Vol. 43, No. 2, June 2017, 103-114 http://dx.doi.org/10.15230/SCSK.2017.43.2.103
1)
† 주 저자 (e-mail: [email protected]) call: 070)5117-0034
떡쑥 추출물의 항염증 및 항알러지 효과
노 경 백†⋅이 정 아⋅박 준 호*⋅정 광 선*⋅정 은 선⋅박 덕 훈
바이오스펙트럼(주) 생명과학연구소, *스킨큐어(주) 생명과학연구소 (2017년 3월 22일 접수, 2017년 5월 8일 수정, 2017년 5월 17일 채택)
Anti-Inflammatory and Anti-allergic Effects of Gnaphalium affine Extract
Kyung-Baeg Roh†, Jung-A Lee, Junho Park*, Kwangseon Jung*, Eunsun Jung, and Deokhoon Park
Biospectrum Life Science Institute, 767 Sinsu-ro, Yongin-si, Gyeonggi-do 16827, Korea
*Skincure Life Science Institute, 767 Sinsu-ro, Yongin-si, Gyeonggi-do 16827, Korea (Received March 22, 2017; Revised May 8, 2017; Accepted May 17, 2017)
요 약: 떡쑥(Gnaphalium affine D. Don, GA)은 동아시아 지역에서 식용으로 사용되고 있으며, 예로부터 전통 적인 민간요법 약재로 사용되어 왔다. 현재 떡쑥 추출물(GA extract, GAE)의 항산화 활성과 항보체 활성 등은 알려져 있으나, 항염과 항알러지 효능 및 그 작용 기작은 자세히 알려져 있지 않다. 본 연구에서는 염증 매개인자 인 산화질소, 프로스타글란딘 E2, Toll-유사수용체 4, 에오탁신-1, 히스타민의 활성화에 대한 GAE의 저해효과 를 평가하였다. 본 연구를 통해, GAE는 유도성 산화질소 합성효소와 COX-2의 발현을 저해함을 확인하였으며,
이를 통해 산화질소와 프로스타글란딘 E2의 생성을 저해함을 확인하였다. GAE는 LPS로부터 유도된 Toll-유
사수용체 4의 발현에도 영향을 미치는 것을 확인하였으며, A23187로부터 유도되는 비만세포의 히스타민 방출 의 억제에도 효과적으로 작용하는 것을 확인하였다. 또한 IL-4로부터 유도된 에오탁신-1의 생성도 효과적으로 억제하는 결과를 확인하였다. 이상의 결과로부터 GAE는 항염증과 항알러지 효능을 가진다고 사료되며, 향후 항염증 및 항알러지 화장품 원료로서의 이용가능성을 보였다.
Abstract: Gnaphalium affine D. DON (GA) has been used as a vegetable as well as a folk medicine in East Asia. The antioxidant and anti-complementary activity of GA extract (GAE) has also been reported. However, little is known about its anti-inflammatory and anti-allergic effect and mechanism of action. In this study, we evaluated the inhibitory effects of GAE on the production of inflammatory mediators such as NO, PGE2, TLR4, eotaxin-1 and histamine. Our results suggest that GAE inhibits the production of NO and PGE2 by inhibiting transcriptional activation via the involvement of iNOS and COX-2. The LPS-induced expression of Toll-like receptor 4 (TLR4) was also attenuated. In addition, GAE inhibited A23187-induced histamine release from MC/9 mast cells. It also inhibited the production of eotaxin-1 induced by IL-4. Collectively, these results suggest that GAE may have considerable potential as a cosmetic ingredient with anti-inflammatory and anti-allergic properties.
Keywords: Gnaphalium affine D. DON, nitric oxide, prostaglandin E2, eotaxin-1, histamine
1. Introduction
Cosmetics are commercially available products that are used to improve the appearance of the skin. Nowadays, natural ingredients are becoming more prevalent in cos- metic industry, due to concerns about synthetic in- gredients and chemical substances. Natural ingredients have been used for skin care purposes for centuries, and recent research has proven their scientific efficacy.
