서 론
뇌허혈 후 발생되는 c y c l o o x y g e n a s e ( C O X )의 활성화 에 의해 arachidonic acid를 경유한 p r o s t a g l a n d i n ( P G )과 t h r o m b o x a n e ( T X )의 대사가 촉진된다. COX에 는 type 1(COX-1)과 type 2(COX-2) 두 가지 종류가 있다. COX-1은 다양한 조직에서 발현되는데 혈소판, 위장
관 점막 및 신장 등에서 관찰할 수 있고,1 , 2C O X - 2는 외부 자극에 의해 유도가 가능한 효소로 주로 백혈구 및 뇌조직 에서 발현된다.3C O X - 2의 활성화는 염증반응에 의한 조직 손상의 중요한 기전으로 뇌허혈성 손상시 동반되는 염증반 응과 관련이 있다.4 - 6 C O X - 2의 활성화는 다양한 e i c o s a n o i d s와 유리기(free radical)를 생산하여 뇌허혈 시에 중요한 이차적 손상기전으로 작용하는데 허혈과 재관 류 시에 뇌혈관세포에서 그리고 뇌허혈에 의한 경색부위 주 변의 신경조직에서 C O X - 2의 발현이 관찰된다.7 - 8
최근 C O X의 활성화를 억제하여 뇌허혈에 의한 손상을 억제하고자하는 연구들이 진행되고 있다.9 - 1 1 비스테로이드 성 항염증약물들(non-steroidal anti-inflammatory drugs, 이하 N S A I D s로 함)은 C O X - 2의 활성을 억제하므 로 허혈성 신경세포손상 억제에 대한 연구에 사용된다.1 2 - 1 4 그러나 C O X - 2의 활성화에 의한 이차적 신경조직의 손상에
일과성 전뇌허혈 후 뇌조직내 Polyamine 함량 변동과 신경세포 손상에 대한 I b u p r o f e n의 효과
계명대학교 의과대학 신경과학교실, 약리학교실*
이승환환이성룡*환임정근환이 형환이상도
Effect of Ibuprofen on the Changes of Polyamine Level and Neuronal Cell Damage after Transient Global Ischemia in Gerbil
Seung-Hwan Lee, M.D., Seong-Ryong Lee, M.D.,* Jeong-Geun Lim, M.D., Hyung Lee, M.D., Sang-Doe Yi, M.D.
Departments of Neurology and Pharmacology,* School of Medicine, Keimyung University
Background : In brain ischemia, increased arachidonic acid metabolism can play important roles in neuronal dam- age. Ibuprofen was reported to have a protective role against neuronal damage in focal brain ischemia and reperfusion.
The present study was designed to investigate whether ibuprofen can inhibit the global ischemia-induced neuronal dam- age and changes of polyamine (PA) level which is known to related to the neuronal damage, breakdown of blood brain barrier, and brain edema. Methods : Male Mongolian gerbils were used in this study. Transient global ischemia was induced by occlusion of bilateral common carotid arteries for 3 min with microclips. Ibuprofen was administered imme- diately after ischemia. The animals were sacrificed one day after ischemia for PA measurement and sacrificed 5 days after ischemia for histological evaluation. Histological examination was performed by counting surviving neuronal cells in one mm of CA1 area in dorsal hippocampus. Results : Cerebral cortex and hippocampal putrescine(PU) levels in vehicle-treated ischemic group significantly increased comparing to sham-operated animals and the increase of PU was attenuated by ibuprofen administration (50 mg/kg). Hippocampal spermine level decreased significantly after ischemia.
Hippocampal neuronal cell damage in CA1 area was markedly observed in vehicle-treated animals compared to sham operated animals. Ibuprofen administration at the dose of 50 mg/kg significantly inhibited hippocampal CA1 neuronal damage compared to vehicle-treated animals. Conclusions : Ibuprofen attenuates PA response following transient glob- al ischemia and may have putative neuroprotective effect against neuronal damage induced by global ischemia.
J Korean Neurol Assoc 20(3):265~272, 2002
Key Words : Global ischemia, Polyamine, Ibuprofen, Neuronal damage, Hippocampus, Neuroprotection
Manuscript received December 19, 2001.
Accepted in final form March 13, 2002.
* Address for correspondence Sang-Doe Yi, M.D.
Department of Neurology,Keimyung University School of Medicine,
194 Dongsan-dong, Daegu, 700-310, Korea Tel : +8 2 - 5 3 - 2 5 0 - 7 8 3 2 Fax : +8 2 - 5 3 - 2 5 2 - 1 6 0 5 E-mail : s d l e e @ d s m c . o r . k r
관한 연구는 주로 국소 뇌허혈 모델을 사용했으며 전뇌허혈 모델을 통한 연구는 드물다. NSAIDs 중 특히 i b u p r o f e n 의 허혈성 신경세포 손상의 억제작용에 대한 연구는 드물 다. Ibuprofen이 신경 보호작용을 나타낼 수 있는 가능성 을 뒷받침할 수 있는 작용들을 나열해보면 첫째, 뇌혈관 수 축에 의한 혈류량 감소를 억제시키며,1 5 - 1 6둘째, COX의 활 성화를 억제하여 염증에 의한 신경조직 손상을 억제하며,9 셋째, glutamate에 의한 신경 독작용을 억제하는 작용1 7 등이 있다.
