Obesity Regulation through Gut Microbiota Modulation and Adipose Tissue Browning
Yejin Cho1, Rahman Md. Shamim2,3 and Yong-Sik Kim1,2,3*
1College of Medicine, Soonchunhyang University, Cheonan, Chung-nam 31151, Korea
2Department of Microbiology, College of Medicine, Soonchunhyang University, Cheonan, Chung-nam 31151, Korea
3Institute of Tissue Regeneration, College of Medicine, Soonchunhyang University, Cheonan, Chung-nam 31151, Korea Received July 11, 2019 /Revised July 29, 2019 /Accepted August 5, 2019
Obesity, represented by abnormal fat accumulation due to an imbalance between energy intake and expenditure, is a major public health issue worldwide, leading to multiple noncommunicable diseases, including atherosclerosis, hypertension, type 2 diabetes, and cancer. Diverse solutions have been pro- posed to combat obesity. Attention has focused on two types of adipose tissues as a promising ther- apeutic target in obesity: traditional brown and beige or brite. Unlike energy-storing white adipose (endocrine) tissue, traditional brown adipose tissue and beige adipose tissue have energy-dissipating thermogenic properties. Both types of tissue are present in adult humans and inducible through ex- ternal stimuli, such as cold exposure, β3-adrenergic receptor agonists, and phytochemicals. Among these stimuli, microbiota present in the human intestinal tract participate in multiple metabolic activities. Modulation of gut microbiota may offer a potent and possibly curative strategy against vari- ous metabolic diseases. Numerous studies have focused on the effects of established antiobesity treat- ments on the gut microenvironment or brown-adipose-tissue activation. In this review, we focus main- ly on stimuli known to alleviate obesity, weight gain, and metabolic diseases, in addition to known and possible inter-relations between gut microbiota modulation and similar interventions and adipose tissue browning. The findings may pave the way toward new strategies against obesity.
Key words : Brown adipose tissue, gut microbiota, obesity, thermogenesis, Ucp1
*Corresponding author
*Tel : +82-41-570-2413, Fax : +82-41-575-2412
*E-mail : [email protected]
This is an Open-Access article distributed under the terms of the Creative Commons Attribution Non-Commercial License (http://creativecommons.org/licenses/by-nc/3.0) which permits unrestricted non-commercial use, distribution, and reproduction in any medium, provided the original work is properly cited.
ISSN (Print) 1225-9918 ISSN (Online) 2287-3406 Journal of Life Science 2019 Vol. 29. No. 8. 922~940 DOI : https://doi.org/10.5352/JLS.2019.29.8.922
Introduction
Obesity is usually represented by anomalous or inordi- nate fat accumulation that can harm one’s health, and is thought to result from an upset energy balance owing to either overabundant energy intake or deficient energy ex- penditure [49]. It has been receiving increasing attention as a major global issue of public health. If obesity is roughly defined as a body–mass index (BMI) of 30 or over and over- weight as a BMI of 25 or higher, then more than 1.9 billion adults and 124 million children and adolescents were obese or overweight in 2016, which means that the obese pop- ulation has almost tripled since 1975[90]. Given that obesity is tightly linked with multiple chronic noncommunicable diseases including diabetes, cancers, and cardiovascular and
musculoskeletal disorders [89], innumerable societal and bi- omedical efforts have been applied to the fight against obesity.
Although the primary intervention for patients with obe- sity is aimed at lifestyle changes including diet, behavioral patterns, and physical activity, those who find this approach ineffective have relied upon medication, medical devices, and surgical interventions [56]. Pharmacotherapy with a life- style intervention contributes to weight loss, but the efficacy often ceases after discontinuation of the medication [9].
Bariatric surgery impressively reduces body weight and alle- viates the comorbidities of obesity (especially of extreme obesity), yet the financial burden and serious risks to health hinder its widespread clinical application [92]. Medical de- vices with minimal invasiveness have been approved for short-time use, and the weight loss persists after their re- moval; however, the device use is limited to patients without active gastric diseases, nonsteroidal anti-inflammatory drug use, or previous gastric surgical procedures [56]. Thus, sus- tainable noninvasive remedies with fewer adverse effects have been pursued.
- Review -
Journal of Life Science 2019, Vol. 29. No. 8 923
White, Brown, and Beige Adipose Tissues
Three distinct types of mammalian adipose tissue have been found so far: white, brown, and beige (or brite, brown- in-white). White adipose tissue (WAT), which has tradition- ally been the best-known type of adipose tissue, is currently thought to be an active endocrine organ (secreting proteins and steroid hormones) rather than a mere energy repository [58]. On the other hand, two types of major thermogenic adipose tissues—brown and beige—dissipate energy, usu- ally in the form of heat, as uncoupling protein 1 (UCP1) uncouples anion flow or the oxidative phosphorylation path- way from ATP synthesis [112]. “Classical” brown-adipocyte development starts prenatally in dermomyotomes, and transdifferentiation into brown adipocytes is known to be irreversible in vivo after the commitment phase; by contrast, beige adipocytes have inducible and reversible traits: tempo- rary expression of thermogenic attributes depending on ex- ternal stimuli such as cold acclimation, tissue injury, and prolonged treatment with PPARγ (peroxisome proliferator- activated receptor γ) or β3-adrenergic receptor agonists [51, 130]. This formation of UCP1-rich thermogenic adipocytes in white adipose depots is referred to as browning [19].
Previously, brown adipose tissues (BATs) have been thought to be present and practically useful in small mam- mals and infants, whereas adult human brown adipocytes have been thought to merely exist or manifest hardly any activity [47]. Nevertheless, significant brown adipose depots have been revealed to be present in adult humans [123], es- pecially more frequently among lean people [120]. Further- more, human thermogenic adipose activity has been found to be inducible by such stimuli as cold exposure, dietary polyphenol consumption, stimulation of the sympathetic nervous system, β-adrenergic receptor agonists, and combi- nations of the above-mentioned factors [13, 26, 87, 110, 120].
Such findings have led to a paradigm shift in the remedies for obesity and for associated metabolic disorders: from tra- ditional quantitative changes in adipose tissue to the in- duction of a thermogenic brown or beige adipose tissue.
Numerous measures were explored recently to promote brown adipose thermogenesis, including irisin and thyroid hormone induction, β3-adrenergic receptor agonists, thiazo- lidinediones, and phytochemicals [11, 12, 117, 126]. Among such browning stimuli, in this review, we will focus on the effects of the gut microbiota.
The Gut Microbiota as a Modulator of Human Metab- olism
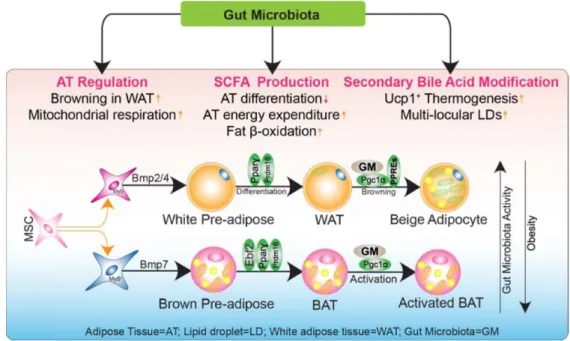
Trillions of microbial cells coexist with human host cells, especially within intestines, and engage in numerous pro- tective, structural, and metabolic activities. The gut micro- biome interacts with the host similarly to the way a complex endocrine organ does, thereby 1) affecting distant organs and systems by the release of metabolic products into the circulation and 2) being affected by hormonal signals se- creted by the host. Hence, it is suggested that the gut micro- biota—at least in part—is causally linked to major metabolic disorders including obesity [24]. When the microbiotas of subjects that have received browning-inducing stimuli are transplanted to germ-free recipients, the browning of WAT and activation of BAT strengthen in the recipients [115].