Therefore, consumers are looking for natural cosmetic in- gredients that actually do something beneficial to their skin, because these natural products are safe to use and do not have any side effects.
Gnaphalium affine D. Don (GA) has been used as a food and folk medicine for its antitussive and expectorant activities[1]. A recent study showed that GA extract (GAE) exhibits anti-complementary, antifeedant, anti- microbial and antioxidant activities[2-4]. Moreover, it had been reported to be useful for the treatment of respiratory diseases[5]. However, the precise biochemical mecha- nisms underlying its anti-inflammatory and anti-allergic effects have not yet been clarified.
Inflammation is characterized by the coordinate activa- tion of various signaling pathways that regulate the ex- pression of inflammatory mediators in resident tissue cells and leukocytes recruited from the blood. It is a complex biological process involving the response of vascular tis- sues to stimuli, such as bacteria, damaged cells, or irritants. Lipopolysaccharide (LPS), a component of the outer membrane of gram negative bacteria, activates pro-inflammatory mediators which include tumor necrosis actor alpha (TNF-a), interleukins (ILs), matrix metal- loproteinases (MMPs), and related pro-inflammatory me- diators such as nitric oxide (NO) and prostaglandin E
2(PGE
2)[6-9]. Cyclooxygenase-2 (COX-2) and inducible nitricoxide synthase (iNOS) are related to the production of two major inflammatory mediators, viz. PGE
2and NO, respectively[10].
The expression of NO and PGE
2is first regulated and modulated by cellular receptor molecules such as Toll-lik- ereceptors (TLRs) and cluster of differentiation 14
(CD14). TLR4, located on the cel plasma membrane or within the endosome, associates with CD14 and myeloid differentiation factor-2 (MD-2) to recognize LPS[11].
LPS/lipopolysaccharide binding protein (LBP) complex is subsequently recognized by CD14 and then presented to the TLR4/MD-2 complex[12]. TLR4 activation induces a pro-inflammatory environment through the mitogen-activated protein kinases (MAPKs), NF-κB and AP-1 path- ways[13]. The expression of iNOS and COX-2 is induced by mitogenic and proinflammatory stimuli and is regu- lated by p38 MAPK and/or NF-κB[14,15]. These intra- cellular signaling pathways are involved in inflammatory responses.
Mast cells and eosinophilic granulocytes are crucial effector cells, in allergic disorders and auto-immune disorders. Their products, such as cytotoxic granule proteins, histamine, leukotrienes, and cytokines are in- volved in the pathological changes seen at the sites of inflammation. Mast cells play a major role in inflammatory and allergic reactions. Mast cells are localized in the con- nective tissue and do not usually circulate in the blood stream[16]. Eosinophils are recruited to the sites of in- flammation by locally released chemotactic factors. The CC chemokine, eotaxin-1, a potent chemoattractant for eosino- phils, is a critical mediator for the development and perpetu- ation of allergen-induced eosinophilic inflammation[17].
Eotaxin-1 is produced at high levels in patients with atopic dermatitis, and is localized to the inflammation site. In the present study, we investigated the inhibitory effect of GAE and the mechanism of its anti-inflammatory and anti-aller- gic effect.
2. Materials and Methods
2.1. Preparation of Gnaphalium affine D. Don Extract
GA was harvested in Jeju Island, Korea, from June
2013. The whole plant of GA was thoroughly washed
with water, shade dried and grind into powder. The dried
powder (90 g) was extracted with 70% ethanol (2 L) for
24 h, and the extract was incrassated by a rotary evapo-
rator for 3 h. To remove the ethanol from the extract,
it was mixed with water and incrassated again.
Subsequently, the extracted liquid was filtered through fil- ter paper and frozen on a freezing tray for 48 h. Freeze drying for 60 h afforded a perfectly dried extract of GA (20 g), which was dissolved in dimethyl sulfoxide (DMSO, Sigma, USA) for the experiments.