포유류에 존재하는 p o l y a m i n e ( P A )은 p u t r e s c i n e (PU), spermidine(SD) 및 spermine(SM) 등으로 구성 되며 P A은 거의 모든 살아있는 세포에 존재하면서 세포의 성장, 발육 및 분화 등에 관여한다.1 8 - 1 9내인성 P A은 중추 신경계에서 다양한 작용을 하는데 주로 신경전달체나 신경 조절물질로 작용한다.2 0 또한 뇌내 P A은 다양한 종류의 자 극들 즉 화학적, 물리적 및 대사성 자극 등에 의해 합성이 촉진되면 P A이 축적되어 신경세포 손상, 뇌부종 및 혈뇌장 벽(blood brain barrier)의 파괴 등을 일으킨다. PA은 세포내 칼슘 함량을 증가시키고,2 1 - 2 2흥분성 아미노산을 유 리시키는 작용을 나타낸다.2 3 Paschen 등2 4은 허혈과 P A 대사의 관계를 연구한 결과 특히 P U은 정상적으로 뇌에서 매우 낮은 함량으로 존재하는데 허혈에 의해 함량이 과다하 게 증가하면 신경세포에 매우 유해하다고 하였다. 따라서 PA 대사에 관련된 효소를 억제하여 P A의 과다한 축적을 억제하여 뇌를 보호하려는 연구들이 시도되고 있다.2 5 - 2 6
이에 본 연구에서는 gerbil 모델을 사용하여 일시적 전뇌 허혈을 유발시킨 후, NSAIDs의 하나인 i b u p r o f e n이 허 혈성 손상과 관련된 P A의 함량증가와 신경세포 손상을 억 제할 수 있는지 규명하고자 하였다.
대상과 방법 1. 실험동물
본 연구에서 체중 60~80 g의 웅성 Mongolian ger- b i l s (Meriones ungiculatus)을 같은 장소에서 동일한 사료로 사육하여 사용하였다.
2. 전뇌허혈 유발 수술 및 약물 투여
Chloral hydrate(Sigma Chemical Co., St.
Luise, MO, USA) 400 mg/kg을 복강 내에 투여하여 마취시킨 후 목 중앙에 2 cm 정도의 피부를 절개한 후 양 측 총경동맥을 주위의 미주신경과 정맥들로 부터 조심스럽 게 분리한 후 미세 클립( m i c r o c l i p )으로 3분간 결찰한 후 재관류를 실시하였다. 재관류는 육안으로 확인하였고 이후 피부를 봉합하였다. 저체온으로 인한 영향을 배제하기 위해 열판(CMA, Stockholm, Sweden)과 백열등을 이용하여 수술 및 수술 후 세 시간 동안 직장온도를 3 7±0 . 5℃로 유 지하였다. 본 실험에서 ibuprofen(Sigma Chemical Co., St. Luise, MO, USA)은 증류수에 녹인 후 사용하 였는데 투여용량을 각각 10, 25 및 50 mg/kg으로 하였 고 몸무게 100 g 당 0.2 ml가 되도록 농도를 맞추어 주
사하였다. 약물은 허혈 유발 직후에 복강내로 투여하였다.
허혈 대조군은 약물과 동일한 양의 증류수를 역시 허혈 유 발 직후에 복강내로 투여하였다.
3. 실험군
본 실험에는 5 종류의 군으로 나누었으며 아래와 같다.
(1) Sham 수술군: 마취 후 총경동맥을 노출시키고 허 혈 및 약물처치를 하지 않은 비허혈 대조군(PA 측정, n=5; 조직학적 관찰, n=6)
(2) Vehicle 투여군: 허혈 유발 수술 및 약물과 동일한 양의 증류수를 투여한 군(PA 측정, n=5; 조직학적 관찰, n = 6 )
(3) Ibuprofen 10 mg/kg 투여군: 허혈 유발 수술 및 10 mg/kg의 i b u p r o f e n을 투여한 군(PA 측정, n=4; 조 직학적 관찰, n=6)
(4) Ibuprofen 25 mg/kg 투여군: 허혈 유발 수술 및 25 mg/kg의 i b u p r o f e n을 투여한 군(PA 측정, n=4; 조 직학적 관찰, n=6)
(5) Ibuprofen 50 mg/kg 투여군: 허혈 유발 수술 및 50 mg/kg의 i b u p r o f e n을 투여한 군(PA 측정, n=5; 조 직학적 관찰, n=8)
4. PA 추출 및 측정
P A의 추출과정은 S p r a g g와 H u t c h i n g s의 방법2 7에 준 하여 실시하였다. 수술 2 4시간 후 실험동물을 희생시켜 두 개골에서 즉시 뇌를 꺼내어 해마와 대뇌피질을 즉시 분리한 후 Eppendorf 튜브에 담아 측정 시까지 - 7 0℃ 냉동상태 에 보관하였다. 얼음으로 냉각시킨 0.4 M perchloric acid 용액(2 mM disodium EDTA 및 4×1 0- 5 M의 1 , 8 - d i a m i n o o c t a n e을 함유)을 조직 무게의 1 0배에 해 당되는 부피로 첨가한 후 뇌조직을 h o m o g e n i z e r를 이용 해서 균질화시켰다. 원심분리(12,000 g, 4℃, 15 min) 후 상층액을 분리하여 진공 건조시킨 후 1 M의 s o d i u m b i c a r b o n a t e용액 100 μl를 넣어 용해시킨 후 300 μl의 4-fluoro-3-nitrobenzotrifluride(FNBT) 시약을 넣어 6 0℃에서 20 분간 반응시키고 40 μl의 1 M histidine을 첨가하여 5분간 추가반응 시킨 후에 얼음으로 냉각시켰다.