Such findings suggest that the gut microbiota is responsible for the transdifferentiation of adipocytes and controls ther- mogenic functions (Fig. 1). Thus, modulation of the gut mi- crobiota through dietary manipulation, exposure to a lower temperature, or probiotic or nutraceutical supplements may be promising antiobesity treatments.
Probiotics and Prebiotics Are Expected to Promote Adipocyte Browning
Firmicutes and Bacteroidetes are the most dominant taxa of the human gut microbiota, although some elusive but sig- nificant variations are present across different human pop- ulations [16]. Bifidobacterium and Lactobacillus are the most extensively utilized bacterial genera because of various pro- biotic properties [104]. Thus, they are currently promising candidates for a browning inducer. Accumulated data on probiotic and synbiotic interventions into obesity and meta- bolic diseases back up the potential therapeutic effects of the intestinal microbiota (Table 1). Lactobacillus reuteri 263, a probiotic strain known to alleviate renal fibrosis and in- sulin resistance, has been found to upregulate thermogenesis in WAT; to induce the expression of browning-related genes, including Ucp1, Pparγ, Prdm16, Pgc1α, Bmp7, and Fgf21; and to upregulate mitochondrial respiration in WAT [21].
There are documented cases where supplements reversed the changes in the gut microbiota composition caused by genetically or nutritionally induced obesity (Table 2). Li et al. have demonstrated that fucosylated chondroitin sulfate, a sulfated polysaccharide in the sea cucumber Isostichopus badionotus, normalizes multiple metabolic-syndrome–re- lated indices [65]. An et al. have reported that cordycepin,
924 생명과학회지 2019, Vol. 29. No. 8
Journal of Life Science 2019, Vol. 29. No. 8 925
926 생명과학회지 2019, Vol. 29. No. 8
Journal of Life Science 2019, Vol. 29. No. 8 927
928 생명과학회지 2019, Vol. 29. No. 8
Journal of Life Science 2019, Vol. 29. No. 8 929
930 생명과학회지 2019, Vol. 29. No. 8
Journal of Life Science 2019, Vol. 29. No. 8 931
932 생명과학회지 2019, Vol. 29. No. 8
Fig. 1. Effects of gut microbiota on the differentiation into brown adipocytes. In response to thermogenic stimuli, preadipocytes differentiated by bone morphogenetic proteins (BMPs) [118, 124] show browning and the following changes. Activation of brown preadipocytes and beiging of white adipocytes are promoted by transcription factors including Pparγ, Prdm16 (PR domain–containing 16), and Pgc1α (Pparγ-coactivator 1α), concomitantly with mitochondrial biogenesis genes represented by Ucp1. BAT: Brown adipose tissue; MSC: Mesenchymal stem cell.
a major active ingredient of Cordyceps militaris, attenuates an increase in fat mass and body weight and reverses the gut microbiome alteration induced by a high-fat diet [7].
Decreased ambient temperature, in concert with Ucp1 in- duction, changes gut microbiome composition to suppress diet-induced obesity [136]. Concretely speaking, the most well-pronounced microbiota alterations after such anti- obesity manipulations are downregulated Firmicutes and upregulated Bacteroidetes. On the other hand, type 2 dia- betes mellitus (T2DM) patients and mice with high-fat diet–
induced obesity have shown a lower proportion of Firmi- cutes and a higher relative abundance of Bacteroidetes in the gut microbiota, with additional upregulation of gut Proteobacteria in T2DM patients [7, 104]. Thus, the phylum ratio of Firmicutes to Bacteroidetes is deemed to be worth further studies for the promotion of energy expenditure and WAT browning. Still, the specific correlation between the intestinal microflora and reduced body weight gain were left for further studies.
Bacterial Metabolites That Are Expected to Promote Adipocyte Browning
The gut microbiota ferments indigestible dietary fiber and produces diverse metabolites, mostly consisting of short-
chain fatty acids (SCFAs) as final byproducts [72]. Most SCFAs found in human intestines are acetate (C2), propio- nate (C3), and butyrate (C4) [25]. SCFAs act as the signals from the microbiota, thereby affecting host fatty-acid, sugar, and cholesterol metabolic pathways; promoting fatty acid ca- tabolism; and suppressing fat accumulation in adipose tissue and gluconeogenesis in the liver [24, 31].
Kong et al. have found that high-calorie diets can alter the gut microbiota by making it less diverse and more en- riched with obesity-related genera, which partly contribute to the weight gain and metabolic disorders. Additionally, these diets can suppress SCFA-producing bacteria [62]. It is widely known that probiotics improve the gut microenviron- ment by neutralizing the high-calorie diet-induced proin- flammatory bacteria [33, 70]. Fecal fermentation products of dietary fiber derived from common beans (Phaseolus vulgaris) hinder the expression of some transcription factors critical for adipogenesis, PPARγ and C/EBPα, thereby hampering adipocytic differentiation in its early stage [69]. Nonetheless, the fermentation product increases the expression of other transcription factors, PPARδ and UCP2, which induce adipo- cyte energy expenditure [69]. Because various types of diet- ary fiber are foods for the hindgut microbiota, the composi- tion of consumed fiber affects the composition of the gut
Journal of Life Science 2019, Vol. 29. No. 8 933
Fig. 2. Mechanisms of known antiobesity interventions intended to modify human metabolism and centered on gut microbiota modulation. BCoAT: Butyryl-CoA: acetate coenzyme A-transferase; DIO2: Type 2 deiodinase; GPR43: G-protein-coupled re- ceptor 43; IL-6: Interleukin 6; LPS: Lipopolysaccharide; mmdA: Methylmalonyl-CoA decarboxylase; SCFA: Short chain fatty acid; TGR5 (GPBAR1): G protein-coupled bile acid receptor 1; TNF-α: Tumor necrosis factor-alpha.
microbiota, resulting in different types of SCFAs [69].
Cholic acids have been demonstrated to promote Ucp1- dependent thermogenesis and formation of multilocular lip- id droplets and to suppress diet-induced weight gain under thermoneutrality in mice [137]. The composition of secon- dary bile acids is modified differently, depending on the gut microbiota, and greatly affects the cholesterol level [54, 113].
The microbiota profile induced by a high-fat diet drastically raises serum levels of cholesterols and triglycerides, and the profile of serum parameters is normalized by simvastatin, a drug known to decrease low-density lipoprotein cholester- ol and prevent cardiovascular diseases [46]. The attenuation of the hypolipidemic effect of simvastatin by an induced gut microbiota alteration is believed to be related to the regu- lation of bile acid synthesis from cholesterol [46]. Neverthe- less, those authors could not ascertain whether this effect is primarily triggered by downregulation of gram-positive bacteria, upregulation of gram-negative bacteria, or a combi- nation.
Lifestyle Changes Inducing Adipose Browning and Microbiota Modulation
Caloric restriction where intake of necessary nutrients is guaranteed restrains body weight gain, recovers insulin sen- sitivity, and promotes browning of white fat depots, con-
currently with modification of the gut microbiota [36, 37].
The caloric-restriction-induced microbiota is believed to de- crease blood lipopolysaccharide levels by downregulating major enzymes through suppression of Kdo2-lipid A biosyn- thesis and the lipid A-Ara4N pathway, and to hinder bacte- rial lipopolysaccharide biosynthesis [36]. The alteration of the intestinal microenvironment, concurrent with the above properties, is deemed to be the chief cause of such metabolic alterations including improved glucose tolerance and insulin sensitivity.
It has been well established that physical exercise can also alter the human gut microbiota positively although the effect of physical activity on BAT activation is less remarkable.