2.2. Chemical Analysis
The 70% ethanol extracts of GA were quantitative ana- lyzed by HPLC. A Shimadzu (Japan) HPLC system with a CBM-20A controller, SPD-M20A PDA detector, LC-20AD pump, and a SIL-20A auto sampler was used for analysis of GA. Data acquisition was achieved using the Shimadzu Lab Solution. All chromatographic separa- tions were conducted on a CAPCELL PAK C18 UG120 column (250 mm × 4.6 mm, 5 mm, Shiseido, Japan) at ambient temperature with detection at 273 nm. The mo- bile phase consisted of 0.1% trifluoroacetic acid contain- ing water (A) and acetonitrile (B) with gradient elution as follows: 0-15 min, 5-10% B; 15-30 min, 10-30% B;
30-50 min, 30-50% B; 50-70 min, 50-80% B; 70-75 min, 80-100% B; 75-85 min, 100% B; 85-95 min, 100-5% B;
95-100 min, 5% B. The amount of chlorogenic acid, caf- feic acid, scopoletin, luteolin, and apigenin in the 70%
ethanol extract of GA were determined.
2.3. Cell Culture and Reagents
The RAW264.7, mouse macrophage cell line was main- tained in Dulbeccoʼs Modified Eagleʼs Medium (DMEM, Gibco, USA), containing 10% fetal bovine serum (FBS, Gibco) and 1% penicillin/streptomycin (Invitrogen, USA), at 37 ℃, under 5% CO
2. NIH/3T3 mouse fibroblast cell line was maintained in DMEM, containing 10% FBS and 1% penicillin/streptomycin, at 37℃, under 5% CO
2. MC/9 mouse mast cell line was maintained in DMEM, containing 10% T-cell culture supplement with ConA (T-STIM, USA), 10% FBS, 0.05 mM 2-mercaptoethanol (Sigma, USA), and 1% penicillin/streptomycin, at 37 ℃, under 5%
CO
2. Lipopolysaccharides (LPS), A23187, and Griess reagent were purchased from Sigma. Mitogen-activated kinases (MAPKs) antibodies (p44/42 MAPK, phos-
pho-p44/42 MAPK, p38 MAPK, phospho-p38 MAPK, SAPK/JNK, phospho-SAPK/JNK) were purchased from Cell Signaling Technology, Inc. (USA). Mouse IL-4, mouse CD14-FITC, mouse TLR4-PE, rat IgG2a kap- pa-FITC, and mouse IgG1-PE were all purchased from eBioscience (USA).
2.4. Nitric Oxide Determination
The concentration of NO in the culture supernatant was determined as nitrite, a major stable product of NO. The cells were incubated in the presence or absence of 10, 100, and 200 µg/mL GAE for 1 h and induced by LPS (200 ng/mL) for 24 h. 50 mL of cell culture supernatant was incubated with equal volume of Griess reagent for 30 min at room temperature. The absorbance was measured by a spectrometer at a wavelength of 540 nm[18].
2.5. Cell Viability Assay
Cell viability was measured using the MTT (3-[4,5- dimethylthiazol-2-yl]-2,5-diphenyltetrazolium bromide;
USB Corp., USA) assay. The cells were treated with 10, 100, 200, and 400 µg/mL GAE for 24 h. Then, MTT re- agent (1 mg/mL) was added to each well, and the cells were incubated for 3 h. The medium was removed, and the cells were solubilized with DMSO. The absorbance was measured by spectrophotometer at a wavelength of 570 nm.
2.6. Enzyme-linked Immunosorbent Assay (ELISA)
The PGE
2and histamine concentrations were quanti-
fied by using a commercially available ELISA kit (Enzo
Life Sciences Inc., USA). The eotaxin-1 concentrations
were quantified by using a commercially available
ELISA kit eBioscience (USA). RAW264.7 cells were
treated with 10, 100, and 200 µg/mL GAE and induced
by LPS (200 ng/mL) for 24 h. The cell culture super-
natant was assayed for PGE
2. NIH/3T3 cells were treated
with 10, 100, and 200 µg/mL GAE and induced by IL-4
(50 ng/mL) for 24h. The cell culture supernatant was as-
sayed for eotaxin-1. MC/9 cells were treated with 10,
100, and 200 µg/mL GAE and induced by A23187 (500
nM) for 12 h. The cell culture supernatant was assayed for histamine. The standard curve was linearized and subjected to regression analysis. The PGE
2, eotaxin-1, and histamine concentrations were determined using a standard curve[19-21]. All samples and standards were measured in duplicate.