그후 2 ml의 2 - m e t h y l b u t a n e으로 2회 P A을 추출해내 었다. 5분간 원심분리 후 유기 용매층을 분리하여 다른 시 험 관 에 옮 겨 질 소 가 스 하 에 서 건 조 시 킨 후 methanol(500 μl )로 재용해 시켰다. 재용해시킨 용액 20 μl를 Hamilton 주사기를 사용해 크로마토그라피에 주 사하여 분석하였다. 크로마토그라피에 이용된 이동상 (mobile phase)은 a c e t o n i t r i l e과 증류수의 비율을 8 6 대 1 4 (부피비)로 하였고 분석전 초음파를 이용해 1 0분간 가스제거( d e g a s s i n g )를 시킨 후 사용하였고, 이동상의 유 속은 분당 1 mL로 하였고, column은 R a i n i n사의 O D S C18 column [250 mm(길이)×4 mm(내경) ]을 사용하 였다. 분석은 UV(ultraviolet) 검출기로 242 nm에서 실 시하였다.
5. 조직학적 검사
실험동물을 허혈 5일 경과 후 chloral hydrate로 깊이 마취시키고 흉곽을 연 후 좌심실을 통해 헤파린(2 IU/ml) 이 처리된 phosphate-buffered saline(PBS, pH 7.2) 과 10% formalin 고정액으로 관류시켰다. 두개골에서 뇌 를 꺼낸 후 10% formalin에 보관하여 추가 고정시켰다 (24~48 시간). 뇌를 파라핀으로 포매시키고 r o t a r y m i c r o t o m e을 이용하여 6 μm 두께의 관상절편을 제작하 였다. 관상 뇌절편은 hematoxylin 및 e o s i n으로 염색하 였다. 해마 CA1 부위의 신경세포 손상정도는 정상 신경세 포를 현미경하에서 헤아려 결정하였고 좌, 우 양측 해마의 CA1 부위 중 정중간 부위의 1 mm 폭내에 존재하는 정상 추체 신경세포를 관찰하여 평균 수치를 계산하였다. 생존세 포와 손상된 세포와의 구분은 Eke 등2 8의 기준을 따랐다.
6. 시약
PU, SD, SM 및 1 , 8 - d i a m i n o o c t a n e은 S i g m a사 (Sigma Chemical Co., St. Luise, MO, USA)의 제품 을, FNBT는 A l d r i c h사(Sigma Chemical Co., St.
Luise, MO, USA)의 제품을 사용하였으며, acetonitrile 은 J.T. Baker사(J.T.Baker Co., Phillips- burg, NJ, USA)의 제품을 사용하였다.
7. 통계분석
본 실험의 결과 분석(PA 및 조직학)은 A N O V A를 이용 하였으며 사후 검증은 Scheffe test를 이용하였다. 모든 실험 결과는 평균±표준오차로 표시하였고, p 값이 0 . 0 5 미만일 때 통계학적으로 의의가 있다고 하였다.
결 과
1. 전뇌허혈에 의한 대뇌 피질P A의 함량변동 및 i b u p r o f e n의 효과
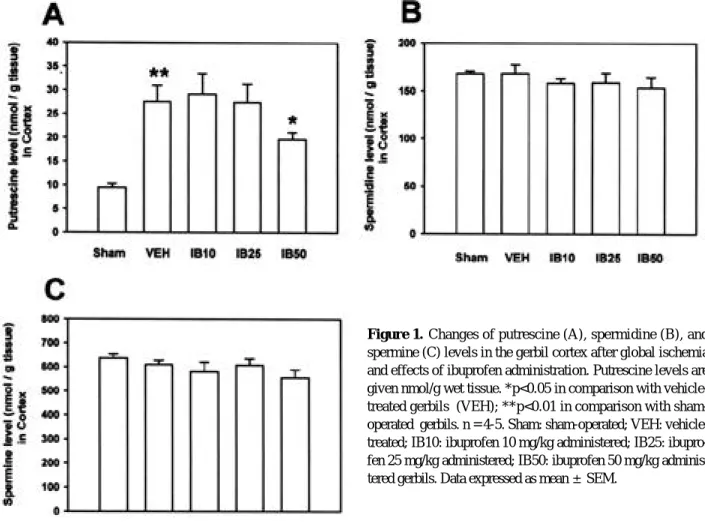
P U의 함량은 대조군인 sham 수술군( 9 . 5±0 . 8 nmol/g tissue)에 비하여 vehicle 투여군( 2 7 . 5±3 . 5 nmol/g tissue)에서 유의하게 증가하였으며( p < 0 . 0 1 , Fig. 1-A), 이러한 증가에 대해 ibuprofen 10 mg/kg 투여군( 2 9 . 2±4.3 nmol/g tissue)이나 25 mg/kg 투 여군( 2 7 . 4±3.9 nmol/g tissue)에서는 유의한 변동이 없었으나, 50 mg/kg 투여군에서는 유의하게 감소하였다 ( 1 9 . 7±1.5 nmol/g tissue, p<0.05, Fig. 1-A).
