Korean J Vet Res(2017) 57(2) : 89~95 https://doi.org/10.14405/kjvr.2017.57.2.89
89
<Original Article>
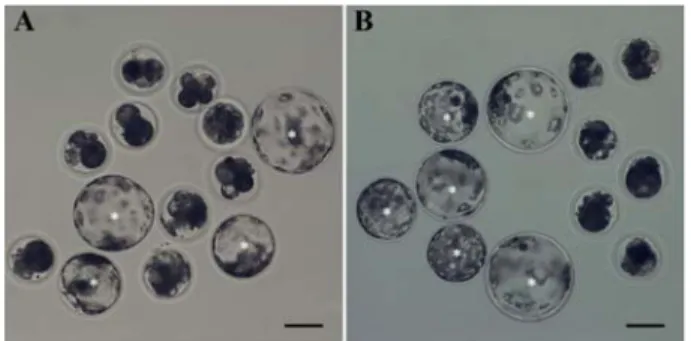
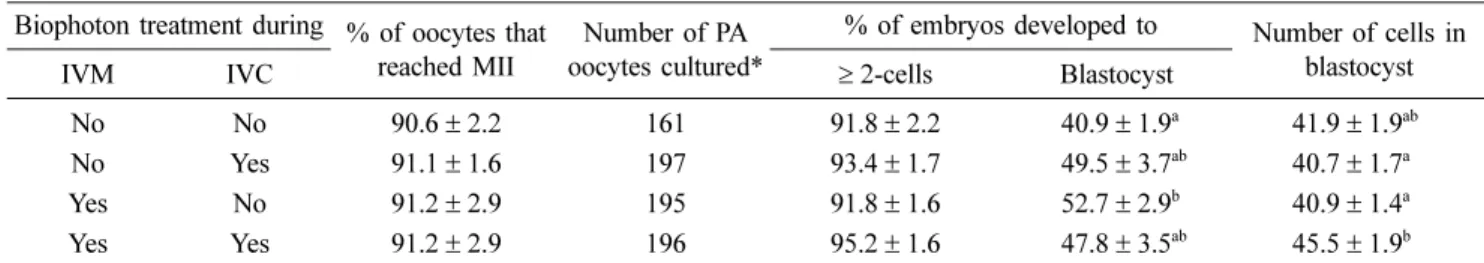
Oocyte maturation under a biophoton generator improves preimplantation development of pig embryos derived by parthenogenesis
and somatic cell nuclear transfer
Joohyeong Lee
1,2, Hyeji Shin
1, Wonyou Lee
3, Seung Tae Lee
4, Geun-Shik Lee
1,2, Sang-Hwan Hyun
5, Eunsong Lee
1,2,*
1
Laboratory of Theriogenology, College of Veterinary Medicine,
4Department of Animal Life Science and Division of Applied Animal Science, College of Animal Life Science, and
2Institute of Veterinary Science, Kangwon National University, Chuncheon 24341, Korea
3
Biolight Corporation, Seoul 05839, Korea
5