2.7. Transient Transfection and Luciferase Assay RAW264.7 cells were transfected with the iNOS, COX-2, AP-1, and NF-κB luciferase reporters using SuperFect transfection reagent (Qiagen, Germany) ac- cording to the manufacturer’s instructions. After 24 h of incubation, the cells were incubated in the presence or absence of GAE (10, 100, and 200 µg/mL) induced by LPS (200 ng/mL) for 24 h. The cells were then harvested and lysed by luciferase cell culture lysis reagent (Promega, USA), and the 100 mL of supernatant was as- sayed for their luciferase activity using a dual luciferase assay system (Promega, USA), and an Infinite 200 PRO luminometer (Tecan, Switzerland).
2.8. Western Blotting
Western blotting was performed to measure the levels of proteins associated with the MAPKs pathways.
RAW264.7 cells treated with 10, 100, and 200 µg/mL GAE and induced by LPS (200 ng/mL) for 24 h. The cells were then harvested and lysed by protein extraction solution, PRO-PREP
TM(iNtRON Biotechnology, Korea).
The protein extracts (30 µg) from lysed cells were loaded on a NuPAGE Novex 10% Bis-Tris Gel 1.0 mm, 10 well (Invitrogen, USA) in each lane and transferred to a nitrocellulose membrane by using iBlot
TMGel Transfer Device (Invitrogen, USA). The membranes were blocked with 5% bovine serum albumin (BSA) for 1 h and then incubated with primary antibodies (1 : 1000 dilution of stock) in 10 mL of blocking buffer, followed by incubation with horse radish peroxidase-conjugated anti-mouse or rabbit IgG secondary antibody (1 : 1000 dilution of stock) and detected using chemiluminescent HRP substrate (SurModics, USA) on auto radiographic film.
2.9. Flow Cytometry
To assess the expression of TLR4 and CD14 on the surface of the RAW264.7 cells by flow cytometry, RAW264.7 cells were treated with 10, and 100 µg/mL GAE and induced by LPS (200 ng/mL) for 24 h.
RAW264.7 cells were suspended in PBS containing 1%
FBS and then, the cells were stained with fluorescent antibodies, 0.25 µg of anti-mouse TLR4-PE, 0.5 µg of anti-mouse CD14-FITC, 0.5 µg of mouse IgG1 kappa isotype control PE, and 0.5 µg of rat IgG2a kappa isotype control FITC for 40 min on ice and then analyzed using flow cytometry (Accuri
TMC6 Flow Cytometer, BD biosciences, USA).
2.10. Statistical Analysis
All data are expressed as means ± standard deviations.
Differences between the control and the treatment group were evaluated by one way ANOVA. P < 0.05 or 0.01 was considered statistically significant.
3. Results
3.1. Chemical Composition Analysis of GAE
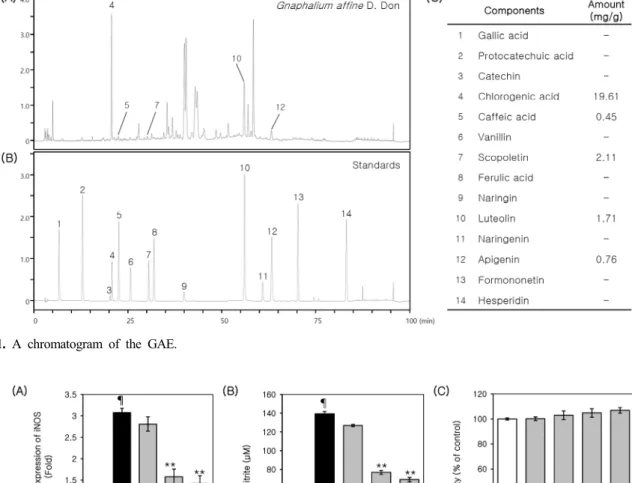
GAE used in this study contained 1.96% of chlorogenic acid, 0.045% of caffeic acid, 0.211% of scopoletin, 0.17%
of luteolin, and 0.076% of apigenin. Chromatograms of typical extracts and the mixed standards were shown in Figure 1A and Figure 1B. The results of quantitative anal- ysis were summarized in Figure 1C.