S D의 함량은 대조군인 sham 수술군( 1 6 8 . 2±2 . 5 nmol/g tissue)에 비하여 vehicle 투여군( 1 6 8 . 4±8 . 8 nmol/g tissue)에서 유의한 변동을 보이지 않았으며 (Fig. 1-B), ibuprofen 10, 25 및 50 mg/kg 투여군 (각각 1 5 8 . 2±5.2 nmol/g tissue, 158.8±5 . 2 nmol/g tissue 및 1 5 3 . 6±11.1 nmol/g tissue)에서
Figure 1. Changes of putrescine (A), spermidine (B), and spermine (C) levels in the gerbil cortex after global ischemia and effects of ibuprofen administration. Putrescine levels are given nmol/g wet tissue. *p<0.05 in comparison with vehicle- treated gerbils (VEH); **p<0.01 in comparison with sham- operated gerbils. n = 4-5. Sham: sham-operated; VEH: vehicle- treated; IB10: ibuprofen 10 mg/kg administered; IB25: ibupro- fen 25 mg/kg administered; IB50: ibuprofen 50 mg/kg adminis- tered gerbils. Data expressed as mean ±S E M .
도 vehicle 투여군에 비하여 유의한 함량변동이 없었다 (Fig. 1-B). SM의 함량은 대조군인 sham 수술군( 6 3 6
±17.2 nmol/g tissue)에 비하여 vehicle 투여군( 6 1 0
±17.2 nmol/g tissue)에서 유의한 변동을 보이지 않았 으며(Fig. 1-B), ibuprofen 10, 25 및 50 mg/kg 투 여군(각각 5 8 0±40 nmol/g tissue, 607.9±2 6 . 6 nmol/g tissue 및 5 5 5 . 4±32.8 nmol/g tissue)에서 도 vehicle 투여군에 비하여 유의한 함량 변동이 없었다 (Fig. 1-B).
2. 전뇌 허혈에 의한 해마 P A의 함량 변동 및 i b u p r o- f e n의 효과
P U의 함량은 대조군인 sham 수술군( 9 . 9±0 . 4 nmol/g tissue)에 비하여 vehicle 투여한 허혈 손상군 ( 2 2 . 1±2.1 nmol/g tissue)에서 유의하게 증가하였으며 (p<0.01, Fig. 2-A), 이러한 증가에 대해 i b u p r o f e n 10 mg/kg 투여군( 2 2 . 4±2.2 nmol/g tissue)이나 2 5 mg/kg 투여군( 2 0 . 5±3.6 nmol/g tissue)에서 유의한 변동이 없었는데 비해, 50 mg/kg 투여군에서는 유의하게 감소하였다( 1 5 . 9±1.4 nmol/g tissue, p<0.05, Fig.
2 - A ) .
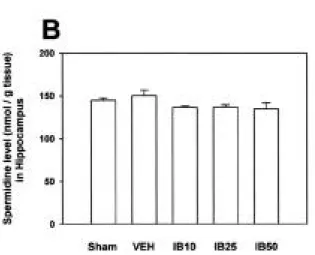
S D의 함량은 대조군인 sham 수술군( 1 4 4 . 7±2 . 7 nmol/g tissue)에 비하여 vehicle 투여군( 1 5 0 . 2±6 . 3 nmol/g tissue)에서 유의한 변동이 없었으며(Fig. 2-B),
ibuprofen 10, 25 및 50 mg/kg 투여군(각각 1 3 6 . 3±
1.8 nmol/g tissue, 136.7±2.8 nmol/g tissue 및 1 3 4 . 7±7.2 nmol/g tissue)에서도 vehicle 투여군에 비하여 유의한 함량 변동이 없었다(Fig. 2-B).
S M의 함량은 대조군인 sham 수술군( 4 5 0 . 6±1 1 . 4 nmol/g tissue)에 비하여 vehicle 투여군( 4 0 2 . 8±1 1 . 7 nmol/g tissue)에서 유의하게 감소하였으며(Fig. 2-C), ibuprofen 10, 25 및 50 mg/kg 투여군(각각 4 1 2 . 0±
39.4 nmol/g tissue, 402.2±25.9 nmol/g tissue 및 3 8 5±16.2 nmol/g tissue)에서도 vehicle 투여군에 비하여 유의한 함량 변동을 보이지 않았다(Fig. 2-C).
3. 조직학적 소견
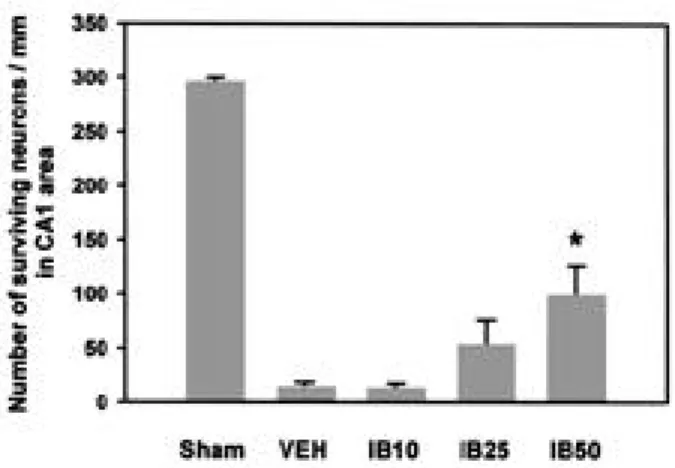
G e r b i l의 뇌조직 절편에 대한 광학현미경 소견은 해마 CA1 부위의 추체신경세포가 sham 수술군에서 2 9 4 . 5±
5.4 개에 비해 v e h i c l e을 투여한 허혈 손상군에서는 1 3 . 3
±3.7 개로 현저히 감소한 것을 볼 수 있었다( p < 0 . 0 1 , Fig. 3과 4). 이러한 신경세포손상에 대한 i b u p r o f e n의 효과를 관찰한 결과 10 mg/kg 투여군에서는 1 2 . 2±5 . 5 개로 감소하지 않았으며, 25 mg/kg 투여군에서는 5 1 . 7
±25.3 개로 신경세포 손상이 감소하였으나 유의하지 않았 다. Ibuprofen 50 mg/kg 투여군에서는 9 7 . 9±2 8 . 5 개로 신경세포손상이 유의하게 감소하였다(p<0.05, Fig.