Allen et al. have demonstrated that exercise leads to the in- duction of SCFA-regulating genes BCoAT (butyryl-CoA: ace- tate coenzyme A-transferase) and MmdA (methylmalonyl- CoA decarboxylase) and increases fecal SCFA concentration in lean people [4]. Exercise changes butyrate-regulating bac- terial groups, with upregulation of Roseburia spp., Lachnospira spp., Clostridiales spp., Faecalibacterium spp., and Lachnospi- raceae spp. and downregulation of Bacteroides spp. and Rikenella spp., especially in lean subjects [4]. These changes, however, are rolled back after a washout period returning to a sedentary lifestyle [4]. In rodent studies, exercise was found to induce subcutaneous WAT beiging, but the under-
934 생명과학회지 2019, Vol. 29. No. 8
lying mechanism and applicability to humans are not yet clear [32]. The conclusions of rodent and human studies have been inconsistent, but moderate physical activity tends to not raise BAT thermogenic activity.
Conclusion
In this review, we covered antiobesity interventions that normalize metabolism by manipulating the gut microbiota and promoting thermogenesis in adipose depots (Fig. 2).
Nevertheless, because the idea of inducing browning as a countermeasure against obesity and metabolic disorders came to the fore only recently, there is still a lot to be dis- covered about the underlying mechanisms. In addition, the attributes of different fat depots and the inter-relations among adipose tissues and other organs are important for the practical application of microbiota-based techniques to a clinical human antiobesity program and require further research. The effects of identical treatments can be different, or even opposite, depending on such factors as sex, the loca- tion of fat depots, and the BMI of the subjects [4, 135]. The causal relations must also be thoroughly examined: the pos- sibility that a common cause of a microbial change and of browning promotion exists—or the causality has been un- derstood reversely—should never be ignored [83]. Thus, ex- tensive preclinical and clinical trials are needed to determine proper ways to introduce specific phyla of the gut micro- biota into a human recipient without adverse effects or inter- ference by the patient’s existing normal microflora, and to guarantee the effectiveness of such treatments.
Acknowledgement
This research was supported by the research fund of Soonchunhyang University.
References
1. Agerholm-Larsen, L., Raben, A., Haulrik, N., Hansen, A. S., Manders, M. and Astrup, A. 2000. Effect of 8 week intake of probiotic milk products on risk factors for cardiovascular diseases. Eur. J. Clin. Nutr. 54, 288-297.
2. Alberdi, G., Rodríguez, V. M., Miranda, J., Macarulla, M.
T., Churruca, I. and Portillo, M. P. 2013. Thermogenesis is involved in the body-fat lowering effects of resveratrol in rats. Food Chem. 141, 1530-1535.
3. Alisi, A., Bedogni, G., Baviera, G., Giorgio, V., Porro, E., Paris, C., Giammaria, P., Reali, L., Anania, F. and Nobili,
V. 2014. Randomised clinical trial: The beneficial effects of vsl#3 in obese children with non-alcoholic steatohepatitis.
Aliment. Pharmacol. Ther. 39, 1276-1285.
4. Allen, J. M., Mailing, L. J., Niemiro, G. M., Moore, R., Cook, M. D., White, B. A., Holscher, H. D. and Woods, J. A. 2018.
Exercise alters gut microbiota composition and function in lean and obese humans. Med. Sci. Sports Exerc. 50, 747-757.
5. Aller, R., De Luis, D., Izaola, O., Conde, R., Gonzalez Sagrado, M., Primo, D., De La Fuente, B. and Gonzalez, J.
2011. Effect of a probiotic on liver aminotransferases in non- alcoholic fatty liver disease patients: A double blind randomized clinical trial. Eur. Rev. Med. Pharmacol. Sci. 15, 1090-1095.
6. An, H. M., Park, S. Y., Lee, D. K., Kim, J. R., Cha, M. K., Lee, S. W., Lim, H. T., Kim, K. J. and Ha, N. J. 2011.
Antiobesity and lipid-lowering effects of bifidobacterium spp. In high fat diet-induced obese rats. Lipids Health Dis.
10, 116.
7. An, Y., Li, Y., Wang, X., Chen, Z., Xu, H., Wu, L., Li, S., Wang, C., Luan, W., Wang, X., Liu, M., Tang, X. and Yu, L. 2018. Cordycepin reduces weight through regulating gut microbiota in high-fat diet-induced obese rats. Lipids Health Dis. 17, 276.
8. Anhê, F. F., Nachbar, R. T., Varin, T. V., Trottier, J., Dudonné, S., Le Barz, M., Feutry, P., Pilon, G., Barbier, O.
and Desjardins, Y. 2019. Treatment with camu camu (myrciaria dubia) prevents obesity by altering the gut micro- biota and increasing energy expenditure in diet-induced obese mice. Gut 68, 453-464.
9. Apovian, C. M., Aronne, L. J., Bessesen, D. H., McDonnell, M. E., Murad, M. H., Pagotto, U., Ryan, D. H., Still, C. D.
and Endocrine, S. 2015. Pharmacological management of obesity: An endocrine society clinical practice guideline. J.
Clin. Endocrinol. Metab. 100, 342-362.
10. Arias, N., Picó, C., Teresa Macarulla, M., Oliver, P., Miranda, J., Palou, A. and Portillo, M. P. 2017. A combination of re- sveratrol and quercetin induces browning in white adipose tissue of rats fed an obesogenic diet. Obesity 25, 111-121.
11. Askari, H., Rajani, S. F., Poorebrahim, M., Haghi-Aminjan, H., Raeis-Abdollahi, E. and Abdollahi, M. 2018. A glance at the therapeutic potential of irisin against diseases involv- ing inflammation, oxidative stress, and apoptosis: An in- troductory review. Pharmacol. Res. 129, 44-55.
12. Azhar, Y., Parmar, A., Miller, C. N., Samuels, J. S. and Raya- lam, S. 2016. Phytochemicals as novel agents for the in- duction of browning in white adipose tissue. Nutr. Metab.
(Lond.) 13, 89.
13. Bahler, L., Verberne, H. J., Admiraal, W. M., Stok, W. J., Soeters, M. R., Hoekstra, J. B. and Holleman, F. 2016. Differ- ences in sympathetic nervous stimulation of brown adipose tissue between the young and old, and the lean and obese.
J. Nucl. Med. 57, 372-377.
14. Bomhof, M. R., Saha, D. C., Reid, D. T., Paul, H. A. and Reimer, R. A. 2014. Combined effects of oligofructose and bifidobacterium animalis on gut microbiota and glycemia in obese rats. Obesity 22, 763-771.
Journal of Life Science 2019, Vol. 29. No. 8 935
15. Brahe, L. K., Le Chatelier, E., Prifti, E., Pons, N., Kennedy, S., Blædel, T., Håkansson, J., Dalsgaard, T. K., Hansen, T.
and Pedersen, O. 2015. Dietary modulation of the gut micro- biota–a randomised controlled trial in obese postmeno- pausal women. Br. J. Nutr. 114, 406-417.
16. Brooks, A. W., Priya, S., Blekhman, R. and Bordenstein, S.
R. 2018. Gut microbiota diversity across ethnicities in the united states. PLoS Biol. 16, e2006842.
17. Cani, P. D., Joly, E., Horsmans, Y. and Delzenne, N. M. 2006.
Oligofructose promotes satiety in healthy human: A pilot study. Eur. J. Clin. Nutr. 60, 567-572.
18. Cani, P. D., Lecourt, E., Dewulf, E. M., Sohet, F. M., Pachikian, B. D., Naslain, D., De Backer, F., Neyrinck, A.
M. and Delzenne, N. M. 2009. Gut microbiota fermentation of prebiotics increases satietogenic and incretin gut peptide production with consequences for appetite sensation and glucose response after a meal. Am. J. Clin. Nutr. 90, 1236- 1243.