3.2. GAE Inhibits iNOS Expression and NO Production in LPS-induced RAW264.7 Cells
NO is synthesized from L-arginine in a reaction cata-
lyzed by the nitric oxide synthase (NOS) family of
proteins. iNOS is primarily responsible for the production
of NO in inflammatory processes, is not typically ex-
pressed in resting cells, but is induced by certain cyto-
kines or microbial products. The aberrant release of NO
can lead to the amplification of inflammation, as well as
tissue injury[22]. In this study, we investigated the effect
of GAE on LPS-induced iNOS expression and NO pro-
duction in RAW264.7 cells. As a first step, a luciferase reporter assay was performed to measure iNOS expression. As shown in Figure 2A, the LPS-induced acti- vation of iNOS promoter was markedly attenuated by GAE (10, 100 and 200 µg/mL). To examine the effect of GAE on NO production, we measured the level of nitrite using the Griess reaction. As shown in Figure 2B, GAE (10, 100, and 200 µg/mL) significantly inhibited the
LPS-induced nitrite production in a concentration depend- ent manner. However, cytotoxicity was not observed in GAE treated RAW264.7 cells at the concentration range of 10-400 µg/mL (Figure 2C). These results indicate that the GAE-mediated inhibition of NO production is asso- ciated with the inhibition of iNOS expression at the tran- scription level.
Figure 1. A chromatogram of the GAE.
Figure 2. Effects of GAE on iNOS expression and NO production in LPS-induced RAW264.7 cells. (A) The cells were pretreated with GAE for 1 h and then stimulated with LPS. Luciferase activity was calculated against an LPS-unstimulated control. (B) The cells were pretreated with the indicated concentrations of GAE for 1 h and then further incubated with LPS (200 ng/mL) for 24 h.
The amount of NO produced was then determined using a Griess assay. (C) Cell viability was measured by MTT assay. The results are mean ± standard deviation (S.D.) (n = 3). ¶P < 0.01 vs. LPS-untreated control. **P < 0.01 vs. LPS-treated control.
3.3. GAE Inhibits COX-2 Expression and PGE
2Production in LPS-induced RAW264.7 Cells
Prostaglandins (PGs) play a key role in the production of the inflammatory response. Their gene expression is significantly upregulated in inflamed tissue and they con- tribute to the development of acute inflammation. PGE
2is one of the most abundant PGs produced in the body and is an important mediator of the immune system. PGE
2is also involved in the process leading to the signs of in- flammation, such as redness, swelling, and pain[23]. In the inflammatory process, the synthesis of PGE
2is mainly controlled by the local expression and activity of COX-2[23]. Thus, we examined the effect of GAE on the LPS-induced expression of COX-2 using the luciferase reporter assay. As shown in Figure 3A, the LPS-induced activation of COX-2 promoter was markedly attenuated by GAE (10, 100 and 200 µg/mL). Consistent with these results, PGE
2production was also reduced by GAE (10, 100 and 200 µg/mL) (Figure 3B). These results indicate that the GAE-mediated inhibition of PGE
2production is associated with the inhibition of COX-2 expression.
