑크톤들이확인되어(Miller et al., 2019; Tsukamoto and Miller, 2021),
이들이분해되어응집된산물인부드러운
marine snow
를주먹이로이용할가능성이시사되었다
.
또한,
많은해산어자어의경우순간적으로먹이를물과함께흡입하여포획하는섭식행동을보이는반 면
(Hernández, 2000),
뱀장어자어는슬러리형의먹이공급시 사료에머리를박고입을벌린채앞으로유영하며먹이를섭취 하는행동특성이나타났다.
따라서뱀장어pre-leptocephalus
사육시돌출된앞니를박아넣고앞으로전진하며섭취할수 있는부드러운형태의먹이가적합할것으로판단된다.
반면, leptocephalus
단계의부화후50
일에는돌출된앞니의감소와 외피의발달로외부이빨이점차보이지않게변하였다.
이러한 구강구조의변화는자연환경에서이들의먹이원과섭식방법 의변화를반영하는현상으로생각되며, leptocephalus
의치아 구조는젤라틴형태의물체를잡는데적합하다는보고로미루 어보아(Westerberg, 1990), leptocephalus
단계에서도지속적 으로공급하는슬러리형태의사료를섭식기관의발달단계를 고려하여적합한형태로개선해야할것이다.
뱀장어자어의식도는성장함에따라길게발달하여일반적인 자어에비해긴구조가관찰되었다
.
이와함께,
상부식도조임근 은부화후5
일에점막층,
점막하층,
근육외층,
바깥막구조를갖 췄으며성장함에따라그부위가넓어지고근육외층이증가하 였다.
인간의경우상부식도조임근은주로식도로공기가들어 가는것을방지하며하부식도조임근은위안의산성내용물이 역류하지못하도록방지하는역할을하는것으로알려져있지 Fig. 4. Longitudinal histological sections of the oesophagus in Anguilla japonica larvae. Abbreviations: DAH, days after hatching; OE, oesophagus; H, heart; BC, buccal cavity; M, mucosa; SM, submucosa; ME, muscularis externa; A, adventitia; YSL, yolk syncytial layer.OD YS YSL OD YS OD
OE OE OE I
YSL YS
I
YSL YS AN I
OD YS OE
L and P OE L P YS I
OD GB AN
OD YS OE
L P
I AN YSL OE L GB I
P AN
L and P
OE L
P GB I
I L GB OE
P
AN OE L GB I
P
GB I L OE
P GB
(D)
(A) (B) (C)
(E) (F)
(G) (H) (I)
(J) (K) (L)
OD YS
OE OE
YSL OD YS YSL OD
OE I
YS
YS I AN YSL
OE OD
I
YS ODOE YS I
AN
YSL L I AN OE L I
OE L I AN
OD OE
L AN
GB
GB I YS OE
L I
OE OE L I
0 DAH 1 DAH 2 DAH
3 DAH
6 DAH
4 DAH 5 DAH
7 DAH 10 DAH
30 DAH 50 DAH
15 DAH
(A) (B) (C)
(D) (E) (F)
(G) (H) (I)
OE
H
H
OE OE
H BC
T BC OE
H H
T
BC BC OE
T
50 μm 50 μm 50 μm
100 μm
100 μm 100 μm 200 μm
BC BC
BC T T
T
OV OV
T EP
EP
50 μm 50 μm
0 DAH 2 DAH 3 DAH
4 DAH 5 DAH 7 DAH
30 DAH 50 DAH
10 DAH
BC
50 μm 50 μm
50 μm
OE H
OE
OE
H
YSL
SM M ME A OD
H
(A) (B)
(C)
100 μm
OE H
(D)
BC
0 DAH 5 DAH
7 DAH 30 DAH
100 μm 100 μm 100 μm
100 μm 100 μm
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
Posterior Middle
Anterior
50 μm 50 μm 50 μm
50 μm
50 μm
100 μm
50 μm 50 μm
50 μm 50 μm
H YSL
OE I
YS OD
I YS YSL
AN AN
YS YSL I UB
YSL P
YS
AI MI
YS
PI
YSL AN
UB
P
ZG
AI MI PI
AN C UB
C
AI MI
C
C P
ZG ZG
MF AI OE
ZG
P MI
RV
C
AN
AN MF
C C
PI UB
UB C
C PI
MF L
P P
GB OE (A)
C C C
(B) (C)
(D)
(E)
(F)
(G)
MF
C C
C C C
0 DAH 1 DAH 2 DAH
5 DAH
7 DAH
15 DAH
30 DAH
Anterior
Anterior
Anterior
Anterior
Middle
Middle
Middle
Middle
Posterior
Posterior
Posterior
Posterior
Oesophagus Intestine
(B)
50 μm 50 μm 50 μm
50 μm 50 μm
50 μm
50 μm 50 μm
50 μm 50 μm 50 μm
50 μm
(A)
(C)
(D)
OE OE OE
OE OE
OE L
L
P P
P ZG
ZG P GB ZG
ZG GC
GC
GC
GC
GC GC
GC
GC GC
GC GC
GC GC GC GC
GC GC