3 , 4 ) .
Figure 2. Changes of putrescine (A), spermidine (B), and spermine (C) levels in the gerbil hippocampus after global ischemia and effects of ibuprofen administration. Putrescine levels are given nmol/g wet tissue. *p<0.05 in comparison with vehicle-treated gerbils (VEH); **p<0.01 in comparison with sham-operated gerbils. n = 4-8. Sham: sham-operated;
VEH: vehicle-treated; IB10: ibuprofen 10 mg/kg adminis- tered; IB25: ibuprofen 25 mg/kg administered; IB50: ibupro- fen 50 mg/kg administered gerbils. Data expressed as mean ± SEM.
고 찰
본 연구에서 g e r b i l에서 유발시킨 일과성 전뇌허혈 후 야 기된 PA 특히, PU의 함량 증가와 해마 신경세포 손상이 i b u p r o f e n의 투여에 의해 유의하게 억제되었다.
1. 전뇌 허혈에 의한 polyamine 함량 변동에 대한 i b u p r o f e n의 효과
본 연구는 i b u p r o f e n이 허혈에 의한 P A의 함량변동에 미치는 영향에 대해 처음 보고하는 것이다.
P A은 세포 성장, 암발생, 및 조직 손상 등에 관여한
다.1 8 , 2 5뇌내 PA 대사는 간질발작, kainate의 투여에 의
한 흥분성 뇌신경손상 및 허혈 등 다양한 자극이나 손상에 의해 영향을 받을 수 있으며, 또한 뇌조직 손상에 중요한 기전이 된다.2 9 - 3 2 P U는 PA 대사에서 속도조절 효소인 ornithine decarboxylase(ODC)의 활성증가에 의해 생 성이 증가되며, SD과 S M의 diamine 전구체로써 정상적 으로 뇌에 아주 미량으로 존재하나 스트레스성 자극 시에 효소의 활성화로 그 농도가 급격히 증가하는 물질이며 축적 시 신경조직에 유해하다. SD과 S M이 SD/SM N1- a c e t y l t r a n s f e r a s e ( S S A T )에 의해 아세틸화가 일어나고 이후 이들 물질이 PA oxidase에 의해 P U이 다시 생성되 는 반응도 P U의 함량 증가에 중요한 경로로 간주되고 있
다.3 3 SSAT 효소는 O D C와 마찬가지로 스트레스성 자극에 의해 빠른 시간 내에 활성화가 이루어지며 반감기가 짧은 효소이다.3 4 ODC 활성도의 증가 없이 P U의 함량 증가가 야기되는 경우가 있는데 이는 S S A T의 활성도가 증가하여 P U의 재합성이 이루어졌기에 가능하다.
PA 중 P U의 함량은 세포괴사 정도와 밀접한 관계가 있 으며,3 5 PU 함량의 증가정도는 스트레스 뿐만 아니라 심한 신경조직손상 정도를 잘 반영한다고 알려져 있다. 이 물질 은 뇌허혈시에 부종이나 허혈성 신경손상 등에 중요한 역할
을 하며3 5 - 3 6또한 간질에 의한 신경손상에도 관여한다.3 2 , 3 7
한편 PA 대사의 억제는 뇌손상을 줄인다는 보고들이 있 다. Paschen 등3 5은 흰쥐의 전뇌허혈 모델에서 O D C의 활 성 억제제 a l p h a - d i f l u o r o m e t h y l o r n i t h i n e이 PU 함량 을 유의하게 감소시킨 것을 관찰하였으며, Baskaya 등2 6 은 O D C를 억제시킨 후 허혈에 의한 부종형성을 억제한다 고 하였다. Dogan 등3 6은 P U의 함량 증가에 관여하는 또 다른 효소인 PA oxidase를 억제하여 허혈 시에 P U의 축 적을 막아 뇌부종 및 신경손상 등을 억제하였다고 하였다.
본 연구에서 3분간의 전뇌 허혈에 의해 대뇌피질과 해마에 서 P U의 함량이 sham 수술군에 비해 유의하게 증가하였 으며, 이 증가는 50 mg/kg의 i b u p r o f e n에 의해 유의하 게 감소하였다. 10 mg/kg와 25 mg/kg의 i b u p r o f e n에 서는 P U의 함량 증가를 유의하게 억제하지 못하는 것으로 Figure 3. Microphotographs of the hippocampal CA1 area in the gerbil 5 days after transient global ischemia (hematoxylin and eosin staining, ×200). Effect of ibuprofen administration on the number of surviving cells in the CA1 area of hippocampus five days after tran- sient global ischemia in gerbils. CA1 area in sham-operated (A), in vehicle-treated (B), in ibuprofen 10 mg/kg administered gerbils (C), in ibuprofen 50 mg/kg administered gerbils (D). Bar = 50 μm .