3.4. GAE Inhibits LPS-induced Phosphorylation of p38 MAPK and JNK
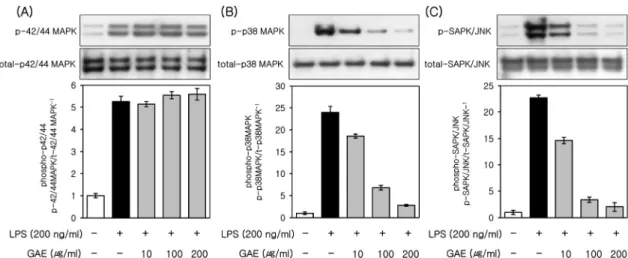
MAPKs are a family of serine/threonine protein kinases
that mediate biological processes and cellular responses to external stress signals. The high activity of MAPKs, and their regulatory role in the synthesis of inflammation me- diators, makes them potential targets for anti-in- flammatory therapeutics[24]. To investigate whether these signaling pathways are involved in the GAE effects, we performed western blot analysis for MAPKs (p42/44 MAPK, p38 MAPK, and SAPK/JNK). As shown in Figure 4, the phosphorylation of MAPKs was induced by LPS (200 ng/mL). In addition, the phosphorylation of p38 MAPK and SAPK/JNK was inhibited by GAE (10, 100 and 200 µg/mL) (Figure 4B and 4C), whereas GAE (10, 100 and 200 µg/mL) had no effect on that of p 42/44 MAPK (Figure 4A). These results suggest that GAE par- ticularly attenuates LPS-induced phosphorylation of p38 MAPK and JNK in the LPS-induced activation of MAPKs.
3.5. Effects of GAE on LPS-induced AP-1 and NF-κB Activation
The activation of the transcription factors, AP-1 or NF-
κB, induces the expression of the target genes that are
involved in inflammation and these two transcription fac-
tors play an important role in the gene expression of
iNOS and COX-2. We thus examined the effects of GAE
Figure 3. Effects of GAE on COX-2 and PGE2 expression in LPS-induced RAW264.7 cells. (A) The cells were pretreated with GAE for 1 h and then stimulated with LPS. The luciferase activity was calculated against an LPS-unstimulated control. (B) The cells were pretreated with GAE for 1 h and then further incubated with LPS (200 ng/mL) for 24 h. PGE2 release was then determined using ELISA. The results are mean ± standard deviation (S.D.) (n = 3). ¶P < 0.01 vs. LPS-untreated control. **P < 0.01 vs. LPS-treated control.on LPS-induced AP-1 and NF-κB activity using the luci- ferase reporter assay. The pretreatment of the cells with GAE markedly inhibited LPS-induced AP-1 and NF-κB activity over a similar concentration in the range of 10-200 µg/mL (Figure 5A and 5B). At a concentration of 200 µg/mL GAE, the LPS-induced activation of AP-1 and
NF-κB was suppressed by 40% and 47% respectively.
3.6. Effects of GAE on LPS-Induced TLR4 and CD14 Expression
LPS activates the TLR4-mediated NF-κB signaling pathway and pro-inflammatory response to regulate the
Figure 4. Effects of GAE on phosphorylation of MAPKs in LPS-induced RAW264.7 cells. The RAW264.7 cells were cultured for 24 h and then treated with LPS (200 ng/mL) in the presence or absence of GAE. Cell lysates were prepared at 20 min and then subjected to Western blot analysis. The bands for phospho-p42/44 MAPK (A), phospho-p38 MAPK (B), and phospho-JNK (C) were detected and normalized to their total forms of MAPKs. The data are representative of at least three independent experiments.¶P < 0.01 vs. LPS-untreated control. **P < 0.01 vs. LPS-treated control.
Figure 5. Effects of GAE on activation of AP-1 and NF-κB in LPS-induced RAW264.7 cells. (A) The AP-1 and (B) NF-κB gene promoter-luciferase reporter vectors were transfected into 70% confluent RAW264.7 cells and cultured for 24 h. The cells were pretreated with GAE for 1 h and then stimulated with LPS. The luciferase activity was calculated against an LPS-unstimulated vehicle control. The results are mean ± standard deviation (S.D.) (n = 3). ¶P < 0.05 vs. LPS-untreated control. **P < 0.05 vs.