19. Cao, L., Choi, E. Y., Liu, X., Martin, A., Wang, C., Xu, X.
and During, M. J. 2011. White to brown fat phenotypic switch induced by genetic and environmental activation of a hypothalamic-adipocyte axis. Cell Metab. 14, 324-338.
20. Cerdo, T., Garcia-Santos, J. A., M, G. B. and Campoy, C.
2019. The role of probiotics and prebiotics in the prevention and treatment of obesity. Nutrients 11, 635.
21. Chen, L. H., Chen, Y. H., Cheng, K. C., Chien, T. Y., Chan, C. H., Tsao, S. P. and Huang, H. Y. 2018. Antiobesity effect of lactobacillus reuteri 263 associated with energy metabo- lism remodeling of white adipose tissue in high-energy- di- et-fed rats. J. Nutr. Biochem. 54, 87-94.
22. Choque Delgado, G. T. and Tamashiro, W. 2018. Role of prebiotics in regulation of microbiota and prevention of obesity. Food Res. Int. 113, 183-188.
23. Chung, H. J., Jae, G. Y., Lee, I. A., Liu, M. J., Shen, Y. F., Sharma, S. P., Jamal, M. A., Yoo, J. H., Kim, H. J. and Hong, S. T. 2016. Intestinal removal of free fatty acids from hosts by lactobacilli for the treatment of obesity. FEBS Open Bio.
6, 64-76.
24. Clarke, G., Stilling, R. M., Kennedy, P. J., Stanton, C., Cryan, J. F. and Dinan, T. G. 2014. Minireview: Gut microbiota:
The neglected endocrine organ. Mol. Endocrinol. 28, 1221- 1238.
25. Cummings, J. H. 1981. Short chain fatty acids in the human colon. Gut 22, 763-779.
26. Cypess, A. M., Weiner, L. S., Roberts-Toler, C., Franquet Elia, E., Kessler, S. H., Kahn, P. A., English, J., Chatman, K., Trauger, S. A., Doria, A. and Kolodny, G. M. 2015.
Activation of human brown adipose tissue by a beta3-a- drenergic receptor agonist. Cell Metab. 21, 33-38.
27. Dardmeh, F., Alipour, H., Gazerani, P., van der Horst, G., Brandsborg, E. and Nielsen, H. I. 2017. Lactobacillus rham- nosus pb01 (dsm 14870) supplementation affects markers of sperm kinematic parameters in a diet-induced obesity mice model. PloS One 12, e0185964.
28. de Carvalho Marchesin, J., Celiberto, L. S., Orlando, A. B., de Medeiros, A. I., Pinto, R. A., Zuanon, J. A. S., Spolidorio,
L. C., dos Santos, A., Taranto, M. P. and Cavallini, D. C.
U. 2018. A soy-based probiotic drink modulates the micro- biota and reduces body weight gain in diet-induced obese mice. J. Funct. Foods 48, 302-313.
29. De Lorenzo, A., Costacurta, M., Merra, G., Gualtieri, P., Cioccoloni, G., Marchetti, M., Varvaras, D., Docimo, R. and Di Renzo, L. 2017. Can psychobiotics intake modulate psy- chological profile and body composition of women affected by normal weight obese syndrome and obesity? A double blind randomized clinical trial. J. Transl. Med. 15, 135.
30. Dehghan, P., Gargari, B. P., Jafar-Abadi, M. A. and Alias- gharzadeh, A. 2014. Inulin controls inflammation and meta- bolic endotoxemia in women with type 2 diabetes mellitus:
A randomized-controlled clinical trial. Int. J. Food Sci. Nutr.
65, 117-123.
31. den Besten, G., van Eunen, K., Groen, A. K., Venema, K., Reijngoud, D. J. and Bakker, B. M. 2013. The role of short- chain fatty acids in the interplay between diet, gut micro- biota, and host energy metabolism. J. Lipid Res. 54, 2325- 2340.
32. Dewal, R. S. and Stanford, K. I. 2019. Effects of exercise on brown and beige adipocytes. Biochim. Biophys. Acta Mol. Cell Biol. Lipids 1864, 71-78.
33. Djuric, Z. 2017. Obesity-associated cancer risk: The role of intestinal microbiota in the etiology of the host proinflam- matory state. Transl. Res. 179, 155-167.
34. Edrisi, F., Salehi, M., Ahmadi, A., Fararoei, M., Rusta, F.
and Mahmoodianfard, S. 2018. Effects of supplementation with rice husk powder and rice bran on inflammatory fac- tors in overweight and obese adults following an energy-re- stricted diet: A randomized controlled trial. Eur. J. Nutr. 57, 833-843.
35. Everard, A., Lazarevic, V., Derrien, M., Girard, M., Muccioli, G. G., Neyrinck, A. M., Possemiers, S., Van Holle, A., François, P. and de Vos, W. M. 2011. Responses of gut microbiota and glucose and lipid metabolism to prebiotics in genetic obese and diet-induced leptin-resistant mice. Diabetes 60, 2775-2786.
36. Fabbiano, S., Suarez-Zamorano, N., Chevalier, C., Lazarevic, V., Kieser, S., Rigo, D., Leo, S., Veyrat-Durebex, C., Gaia, N., Maresca, M., Merkler, D., Gomez de Aguero, M., Mac- pherson, A., Schrenzel, J. and Trajkovski, M. 2018. Function- al gut microbiota remodeling contributes to the caloric re- striction-induced metabolic improvements. Cell Metab. 28, 907-921.e907.
37. Fabbiano, S., Suarez-Zamorano, N., Rigo, D., Veyrat-Durebex, C., Stevanovic Dokic, A., Colin, D. J. and Trajkovski, M.
2016. Caloric restriction leads to browning of white adipose tissue through type 2 immune signaling. Cell Metab. 24, 434- 446.
38. Fak, F. and Backhed, F. 2012. Lactobacillus reuteri prevents diet-induced obesity, but not atherosclerosis, in a strain de- pendent fashion in apoe-/- mice. PLoS One 7, e46837.
39. Famouri, F., Shariat, Z., Hashemipour, M., Keikha, M. and Kelishadi, R. 2017. Effects of probiotics on nonalcoholic fatty liver disease in obese children and adolescents. J. Pediatr.
936 생명과학회지 2019, Vol. 29. No. 8
Gastroenterol. Nutr. 64, 413-417.
40. Fathi, Y., Faghih, S., Zibaeenezhad, M. J. and Tabatabaei, S. H. 2016. Kefir drink leads to a similar weight loss, com- pared with milk, in a dairy-rich non-energy-restricted diet in overweight or obese premenopausal women: A random- ized controlled trial. Eur. J. Nutr. 55, 295-304.
41. Fontane, L., Benaiges, D., Goday, A., Llaurado, G. and Pedro- Botet, J. 2018. Influence of the microbiota and probiotics in obesity. Clin. Investig. Arterioscler. 30, 271-279.
42. Genta, S., Cabrera, W., Habib, N., Pons, J., Carillo, I. M., Grau, A. and Sanchez, S. 2009. Yacon syrup: Beneficial ef- fects on obesity and insulin resistance in humans. Clin. Nutr.
28, 182-187.
43. Gobel, R. J., Larsen, N., Jakobsen, M., Molgaard, C. and Michaelsen, K. F. 2012. Probiotics to adolescents with obe- sity: Effects on inflammation and metabolic syndrome. J.
Pediatr. Gastroenterol. Nutr. 55, 673-678.
44. Gomes, A. C., de Sousa, R. G. M., Botelho, P. B., Gomes, T. L. N., Prada, P. O. and Mota, J. F. 2017. The additional effects of a probiotic mix on abdominal adiposity and anti- oxidant status: A double-blind, randomized trial. Obesity 25, 30-38.
