RESEARCH ARTICLE
Received: January 3, 2017, Revised: January 23, 2017, Accepted: January 23, 2017 ISSN 1598-4478 (Print) / ISSN 2233-7679 (Online)
†
Correspondence to: Sang-im Lee
Department of Dental Hygiene, College of Health Science, Dankook University, 119 Dandae-ro, Dongnam-gu, Cheonan 31116, Korea Tel: +82-41-550-1492, Fax: +82-41-559-7934, E-mail: [email protected]
Copyright © 2017 by Journal of Dental Hygiene Science
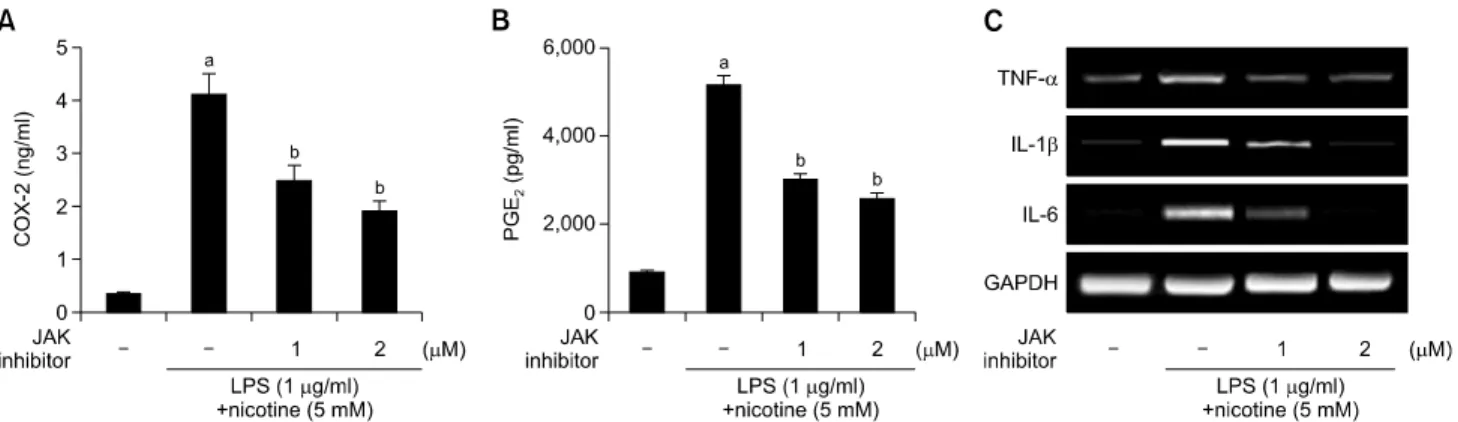
조골세포에서 Porphyromonas gingivalis
Lipopolysaccharide와 니코틴에 의한 염증에 대한 JAK/STAT Pathway의 역할
한양금ㆍ이인수
1
ㆍ이상임2†
대전보건대학교 치위생과,
1
한남대학교 생명·나노과학대학 생명시스템과학과,2
단국대학교 보건과학대학 치위생학과JAK/STAT Pathway Modulates on Porphyromonas gingivalis Lipopolysaccharide- and
Nicotine-Induced Inflammation in Osteoblasts
Yang-keum Han, In Soo Lee 1 , and Sang-im Lee 2†
Department of Dental Hygiene, Daejeon Health Science College, Daejeon 34504,
1
Department of Biological Sciences and Biotechnology, College of Life Science and Nano Technology, Hannam University, Daejeon 34430,
2