LPS-treated control.
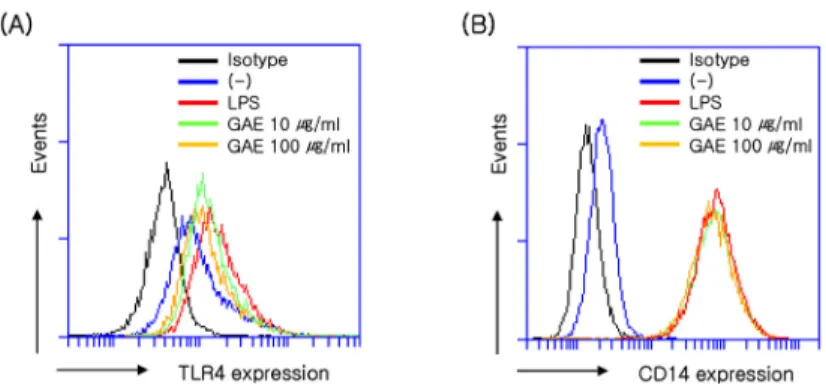
production of inflammatory mediators[25]. The LPS-in- duced signaling through TLR4 and co-receptor CD14 leads to NF-κB activation and cytokine expression[12].
To investigate whether GAE affects LPS-induced TLR4 signaling, the cell surface expression of TLR4 and CD14 on RAW264.7 cells was assessed by flow cytometry. As shown in Figure 6A and 6B, LPS induced the expression of TLR4 and CD14, as shown by the red histogram. The LPS-induced expression of TLR4 was inhibited in a con- centration dependent manner by GAE (10 and 100 µg/mL) (Figure 5A), whereas GAE (10 and 100 µg/mL) had no effect on that of CD14 (Figure 5B).
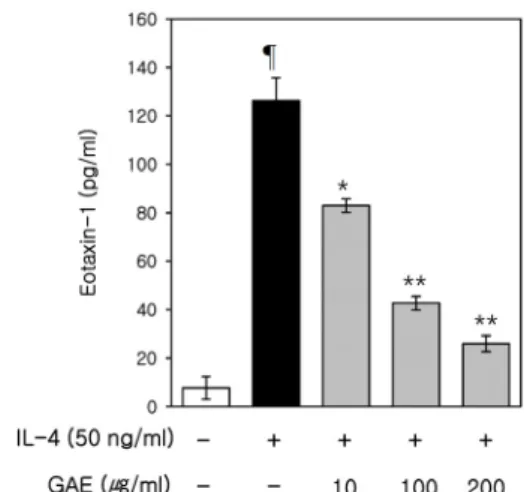
3.7. Effects of GAE on IL-4-Induced Eotaxin-1 Expression To investigate the effect of GAE on eotaxin-1 ex- pression, we investigated the effects of GAE on IL-4-in- duced expression of eotaxin-1 in NIH/3T3 cells. In this study, we used IL-4 as a stimulator which is reported to induce expression of eotaxin-1 in fibroblasts[19]. As shown in Figure 7, IL-4-induced eotaxin-1 expression was attenuated by GAE. These results indicate that GAE ex- erts an inhibitory effect on IL-4-induced eotaxin-1 expression.
3.8. Effects of GAE on A23187-Induced Histamine Release
Among diverse inflammatory mediators, histamine is a representative mediator that plays a various biological
roles in mast cell mediated allergic inflammation[16]. In order to determine whether GAE can modulate histamine degranulation of mast cells, we measured the histamine using a commercially available ELISA kit at 12 h after stimulation. A concentration of 500 nM of A23187 was selected for optimal stimulation in subsequent experiments. As shown in Figure 8, A23187-induced his- tamine release was significantly inhibited by GAE in a
Figure 6. Effects of GAE on TLR4 and CD14 expression in LPS-induced RAW264.7 cells. RAW264.7 cells were cultured for 24 h in DMEM containing GAE. The cell surface expressions of TLR4 (A) and CD14 (B) were determined by flow cytometry. (-) control cells were treated with 0.1% DMSO as a control. (-): LPS-untreated control. The data are representative of at least three independent experiments.Figure 7. Effects of GAE on eotaxin-1 expression in IL-4-induced NIH/3T3 cells. Cells were pretreated with GAE for 1 h, and then further incubated with IL-4 (50 ng/mL), for 24 h.