보아 i b u p r o f e n이 전뇌허혈에 의한 P A의 대사 변화에 영 향을 줄 수 있으려면 적어도 50 mg/kg에서 가능하다는 것을 알 수 있었다. 본 연구에서 2 4시간 재관류 후에 동물 을 희생시켜 P A을 측정한 이유는 Paschen 등2 4의 연구에 근거하였다. 그들은 전뇌 허혈 후 8시간 후부터 P U의 함량 이 유의하게 증가하기 시작하여 2 4시간 재관류 후에 가장 높게 증가하였다.
본 연구에서 전뇌허혈 후 P U의 대사산물 중 해마의 S M 은 유의하게 감소하였으나 SD 함량은 변동이 없었으며 ibuprofen 투여군들에서도 유의한 변동이 없었다. 전뇌허 혈에 기인된 O D C의 활성화에 의한 PU 함량증가는 S - adenosylmethionine decarboxylase를 활성화시켜 S D과 S M의 함량 증가를 예상할 수 있으나, 전뇌허혈에 의 해 SSAT 역시 활성화가 되어 P U의 재합성 과정이 촉진되 어 S D과 S M의 함량이 증가하지 않은 것으로 생각된다. 이 는 일반적으로 외부자극에 의해 뇌내 PA 중 S D과 S M의 함량에는 유의한 변동이 없거나 오히려 유의하게 감소한다 는 연구3 8와 일치하는 결과다.
본 연구 결과만으로는 i b u p r o f e n이 P A의 대사에 어떻 게 영향을 미치는지 증거를 제시하기는 어렵지만, ibupro- fen 투여에 의해 허혈시 관찰되는 P U의 함량증가를 유의 하게 감소시킨 결과는 COX 억제 작용 이외에 i b u p r o f e n 이 또 다른 신경보호 기전이 있음을 시사한다.
2. 전뇌허혈에 의한 해마 신경세포 손상에 대한 i b u p r o f e n의 효과
허혈시에 C O X와 관련된 염증성 신경조직손상에 대해서 는 전뇌허혈보다는 국소 허혈에 의한 뇌경색 발생에 있어
더 많은 연구가 이루어 졌다.7 , 1 4 , 4 0 , 4 1뇌허혈시 중성구의 조 기 축적이 형태학적 및 생화학적으로 밝혀졌다. 정상 뇌의 소혈관에 염증세포가 전혀 없는 것과는 달리 허혈 부위의 뇌내 소혈관은 백혈구로 차고 이들 주위에 상당부분의 부종 이 형성된다. COX의 발현은 주로 국소 허혈에 의한 뇌경 색 부위의 주변조직에서 발견된다. 한편 허혈 손상시 과다 한 g l u t a m a t e의 농도에서 비롯되는 흥분성 신경 독작용은 이미 잘 알려져 있으며 g l u t a m a t e에 의한 염증성 손상에 대한 보고들이 있다. Glutamate는 신경세포에서 C O X - 2 를 발현시키는데,4 2 C O X - 2의 활성화가 g l u t a m a t e에 의 한 독작용에 있어 한 기전을 담당하는 것으로 생각한다. 또 한 역으로 P G는 시냅스 전달체계에도 관여해 g l u t a m a t e 의 분비에 영향을 준다.4 3 또한 허혈시에 g l u t a m a t e와 산 화성 스트레스가 신경세포에서의 전사인자( t r a n s c r i p t i o n f a c t o r )인 nuclear factor kappa B(NF-κB )를 유도한
다.4 4 - 4 5NSAIDs 혹은 그 대사물들이 C O X에 대한 작용 이
외에 다른 기전의 뇌 보호작용을 나타낼 수 있는데 대한 연 구들이 진행되고 있다. NSAIDs가 g l u t a m a t e의 대사효소 에 결합하여 뇌내 g l u t a m a t e의 농도조절에 관여할 수 있 다고 보고되어 COX 억제작용 외에 뇌 보호작용을 나타낼 수 있는 또 하나의 가능성을 제시하였고,4 6 - 4 7신경세포에서 의 N F -κB의 활성을 억제하였다는 보고4 8가 있다. 본 연구 에서는 전뇌허혈에 의한 해마의 신경세포손상을 5 0 m g / k g의 ibuprofen 투여로서 유의하게 감소하였다. 이 러한 결과는 i b u p r o f e n ,9 p i r o x i c a m과 f l u r b i p r o f e n1 2 의 전뇌허혈에 의한 뇌신경세포 손상억제에 관한 연구 성적 들과 일치한다.
결론적으로 i b u p r o f e n의 투여에 의해 일과성 전뇌허혈 에 의한 PA 중 P U의 함량변동이 억제되었고, 해마의 지연 성 신경세포 손상을 억제하였음을 미루어 보아 i b u p r o f e n 도 다른 종류의 NSAIDs 처럼 허혈성 신경세포 손상에 치 료제로 이용될 수 있을 것으로 생각한다. 그러나 i b u p r o- f e n의 약리작용과 PA 대사와의 관계규명에 대해서는 향후 추가 실험이 이루어져야 한다고 생각한다.
R E F E R E N C E S
01. DeWitt DL, Smith WL. Primary structure of prostaglandin G/H synthase from sheep vesicular gland determined from the complementary DNA sequence. Proc Natl Acad Sci USA 1988;85:1412-1416.
02. Reiger MK, DeWitt DL, Shindler MS, Smith WL.
Subcellular localization of prostaglandin endoperoxide synthase-2 in murine 3T3 cells. Arch Biochem Biophys 1993;301:439-444.
03. K u j u b u D A , F l e t c h e r B S , V a r n u m B C , L i m R W , Herschman HR. TIS10, a phorbol ester tumor promotor- inducible mRNA from Swiss 3T3 cells, encodes a novel prostaglandin synthase/cyclooxygenase homologue. J Biol Chem 1991;266:12866-12872.