45. Halkjaer, S. I., Nilas, L., Carlsen, E. M., Cortes, D., Halldórs- son, T. I., Olsen, S. F., Pedersen, A. E., Krogfelt, K. A. and Petersen, A. M. 2016. Effects of probiotics (vivomixx®) in obese pregnant women and their newborn: Study protocol for a randomized controlled trial. Trials 17, 491.
46. He, X., Zheng, N., He, J., Liu, C., Feng, J., Jia, W. and Li, H. 2017. Gut microbiota modulation attenuated the hypo- lipidemic effect of simvastatin in high-fat/cholesterol-diet fed mice. J. Proteome Res. 16, 1900-1910.
47. Heaton, J. M. 1972. The distribution of brown adipose tissue in the human. J. Anat. 112, 35-39.
48. Higashikawa, F., Noda, M., Awaya, T., Danshiitsoodol, N., Matoba, Y., Kumagai, T. and Sugiyama, M. 2016. Antiobesi- ty effect of pediococcus pentosaceus lp28 on overweight subjects: A randomized, double-blind, placebo-controlled clinical trial. Eur. J. Clin. Nutr. 70, 582-587.
49. Hill, J. O., Wyatt, H. R. and Peters, J. C. 2012. Energy balance and obesity. Circulation 126, 126-132.
50. Hume, M. P., Nicolucci, A. C. and Reimer, R. A. 2017.
Prebiotic supplementation improves appetite control in chil- dren with overweight and obesity: A randomized controlled trial. Am. J. Clin. Nutr. 105, 790-799.
51. Ikeda, K., Maretich, P. and Kajimura, S. 2018. The common and distinct features of brown and beige adipocytes. Trends Endocrinol. Metab. 29, 191-200.
52. Jung, S. P., Lee, K. M., Kang, J. H., Yun, S. I., Park, H. O., Moon, Y. and Kim, J. Y. 2013. Effect of lactobacillus gasseri bnr17 on overweight and obese adults: A randomized, dou- ble-blind clinical trial. Kor. J. Fam. Med. 34, 80.
53. Jung, S., Lee, Y. J., Kim, M., Kim, M., Kwak, J. H., Lee, J.
W., Ahn, Y. T., Sim, J. H. and Lee, J. H. 2015. Supplementa- tion with two probiotic strains, lactobacillus curvatus hy7601 and lactobacillus plantarum ky1032, reduced body adiposity and lp-pla2 activity in overweight subjects. J.
Funct. Foods 19, 744-752.
54. Kaddurah-Daouk, R., Baillie, R. A., Zhu, H., Zeng, Z. B., Wiest, M. M., Nguyen, U. T., Wojnoonski, K., Watkins, S.
M., Trupp, M. and Krauss, R. M. 2011. Enteric microbiome metabolites correlate with response to simvastatin treatment.
PLoS One 6, e25482.
55. Kadooka, Y., Sato, M., Imaizumi, K., Ogawa, A., Ikuyama, K., Akai, Y., Okano, M., Kagoshima, M. and Tsuchida, T.
2010. Regulation of abdominal adiposity by probiotics (lactobacillus gasseri sbt2055) in adults with obese tenden- cies in a randomized controlled trial. Eur. J. Clin. Nutr. 64, 636-643.
56. Kahan, S. 2016. Overweight and obesity management strat- egies. Am. J. Manag. Care 22, s186-196.
57. Karbaschian, Z., Mokhtari, Z., Pazouki, A., Kabir, A., Hedayati, M., Moghadam, S. S., Mirmiran, P. and Hekmat- doost, A. 2018. Probiotic supplementation in morbid obese patients undergoing one anastomosis gastric bypass-mini gastric bypass (oagb-mgb) surgery: A randomized, dou- ble-blind, placebo-controlled, clinical trial. Obes. Surg. 28, 2874-2885.
58. Kershaw, E. E. and Flier, J. S. 2004. Adipose tissue as an endocrine organ. J. Clin. Endocrinol. Metab. 89, 2548-2556.
59. Kim, J., Yun, J. M., Kim, M. K., Kwon, O. and Cho, B. 2018.
Lactobacillus gasseri bnr17 supplementation reduces the visceral fat accumulation and waist circumference in obese adults: A randomized, double-blind, placebo-controlled trial. J. Med. Food 21, 454-461.
60. Kim, M., Kim, M., Kang, M., Yoo, H. J., Kim, M. S., Ahn, Y. T., Sim, J. H., Jee, S. H. and Lee, J. H. 2017. Effects of weight loss using supplementation with lactobacillus strains on body fat and medium-chain acylcarnitines in overweight individuals. Food Funct. 8, 250-261.
61. Kondo, S., Xiao, J. Z., Satoh, T., Odamaki, T., Takahashi, S., Sugahara, H., Yaeshima, T., Iwatsuki, K., Kamei, A. and Abe, K. 2010. Antiobesity effects of bifidobacterium breve strain b-3 supplementation in a mouse model with high-fat diet-induced obesity. Biosci. Biotechnol. Biochem. 74, 1656-1661.
62. Kong, C., Gao, R., Yan, X., Huang, L. and Qin, H. 2018.
Probiotics improve gut microbiota dysbiosis in obese mice fed a high-fat or high-sucrose diet. Nutrition 60, 175-184 63. Larsen, N., Vogensen, F. K., Gobel, R. J., Michaelsen, K. F.,
Forssten, S. D., Lahtinen, S. J. and Jakobsen, M. 2013. Effect of lactobacillus salivarius ls-33 on fecal microbiota in obese adolescents. Clin. Nutr. 32, 935-940.
64. Leber, B., Tripolt, N. J., Blattl, D., Eder, M., Wascher, T. C., Pieber, T. R., Stauber, R., Sourij, H., Oettl, K. and Stadlbauer, V. 2012. The influence of probiotic supplementation on gut permeability in patients with metabolic syndrome: An open label, randomized pilot study. Eur. J. Clin. Nutr. 66, 1110- 1115.
65. Li, S., Li, J., Mao, G., Wu, T., Lin, D., Hu, Y., Ye, X., Tian, D., Chai, W., Linhardt, R. J. and Chen, S. 2019. Fucosylated chondroitin sulfate from isostichopus badionotus alleviates metabolic syndromes and gut microbiota dysbiosis induced by high-fat and high-fructose diet. Int. J. Biol. Macromol. 124,
Journal of Life Science 2019, Vol. 29. No. 8 937
377-388.
66. Li, X., Song, Y., Ma, X., Zhang, Y., Liu, X., Cheng, L., Han, D., Shi, Y., Sun, Q. and Yang, C. 2018. Lactobacillus planta- rum and lactobacillus fermentum alone or in combination regulate intestinal flora composition and systemic immunity to alleviate obesity syndrome in high-fat diet rat. Int. J. Food Sci. Technol. 53, 137-146.
67. Liao, A. H., Jiang, C. B., Li, C. C., Chuang, H. C., Chiang Chiau, J. S., Chan, W. T., Yeung, C. Y., Cheng, M. L. and Lee, H. C. 2017. Combining ultrasound and lactobacilli treat- ment for high-fat-diet-induced obesity in mice. J. Anim.
Physiol. Anim. Nutr. 101, 703-712.
68. Lindsay, K. L., Kennelly, M., Culliton, M., Smith, T., Maguire, O. C., Shanahan, F., Brennan, L. and McAuliffe, F. M. 2014.
Probiotics in obese pregnancy do not reduce maternal fast- ing glucose: A double-blind, placebo-controlled, random- ized trial (probiotics in pregnancy study). Am. J. Clin. Nutr.
99, 1432-1439.