Eotaxin-1 release was then determined, using an ELISA. (-):
IL-4-untreated control. The data are representative of at least three independent experiments. ¶P < 0.01 vs. IL-4-untreated control. *P < 0.05 vs. IL-4-treated control, **P < 0.01 vs.
A23187-treated control.
dose-dependent manner. These results suggest that GAE attenuates release of histamine in the A23187-induced de- granulation of mast cells.
4. Discussion and Conclusion
Inflammation and allergies are a pervasive phenomenon that occurs during severe perturbations of homeostasis, such as infection, injury, or exposure to allergen. It is triggered by immune receptors that recognize pathogens and damaged cells. The immune response is governed by strict effector and regulatory processes that normally result in protection from infection and tolerance of in- nocuous environmental antigens. Recently, there has been increasing interest in phyto-constituents such as phenolics, alkaloids, and terpenoids, as more reports are published demonstrating their role in the modulation of in- flammatory responses[26].
Naturally occurring bioactive compounds, such as plant extracts or pure compounds, have great potential for drug development because of the unmatched availability of chemical diversity[27]. It is widely known that phenolic
compounds, derivatives of plant origin, possess a broad spectrum of biological activity. They exert antimicrobial, antiviral, anti-athrosclerosis, cardioprotective, antidiabetic, antioxidant, anti-ageing, antihepatotoxic, antihypertensive, hypolipideamic, antiplatelet, and anti-inflammatory activ- ities[28].
In this study, 70% ethanol extracts of GA were found to possess anti-inflammatory and anti-allergic properties and these are comprised of five major constituents, chlorogenic acid, caffeic acid, luteolin, apigenin, and scopoletin.
Chlorogenic acid, which is observed the most abundant polyphenol compounds from GAE in this study, is a group of phenol secondary metabolites with biological ef- fects related to their antioxidant and anti-inflammatory ac- tivities[29,30]. Qin et al.[31] investigated that natural products which contain chlorogenic acid inhibit histamine release form mast cells. Caffeic acid exhibits a wide range of biological roles including anti-thrombosis, anti-hyper- tension, anti-fibrosis, antivirus and antitumor proper- ties[32,33]. Hossen et al.[34] reported that caffeic acid showed inhibitory effect of histamine release, ad- ministered orally to mice. Luteolin is a widely distributed flavonoid found in many herbal extracts and medicinal plants. Preclinical studies have shown that luteolin pos- sesses a variety of pharmacological activities, including antioxidant, anticancer, anti-allergic, antimicrobial, anti-in- flammatory and anti-allergic activities[35,36]. Apigenin, a common dietary flavonoid abundant in fruits and vegeta- bles which shows promise for its anti-cancer, anti- microbial, anti-inflammatory, and anti-allergic proper- ties[37,38]. Scopoletin is a coumarin compound with an- ti-fungal, anti-oxidant, anticholinesterase, anti-angio- genesis, anti-allergy, and anti-inflammatory properties[39].
The anti-inflammatory and anti-allergic effect observed in this study might be due to the presence of these com- pounds in GAE.
Collectively, these suggest that GAE has potential as a cosmetic ingredient and might represent an alternative to standard treatments for inflammatory and allergic diseases.
Figure 8. Effects of GAE on histamine release in A23187-induced MC/9 cells. Cells were pretreated with GAE for 1 h, and then further incubated with A23187 (500 nM), for 12 h. Eotaxin-1 release was then determined, using an ELISA. (-): A23187-untreated control. The data are representative of at least three independent experiments.
¶P < 0.01 vs. A23187-untreated control. **P < 0.05 vs.
A23187-treated control.