04. Kujubu DA, Herschman HR. Dexamethasone inhibits mitogen induction of the TIS10 prostaglandin synthase/
Figure 4. Effect of ibuprofen administration on the number of surviving cells in the CA1 area of hippocampus 5 days after transient global ischemia in gerbils. *p<0.05 in comparison with vehicle treated-gerbils. Sham, sham-operated (n = 6);
VEH, vehicle-treated (n = 6); IB10: ibuprofen 10 mg/kg administered(n = 6); IB25: ibuprofen 25 mg/kg administered (n = 6); IB50: ibuprofen 50 mg/kg administered gerbils (n = 8). Data are expressed as mean±SEM.
cyclooxygenase gene. J Biol Chem 1992;267:7991-7994.
05. O’Banion MK, Winn VD, Young DA. cDNA cloning and functional activity of a glucocorticoid-regulated inflamma- tory cyclooxygenase. Proc Natl Acd Sci USA 1992;89:4888-4892.
06. Feng L, Sun W, Xia Y, Tang WW, Chanmugam P, Soyoola E, et al. Cloning two isoforms of rat cyclooxyge- nase: differential regulation of their expression. A r c h Biochem Biophys 1993;307:361-368.
07. Nogawa S, Zhang F, Ross ME, Iadecola C. Cyclo-oxyge- nase-2 gene expression in neuronal contributes to ischemic brain damage. J Neurosci 1997;17:2746-2755.
08. Domoki F, Veltkamp R, Thrikawala N, Robins G, Bari F, Louis TM, et al. Ischemia-reperfusion rapidly increases COX-2 expression in piglet cerebral arteries. Am J Physiol 1999;277:H1207-H1214.
09. Patel PM, Drummond JC, Sano T, Cole DJ, Kalkman CJ, Yaksh TL. Effect of ibuprofen on regional eicosanoid pro- duction and neuronal injury after forebrain ischemia in rats. Brain Res 1993;614:315-324.
10. Nakayama M, Uchimura K, Zhu RL, Nagayama T, Rose ME, Stetler RA, et al. Cyclooxygenase-2 inhibition pre- vents delayed death of CA1 hippocampal neurons follow- ing global ischemia. Proc Natl Acad Sci USA 1 9 9 8 ; 9 5 : 10954-10959.
11. Nagayama M, Niwa K, Nagayama T, Ross ME, Iadecola C. The cyclooxygenase-2 inhibitor NS-398 aeliorates ischemic brain injury in wild-type mice but not in mice with deletion of the inducible nitric oxide synthase gene. J Cereb Blood Flow Metab 1999;19:1213-1219.
12. Nakagomi T, Sasaki T, Kirino T, Tamura A, Noguchi M, Saito I, et al. Effect of cyclooxygenase and lipooxygenase inhibitors on delayed neuronal death in the gerbil hip- pocampus. Stroke 1989;20:925-929.
13. Buccellati C, Folco GC, Sala A, Scelsi R, Masoero E, Poggi P, et al. Inhibition of prostanoid synthesis protects against neuronal damage induced by focal ischemia in rat brain. Neurosci Lett 1998;257:123-126.
14. Hara K, Kong DL, Sharp FR, Weinstein PR. Effect of selective inhibition of cyclooxygenase 2 on temporary focal cerebral ischemia in rats. Neurosci Lett 1998;256:53- 56.
15. Grice SC, Chappell ET, Prough DS, Whitley JM, Su M, Watkins WD. Ibuprofen improves cerebral blood flow after global cerebral ischemia in dogs. S t r o k e 1 9 8 7 ; 1 8 : 787-791.
16. Chyatte D. Prevention of chronic cerebral vasospasm in dogs with ibuprofen and high-dose methylprednisolone.
Stroke 1989;20:1021-1026.
17. Casper D, Yaparpalvi U, Rempel N, Werner P. Ibuprofen protects dopaminergic neurons against glutamate toxicity in vitro. Neurosci Lett 2000;289:201-204.
18. Tabor CW, Tabor H. Polyamines. Annu Rev Biochem 1984;53:749-790.
19. Pegg AE. Recent advances in the biochemistry of polyamines in eukaryotes. Biochem J 1986;234:249-262.
20. Williams K, Romano C, Dichter MA, Molinoff PB.
Modulation of the NMDA receptor by polyamines. L i f e Sci 1991;48:469-498.
21. Koenig H, Goldsteine AD, Lu CY. Polyamines regulate calcium fluxes in a rapid plasma membrane response.
Nature 1983;305:530-534.
22. Iqbal Z, Koenig H. Polyamines appear to be second mes- sengers in mediating Ca++ fluxes and neurotransmitter release in potassium-depolarized synaptosomes. B i o c h e m Biophys Res Commun 1985;133:563-573.
23. Bondy SC, Walker CH. Polyamines contribute to calcium- stimulated release of aspartate from brain particulate frac- tions. Brain Res 1986;371:96-100.
24. Paschen W, Schmidt-Kastner R, Hallmayer J, Djuricic B.
Polyamines in cerebral ischemia. Neurochem Pathol 1988;
9:1-20.
25. Dempsey RJ, Carney JM, Kindy MS. Modulation of ornithine decarboxylase mRNA following transient ischemia in the gerbil. J Cereb Blood Flow Metab 1991;
11:979-985.