69. Lu, H. Y., Zeng, H., Zhang, L., Porres, J. M. and Cheng, W. H. 2018. Fecal fermentation products of common bean-derived fiber inhibit c/ebpalpha and ppargamma ex- pression and lipid accumulation but stimulate ppardelta and ucp2 expression in the adipogenesis of 3t3-l1 cells. J.
Nutr. Biochem. 60, 9-15.
70. Lucas, C., Barnich, N. and Nguyen, H. T. T. 2017. Microbio- ta, inflammation and colorectal cancer. Int. J. Mol. Sci. 18, 1310.
71. Luoto, R., Kalliomaki, M., Laitinen, K. and Isolauri, E. 2010.
The impact of perinatal probiotic intervention on the devel- opment of overweight and obesity: Follow-up study from birth to 10 years. Int. J. Obes. (Lond.) 34, 1531-1537.
72. Macfarlane, G. T. and Macfarlane, S. 2012. Bacteria, colonic fermentation, and gastrointestinal health. J. AOAC Int. 95, 50-60.
73. Madjd, A., Taylor, M. A., Mousavi, N., Delavari, A., Malek- zadeh, R., Macdonald, I. A. and Farshchi, H. R. 2015.
Comparison of the effect of daily consumption of probiotic compared with low-fat conventional yogurt on weight loss in healthy obese women following an energy-restricted diet:
A randomized controlled trial. Am. J. Clin. Nutr. 103, 323- 329.
74. Mahadzir, M. D. A., Shyam, S., Barua, A., Krishnappa, P.
and Ramamurthy, S. 2017. Effect of probiotic microbial cell preparation (mcp) on fasting blood glucose, body weight, waist circumference, and faecal short chain fatty acids among overweight malaysian adults: A pilot randomised controlled trial of 4 weeks. Malays. J. Nutr. 23, 329-341.
75. Minami, J., Kondo, S., Yanagisawa, N., Odamaki, T., Xiao, J. Z., Abe, F., Nakajima, S., Hamamoto, Y., Saitoh, S. and Shimoda, T. 2015. Oral administration of bifidobacterium breve b-3 modifies metabolic functions in adults with obese tendencies in a randomised controlled trial. J. Nutr. Sci. 4, e17.
76. Minami, J., Iwabuchi, N., Tanaka, M., Yamauchi, K., Xiao, J. Z., Abe, F. and Sakane, N. 2018. Effects of bifidobacterium breve b-3 on body fat reductions in pre-obese adults: A
randomized, double-blind, placebo-controlled trial. Biosci.
Microbiota Food Health 37, 67-75.
77. Mischke, M., Arora, T., Tims, S., Engels, E., Sommer, N., van Limpt, K., Baars, A., Oozeer, R., Oosting, A. and Bäckhed, F. 2018. Specific synbiotics in early life protect against diet- induced obesity in adult mice. Diabetes Obes. Metab. 20, 1408- 1418.
78. Miyoshi, M., Ogawa, A., Higurashi, S. and Kadooka, Y.
2014. Anti-obesity effect of lactobacillus gasseri sbt2055 ac- companied by inhibition of pro-inflammatory gene ex- pression in the visceral adipose tissue in diet-induced obese mice. Eur. J. Nutr. 53, 599-606.
79. Mohammadi-Sartang, M., Bellissimo, N., de Zepetnek, J. T., Brett, N., Mazloomi, S., Fararouie, M., Bedeltavana, A., Famouri, M. and Mazloom, Z. 2018. The effect of daily for- tified yogurt consumption on weight loss in adults with metabolic syndrome: A 10-week randomized controlled trial. Nutr. Metab. Cardiovasc. Dis. 28, 565-574.
80. Mosqueda-Solís, A., Sánchez, J., Portillo, M. P., Palou, A.
and Picó, C. 2018. Combination of capsaicin and hesperidin reduces the effectiveness of each compound to decrease the adipocyte size and to induce browning features in adipose tissue of western diet fed rats. J. Agric. Food Chem. 66, 9679- 9689.
81. Murakami, Y., Ojima-Kato, T., Saburi, W., Mori, H., Matsui, H., Tanabe, S. and Suzuki, T. 2015. Supplemental epilactose prevents metabolic disorders through uncoupling protein-1 induction in the skeletal muscle of mice fed high-fat diets.
Br. J. Nutr. 114, 1774-1783.
82. Mykhal'chyshyn, H., Bodnar, P. and Kobyliak, N. 2013.
Effect of probiotics on proinflammatory cytokines level in patients with type 2 diabetes and nonalcoholic fatty liver disease. Lik. Sprava. 2, 56-62.
83. Nedergaard, J. and Cannon, B. 2014. The browning of white adipose tissue: Some burning issues. Cell Metab. 20, 396-407.
84. Neyrinck, A. M., Bindels, L. B., Geurts, L., Van Hul, M., Cani, P. D. and Delzenne, N. M. 2017. A polyphenolic ex- tract from green tea leaves activates fat browning in high-fat-diet-induced obese mice. J. Nutr. Biochem. 49, 15-21.
85. Nicolucci, A. C., Hume, M. P., Martinez, I., Mayengbam, S., Walter, J. and Reimer, R. A. 2017. Prebiotics reduce body fat and alter intestinal microbiota in children who are over- weight or with obesity. Gastroenterology 153, 711-722.
86. Nihei, N., Okamoto, H., Furune, T., Ikuta, N., Sasaki, K., Rimbach, G., Yoshikawa, Y. and Terao, K. 2018. Dietary α- cyclodextrin modifies gut microbiota and reduces fat accu- mulation in high-fat-diet-fed obese mice. BioFactors 44, 336- 347.
87. Ohyama, K., Nogusa, Y., Shinoda, K., Suzuki, K., Bannai, M.
and Kajimura, S. 2016. A synergistic antiobesity effect by a combination of capsinoids and cold temperature through promoting beige adipocyte biogenesis. Diabetes 65, 1410- 1423.
88. Omar, J. M., Chan, Y. M., Jones, M. L., Prakash, S. and Jones, P. J. H. 2013. Lactobacillus fermentum and lactobacillus amylovorus as probiotics alter body adiposity and gut mi
938 생명과학회지 2019, Vol. 29. No. 8
croflora in healthy persons. J. Funct. Foods 5, 116-123.
89. World Health Organization. Noncommunicable diseases country profiles 2018: World Health Organization, 2018.
90. World Health Organization. 2018. Obesity and overweight.
Retrieved from https://www.who.int/news-room/fact- sheets/ detail/obesity-and-overweight.
91. Osterberg, K. L., Boutagy, N. E., McMillan, R. P., Stevens, J. R., Frisard, M. I., Kavanaugh, J. W., Davy, B. M., Davy, K. P. and Hulver, M. W. 2015. Probiotic supplementation attenuates increases in body mass and fat mass during high-fat diet in healthy young adults. Obesity 23, 2364-2370.
92. Panteliou, E. and Miras, A. D. 2017. What is the role of bariatric surgery in the management of obesity? Climacteric 20, 97-102.
93. Park, D. Y., Ahn, Y. T., Park, S. H., Huh, C. S., Yoo, S.
R., Yu, R., Sung, M. K., McGregor, R. A. and Choi, M.
S. 2013. Supplementation of lactobacillus curvatus hy7601 and lactobacillus plantarum ky1032 in diet-induced obese mice is associated with gut microbial changes and reduc- tion in obesity. PLoS One 8, e59470.
94. Park, J. E., Oh, S. H. and Cha, Y. S. 2014. Lactobacillus plantarum lg42 isolated from gajami sik-hae decreases body and fat pad weights in diet-induced obese mice. J.
Appl. Microbiol. 116, 145-156.
