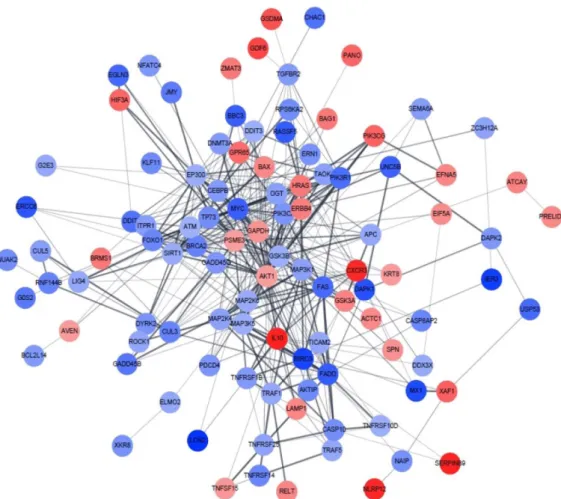
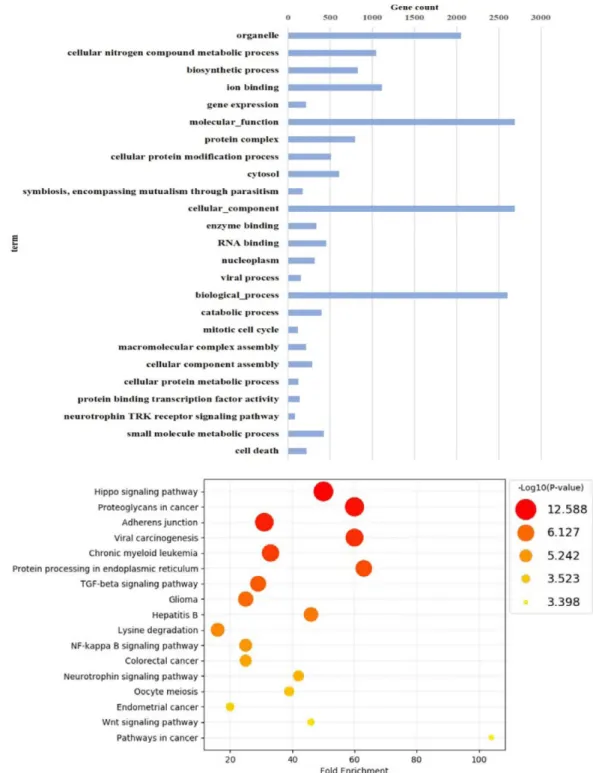
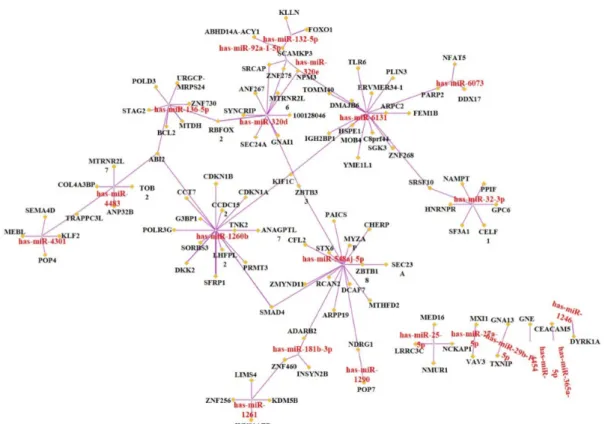
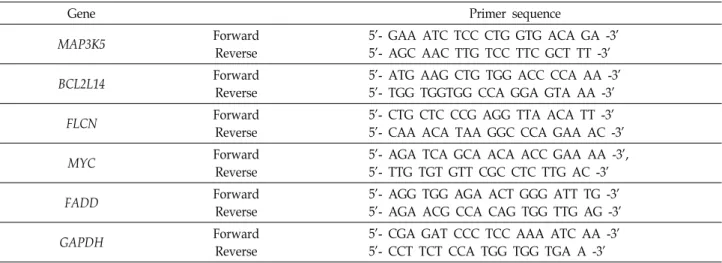
Effects of Sasa quelpaertensis Extract on mRNA and microRNA Profiles of SNU-16 Human Gastric Cancer Cells
Mi Gyeong Jang
1,2, Hee Chul Ko
2and Se-Jae Kim
1,2*
1
Department of Biology, Jeju National University, Jeju 63243, Korea
2
전체 글
1
2
수치

관련 문서
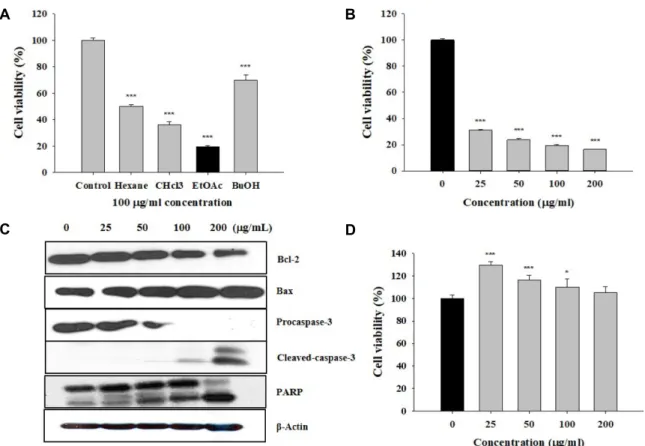
Here, we found that TAMR-MCF-7 cells had undergone EMT, evidenced by mesenchymal-like cell shape, down-regulation of the basal E-cadherin expression
Inhibitory effects of classified methanol extracts of Smilax china L on the COX-2 and iNOS expression of human colorectal cancer cell lines... Inhibitory
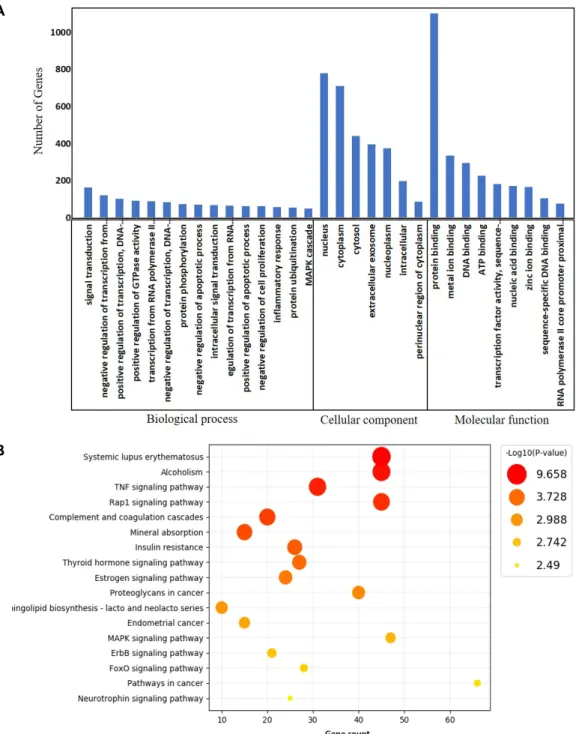
Often found connected to other molecules on the outsides of cells --- cellular recognition, cell signaling, cell
4 > Effects of Taro on COX-2 expression and iNOS expression(hot water) in human thyroid cancer cells. The cells were pretreated for 48hours with either
These results suggest that the bilobalide inhibits the cell proliferation and induces the apoptotic cell death in FaDu human pharyngeal squamous cell carcinoma via both
In the present study we investigated the proliferation effect of human oral cancer KB cell treated with pulsatilla koreana extract.. We analyzed the effects of this
Taken together, these results suggested that latex containing the ficin inhibited the cell growth and induced apoptosis by caspase and Bcl-2 family signaling pathway in
Comparison of cell attachment on the bone grafting materials synthesized as the defined mixing ratio of Hydroxyapatite (HA) and β-tricalcium phosphate (β-TCP) by DAPI staining