26. Baskaya MK, Rao AM, Puckett L, Prasad MR, Dempsey RJ. Effect of difluoro- methylornithine treatment on regional ornithine decarboxylase activity and edema for- mation after experimental brain injury. J Neurotrauma 1996;13:85-92.
27. Spragg BP, Hutchings AD. High-performance liquid chro- matographic determination of putrescine, spermidine, and spermine after deprivation with 4-fluoro-3-nitrobenzotri- fluoride. J Chromatogr 1983;258:289-292.
28. Eke A, Conger KA, Anderson M. Histological assessment of neurons in rat model of cerebral ischemia. Stroke 1990;
21:299-304.
29. Pajunen AEI, Hietala OA, Virransalo E-L, Piha RS.
Ornithine decarboxylase in mouse brain effect of electrical stimulation. J Neurochem 1978;30:281-283.
30. Paschen W, Hallmayer J, Mies G. Regional profile of polyamines in reversible cerebral ischemia of Mongolian gerbils. Neurochem Pathol 1987;7:143-156.
31. Reed LJ, de Belleroche J. Induction of ornithine decar- boxylase in cerebral cortex by excitotxin lesion of nucleus basalis: association with postsynaptic responsiveness and N-methyl-D-aspartate receptor activation. J Neurochem 1990;55:780-787.
32. Martinez E, de Vera N, Artigas F. Differential response of rat brain polyamines to convulsant agents. Life Sci 1991;
48:77-84.
33. Seiler N, Bolkenius FN. Polyamine reutilization and turnover in brain. Neurochem Res 1985;10:529-544.
34. Casero RA Jr., Celano P, Ervin SJ, Applegren NB, Wiest L, Pegg AE. Isolation and characterization of a cDNA clone that codes for human spermidine/spermine N1- acetyltransferase. J Biol Chem 1991;266:810-814.
35. Paschen W, Rohn G, Meese CO, Djuricic B, Schmidt- Kastner R. Polyamine metabolism in reversible cerebral ischemia: effect of α-difluoromethylornithine. Brain Res 1988;453:9-16.
36. Dogan A, Rao AM, Hatcher J, Rao VLR, Baskaya MK, Dempsey RJ. Effects of MDL 72527, a specific inhibitor of polyamine oxidase, on brain edema, ischemic injury volume, and tissue polyamine levels in rats after tempo- rary middle cerebral artery occlusion. J Neurochem 1999;72:765-770.
37. Najm I, el-Skaf G, Massicotte G, Vanderklish P, Lynch G, Baudry M. Changes in polyamine levels and spectrin degradation following kainate-induced seizure activity:
effect of difluoromethylornithine. Exp Neurol 1 9 9 2 ; 116:345-354.
38. Paschen W, Widmann R, Weber C. Changes in regional polyamine profiles in rat brains after transient cerebral ischemia (single versus repetitive ischemia): evidence for release of polyamines from injured neurons. Neurosci Lett 1992;135:121-124.
39. Kawase M, Murakami K, Fujimura M, Morita-Fujimura Y, Gasche Y, Kondo T, et al. Exacerbation of delayed cell injury after transient global ischemia in mutant mice with CuZn superoxide dismutase deficiency. S t o k e 1 9 9 9 ; 3 0 : 1962-1968.
40. Collacomoraes Y, Aspey B, Harrison M, Debelleroche J.
Cyclo-oxygenase-2 messenger RNA induction in focal cerebral ischemia. J Cereb Blood Flow Metab 1 9 9 6 ; 1 6 : 1366-1372.
41. Sanz O, Estrada A, Ferrer I, Planas AM. Differential cellu-
ar distribution and dynamics of HSP70, cyclooxygenase-2 and c-Fos in the rat brain after transient focal ischemia or kainic acid. Neuroscience 1997;80:221-232.
42. Tocco G, Freire-Moar J, Schreiber SS, Sakhi SH, Aisen PS, Pasinetti GM. Maturational regulation and regional induction of cyclooxygenase-2 in rat brain: implications for Alzheimer’s disease. Exp Neurol 1997;144:339-349.
43. Bezzi P, Carmignoto G, Pasti L, Vesce S, Rossi D, Lodi Rizzini B, et al. Prostaglandins stimulate calcium-depen- dent glutamate release in astrocytes. N a t u r e 1 9 9 8 ; 3 9 1 : 281-285.
44. Kaltschmidt C, Kaltschmidt B, Baeuerle PA. Brain synapses contain inducible forms of the transcription fac- tor NF-κB. Mech Dev 1993;43:135-147.
45. Hunot S, Brugg B, Richard D, Michel PP, Muriel MP, Ruberg M, et al. Nuclear translocation of NF- κB is increased in dopaminergic neurons of patients with Parkinson’s disease. Proc Natl Acad Sci USA 1 9 9 7 ; 9 4 : 7531-7536.
46. B u l e r a S J , B i r g e R B , C o h e n S D , K h a i r a l l a h E A . Identification of the mouse liver 44-kDa acetaminophen- binding protein as a subunit of glutamine synthase.
Toxicol Appl Pharmacol 1995;134:313-320.
47. Halmes NC, Hinson JA, Martin BM, Pumford NR.
Glutamate dehydrogenase covalently binds to a reactive metabolite of acetaminophen. Chem Res Toxicol 1 9 9 6 ; 9 : 541-546.
48. Grilli M, Pizzi M, Memo M, Spano P. Neuroprotection by aspirin and sodium salicylate through blockade of NF- kappa B activation. Science 1996;274:1383-1385.