95. Parnell, J. A. and Reimer, R. A. 2010. Effect of prebiotic fibre supplementation on hepatic gene expression and se- rum lipids: A dose–response study in jcr: La-cp rats. Br.
J. Nutr. 103, 1577-1584.
96. Parnell, J. A. and Reimer, R. A. 2009. Weight loss during oligofructose supplementation is associated with decreased ghrelin and increased peptide yy in overweight and obese adults. Am. J. Clin. Nutr. 89, 1751-1759.
97. Pedret, A., Valls, R. M., Calderón-Pérez, L., Llauradó, E., Companys, J., Pla-Pagà, L., Moragas, A., Martín-Luján, F., Ortega, Y. and Giralt, M. 2018. Effects of daily consumption of the probiotic bifidobacterium animalis subsp. Lactis cect 8145 on anthropometric adiposity biomarkers in abdomi- nally obese subjects: A randomized controlled trial. Int. J.
Obes. doi: 10.1038/s41366-018-0220-0.
98. Pyra, K. A., Saha, D. C. and Reimer, R. A. 2012. Prebiotic fiber increases hepatic acetyl coa carboxylase phosphor- ylation and suppresses glucose-dependent insulinotropic polypeptide secretion more effectively when used with metformin in obese rats. J. Nutr. 142, 213-220.
99. Rajkumar, H., Mahmood, N., Kumar, M., Varikuti, S. R., Challa, H. R. and Myakala, S. P. 2014. Effect of probiotic (vsl# 3) and omega-3 on lipid profile, insulin sensitivity, inflammatory markers, and gut colonization in overweight adults: A randomized, controlled trial. Mediators Inflamm.
2014, 348959.
100. Reimer, R. A., Willis, H. J., Tunnicliffe, J. M., Park, H., Madsen, K. L. and Soto-Vaca, A. 2017. Inulin-type fructans and whey protein both modulate appetite but only fructans alter gut microbiota in adults with overweight/obesity: A randomized controlled trial. Mol. Nutr. Food Res. 61, 1700484.
101. Respondek, F., Gerard, P., Bossis, M., Boschat, L., Bruneau, A., Rabot, S., Wagner, A. and Martin, J. C. 2013. Short-chain fructo-oligosaccharides modulate intestinal microbiota and metabolic parameters of humanized gnotobiotic diet in- duced obesity mice. PLoS One 8, e71026.
102. Reynes, B., Palou, M., Rodriguez, A. M. and Palou, A. 2018.
Regulation of adaptive thermogenesis and browning by prebiotics and postbiotics. Front. Physiol. 9, 1908.
103. Russo, F., Linsalata, M., Clemente, C., Chiloiro, M., Orlando, A., Marconi, E., Chimienti, G. and Riezzo, G. 2012. Inulin- enriched pasta improves intestinal permeability and modi- fies the circulating levels of zonulin and glucagon-like pep- tide 2 in healthy young volunteers. Nutr. Res. 32, 940-946.
104. Saez-Lara, M. J., Robles-Sanchez, C., Ruiz-Ojeda, F. J., Plaza-Diaz, J. and Gil, A. 2016. Effects of probiotics and synbiotics on obesity, insulin resistance syndrome, type 2 diabetes and non-alcoholic fatty liver disease: A review of human clinical trials. Int. J. Mol. Sci. 17, 928.
105. Sanchez, M., Darimont, C., Panahi, S., Drapeau, V., Marette, A., Taylor, V., Doré, J. and Tremblay, A. 2017. Effects of a diet-based weight-reducing program with probiotic sup- plementation on satiety efficiency, eating behaviour traits, and psychosocial behaviours in obese individuals. Nutrients 9, 284.
106. Sanchis-Chordà, J., del Pulgar, E. M. G., Carrasco-Luna, J., Benítez-Páez, A., Sanz, Y. and Codoñer-Franch, P. 2018.
Bifidobacterium pseudocatenulatum cect 7765 supple- mentation improves inflammatory status in insulin-re- sistant obese children. Eur. J. Nutr. 1-12.
107. Serrano, A., Asnani-Kishnani, M., Rodríguez, A. M., Palou, A., Ribot, J. and Bonet, M. L. 2018. Programming of the beige phenotype in white adipose tissue of adult mice by mild resveratrol and nicotinamide riboside supplementa- tions in early postnatal life. Mol. Nutr. Food Res. 62, 1800463.
108. Sharafedtinov, K. K., Plotnikova, O. A., Alexeeva, R. I., Sentsova, T. B., Songisepp, E., Stsepetova, J., Smidt, I. and Mikelsaar, M. 2013. Hypocaloric diet supplemented with probiotic cheese improves body mass index and blood pressure indices of obese hypertensive patients-a random- ized double-blind placebo-controlled pilot study. Nutrition 12, 138.
109. Shin, J. H., Nam, M. H., Lee, H., Lee, J. S., Kim, H., Chung, M. J. and Seo, J. G. 2018. Amelioration of obesity-related characteristics by a probiotic formulation in a high-fat di- et-induced obese rat model. Eur. J. Nutr. 57, 2081-2090.
110. Silvester, A. J., Aseer, K. R. and Yun, J. W. 2019. Dietary polyphenols and their roles in fat browning. J. Nutr.
Biochem. 64, 1-12.
111. Sivamaruthi, B. S., Kesika, P., Suganthy, N. and Chaiyasut, C. 2019. A review on role of microbiome in obesity and antiobesity properties of probiotic supplements. Biomed Res. Int. 2019, 3291367.
112. Skulachev, V. P. 1998. Uncoupling: New approaches to an old problem of bioenergetics. Biochim. Biophys. Acta Bioenerg.
1363, 100-124.
113. Staley, C., Weingarden, A. R., Khoruts, A. and Sadowsky,
Journal of Life Science 2019, Vol. 29. No. 8 939
M. J. 2017. Interaction of gut microbiota with bile acid me- tabolism and its influence on disease states. Appl. Microbiol.
Biotechnol. 101, 47-64.
114. Stenman, L. K., Lehtinen, M. J., Meland, N., Christensen, J. E., Yeung, N., Saarinen, M. T., Courtney, M., Burcelin, R., Lähdeaho, M. L. and Linros, J. 2016. Probiotic with or without fiber controls body fat mass, associated with se- rum zonulin, in overweight and obese adults-randomized controlled trial. EBioMedicine 13, 190-200.
115. Stojanovic, O., Kieser, S. and Trajkovski, M. 2018. Common traits between the beige fat-inducing stimuli. Curr. Opin.
Cell Biol. 55, 67-73.
116. Szulińska, M., Łoniewski, I., van Hemert, S., Sobieska, M.
and Bogdański, P. 2018. Dose-dependent effects of multi- species probiotic supplementation on the lipopolysaccharide (lps) level and cardiometabolic profile in obese post- menopausal women: A 12-week randomized clinical trial.
Nutrients 10, 773.
117. Thyagarajan, B. and Foster, M. T. 2017. Beiging of white adipose tissue as a therapeutic strategy for weight loss in humans. Horm. Mol. Biol. Clin. Investig. 31, doi: 10.1515/
hmbci-2017-0016.
118. Tseng, Y. H., Kokkotou, E., Schulz, T. J., Huang, T. L., Winnay, J. N., Taniguchi, C. M., Tran, T. T., Suzuki, R., Espinoza, D. O., Yamamoto, Y., Ahrens, M. J., Dudley, A.
T., Norris, A. W., Kulkarni, R. N. and Kahn, C. R. 2008.
New role of bone morphogenetic protein 7 in brown adipo- genesis and energy expenditure. Nature 454, 1000-1004.
119. Vajro, P., Mandato, C., Licenziati, M. R., Franzese, A., Vitale, D. F., Lenta, S., Caropreso, M., Vallone, G. and Meli, R.
2011. Effects of lactobacillus rhamnosus strain gg in pedia- tric obesity-related liver disease. J. Pediatr. Gastroenterol.
Nutr. 52, 740-743.
120. van Marken Lichtenbelt, W. D., Vanhommerig, J. W., Smulders, N. M., Drossaerts, J. M., Kemerink, G. J., Bouvy, N. D., Schrauwen, P. and Teule, G. J. 2009. Cold-activated brown adipose tissue in healthy men. N. Engl. J. Med. 360, 1500-1508.
121. Verbrugghe, A., Hesta, M., Gommeren, K., Daminet, S., Wuyts, B., Buyse, J. and Janssens, G. P. 2009. Oligofructose and inulin modulate glucose and amino acid metabolism through propionate production in normal-weight and obese cats. Br. J. Nutr. 102, 694-702.
122. Verhoef, S. P., Meyer, D. and Westerterp, K. R. 2011. Effects of oligofructose on appetite profile, glucagon-like peptide 1 and peptide yy3-36 concentrations and energy intake. Br.
J. Nutr. 106, 1757-1762.
123. Virtanen, K. A., Lidell, M. E., Orava, J., Heglind, M., Westergren, R., Niemi, T., Taittonen, M., Laine, J., Savisto, N. J., Enerback, S. and Nuutila, P. 2009. Functional brown adipose tissue in healthy adults. N. Engl. J. Med. 360, 1518- 1525.
124. Wang, E. A., Israel, D. I., Kelly, S. and Luxenberg, D. P.
1993. Bone morphogenetic protein-2 causes commitment and differentiation in c3h10t1/2 and 3t3 cells. Growth Factors 9, 57-71.
125. Wang, Z. B., Xin, S. S., Ding, L. N., Ding, W. Y., Hou, Y.
L., Liu, C. Q. and Zhang, X. D. 2019. The potential role of probiotics in controlling overweight/obesity and asso- ciated metabolic parameters in adults: A systematic review and meta-analysis. Evid. Based Complement. Alternat. Med.
2019, 3862971.
126. Weiner, J., Hankir, M., Heiker, J. T., Fenske, W. and Krause, K. 2017. Thyroid hormones and browning of adipose tissue.
Mol. Cell. Endocrinol. 458, 156-159.
127. Weitkunat, K., Stuhlmann, C., Postel, A., Rumberger, S., Fankhänel, M., Woting, A., Petzke, K. J., Gohlke, S., Schulz, T. J. and Blaut, M. 2017. Short-chain fatty acids and inulin, but not guar gum, prevent diet-induced obesity and insulin resistance through differential mechanisms in mice. Sci.
Rep. 7, 6109.
128. Whelan, K., Efthymiou, L., Judd, P. A., Preedy, V. R. and Taylor, M. A. 2006. Appetite during consumption of enteral formula as a sole source of nutrition: The effect of supple- menting pea-fibre and fructo-oligosaccharides. Br. J. Nutr.
96, 350-356.
129. Woodard, G. A., Encarnacion, B., Downey, J. R., Peraza, J., Chong, K., Hernandez-Boussard, T. and Morton, J. M.
2009. Probiotics improve outcomes after roux-en-y gastric bypass surgery: A prospective randomized trial. J.
Gastrointest. Surg. 13, 1198-1204.
130. Xue, B., Rim, J. S., Hogan, J. C., Coulter, A. A., Koza, R.
A. and Kozak, L. P. 2007. Genetic variability affects the development of brown adipocytes in white fat but not in interscapular brown fat. J. Lipid Res. 48, 41-51.
131. Yin, Y. N., Yu, Q. F., Fu, N., Liu, X. W. and Lu, F. G.
2010. Effects of four bifidobacteria on obesity in high-fat diet induced rats. World J. Gastroenterol. 16, 3394-3401.
132. You, Y., Han, X., Guo, J., Guo, Y., Yin, M., Liu, G., Huang, W. and Zhan, J. 2018. Cyanidin-3-glucoside attenuates high-fat and high-fructose diet-induced obesity by promot- ing the thermogenic capacity of brown adipose tissue. J.
Funct. Foods 41, 62-71.
133. You, Y., Yuan, X., Liu, X., Liang, C., Meng, M., Huang, Y., Han, X., Guo, J., Guo, Y. and Ren, C. 2017. Cyanidin-3-glu- coside increases whole body energy metabolism by upre- gulating brown adipose tissue mitochondrial function. Mol.
Nutr. Food Res. 61, 1700261.
134. Zarrati, M., Salehi, E., Nourijelyani, K., Mofid, V., Zadeh, M. J., Najafi, F., Ghaflati, Z., Bidad, K., Chamari, M., Kari- mi, M. and Shidfar, F. 2014. Effects of probiotic yogurt on fat distribution and gene expression of proinflammatory factors in peripheral blood mononuclear cells in over- weight and obese people with or without weight-loss diet.
J. Am. Coll. Nutr. 33, 417-425.
135. Zhuang, P., Shou, Q., Lu, Y., Wang, G., Qiu, J., Wang, J., He, L., Chen, J., Jiao, J. and Zhang, Y. 2017. Arachidonic acid sex-dependently affects obesity through linking gut microbiota-driven inflammation to hypothalamus-adipose- liver axis. Biochim. Biophys. Acta Mol. Basis Dis. 1863, 2715- 2726.
136. Zietak, M., Kovatcheva-Datchary, P., Markiewicz, L. H.,
940 생명과학회지 2019, Vol. 29. No. 8
초록:장내 미생물의 조절과 지방세포의 갈색지방화를 통한 비만 조절 연구
조예진1․라만 엠디 샤밈2,3․김용식1,2,3*
(1순천향대학교 의과대학 의학과, 2순천향대학교 의과대학 미생물학교실, 3순천향대학교 의과대학 조직재생연구소)
비만은 에너지 섭취와 소비의 불균형으로 인해 유발되는 비정상적인 지방 축적으로, 근래에 다양한 만성질환을 초래하는 주요 국제 보건 문제로 부상하였다. 이러한 이유로, 비만 문제에 대한 여러 해결책들이 제시되고 있다.
에너지를 저장하며 내분비 작용을 하는 백색 지방과 달리 열을 생성하여 에너지를 발산하는 두 종류의 지방조직 인 갈색 지방과 베이지색 지방이 성인에 존재하며 외부 자극에 의해 유도될 수 있다는 것이 밝혀진 이래로, 이들 은 비만 치료의 유망한 표적으로서 각광받고 있다. 이러한 외부 자극 중, 인간 장관계에서 인간과 공존하는 장내 미생물총은 다양한 대사 작용에 참여하며, 이를 조절하는 것이 여러 대사 질환의 치료에 유력한 작용을 할 것으로 보인다. 따라서, 다양한 연구에서 항비만 치료가 장내 미생물 환경 전환이나 갈색 지방 조직 활성화에 미치는 영 향에 초점을 맞추고 있다. 본 총설에서는 비만과 체중 증가, 대사 질환을 해소하는 것으로 알려진 자극과, 장내 미생물총의 변화나 갈색지방의 활성화를 야기하는 자극과, 이 자극들과 장내 미생물총의 조작, 지방조직의 갈색화 사이에서 알려져 있거나 있을 수 있는 상관관계를 중점적으로 다루고자 한다.
Stahlman, M., Kozak, L. P. and Backhed, F. 2016. Altered microbiota contributes to reduced diet-induced obesity upon cold exposure. Cell Metab. 23, 1216-1223.
137. Zietak, M. and Kozak, L. P. 2016. Bile acids induce un-
coupling protein 1-dependent thermogenesis and stimulate energy expenditure at thermoneutrality in mice. Am. J.
Physiol. Endocrinol. Metab. 310, E346-354.