산업용 효모 Hybrid의 선별을 위한 우성선별표지로서의 Aureobasidin A 내성유전자의 이용
전한택†·박은미·김근*
수원대학교 생명공학과
Received: April 6, 2011 / Revised: April 21, 2011 / Accepted: April 22, 2011
The Use of Aureobasidin A Resistant Gene as the Dominant Selectable Marker for the Selection of Industrial Yeast Hybrid. Jeon, Hantaek†, Uhnmee Park, and Keun Kim*. Department of Bioscience and Biotechnology, The University of Suwon, Hwaseong 445-743, Korea − For the strain improvement of the indus- trial polyploid yeast strain through hybridization and protoplast fusion, a dominant selection marker other than a recessive marker such as the auxotrophic marker was required for the selection of the resulting hybrids. In the present investigation, the aureobasidin A resistant gene was tested in relation to whether it can be used as the dominant selectable marker for the isolation of hybrids of the yeast Saccharomyces. The plasmid pAUR112, car- rying the gene responsible for resistance to aureobasidin A, was introduced into the haploid yeast strain K114/
YIp. From the rare-mating between polyploid C6 and haploid K114/YIp carrying pAUR112, many hybrids were obtained from the agar medium containing 0.5 µg/ml of aureobasidin A. The hybrids exhibited characteristics derived from both of the parental strains; and the cell sizes of the hybrids were larger than those of the parental strains. These results showed that the aureobasidin A resistant gene could be successfully used as the dominant selectable marker for the isolation of yeast hybrids resulting from rare-mating.
Key words: Dominant selectable marker, aureobasidin A resistancy, hybrid, rare-mating
서 론
효모균주의 발효적 특성을 개량하기 위해서는 여러 가지 유전적 방법이 사용되고 있으며, 여기에는 선택(selection), 돌 연변이, 교배(hybridization), 원형질체융합(protoplast fusion), 형질전환(transformation) 등을 들 수 있다[19]. 이중에서 특 히 교배, 원형질체융합, 형질전환의 방법을 통해서 발효특성 이 증진된 균주의 선별을 위해서는 반드시 선별표지 (selectable marker)의 도입이 필요하다.
효모들은 본질적으로 포자를 만들어 교배하는 능력이 있 다. 반수체 효모의 경우는 에이(a)타입과 알파(α)타입이 쉽 게 mating하여 이배체(diploid)가 형성하는데 이 이배체는 원 래 두 반수체 효모의 좋은 특성을 동시에 나타내어 균주 개 량이 이루어진다.
Rare-mating은 정상적인 에이타입 반수체와 알파타입 반 수체의 자연적인 교배방법이 아니고, 동일한 mating-type
균주들 간의 hybrid형성이나 반수체와 배수체(polyploid)의 hybrid형성이다. Non-mating type 두 균주의 세포를 높은 농도로 혼합하면 아주 낮은 비율로 hybrid형성이 되는 경 우가 드물게 관찰 되는데, 이것을 rare-mating이라고 부른 다[17].
포자형성 또는 교배가 안 되는 효모들끼리 재조합을 시킬 때는 원형질체를 만든 후 융합하는 방법을 사용한다[19]. 여 기에서는 두 균주의 세포벽을 제거하여 원형질체를 만들고 원형질막을 융합시켜 두 게놈을 하나의 막 안으로 도입 시 킨 후 융합체를 선별하는 것이다. 여기에서는 양쪽 게놈이 모두 각자 선별 표지 유전자를 소유함으로써 단독으로는 선 별배지에서 성장할 수 없고, 융합체만이 선별배지에서 콜로 니를 형성하게 하는 방법을 사용한다.
효모균주 개발에서 일반적으로 도입된 선별 유전자 표 지로는 호흡 결여성 돌연변이와 영양요구성 돌연변이가 있 다. 호흡결여성 돌연변이[18]는 cytochrome a, b가 결핍되 거나 미토콘드리아 DNA의 일부 또는 전부가 파괴되어 있 어서 glycerol 같은 발효가 되지 않는 탄소원을 유일 탄소 원으로 함유하는 배지에서는 성장이 되지 않는다. 이렇게 선별된 호흡결여성 돌연변이는 열성으로 분류되는 표지이 다. 한편 영양요구성 돌연변이도 열성 표지인데, 산업용 효 모 균주는 대부분 이배체 이상의 배수체이기 때문에 강력
*Corresponding author
Tel: +82-31-220-2344, Fax: +82-31-220-2344 E-mail: [email protected]
†Present address: Aujeon Korea, 1720-38, C-4 Taejang 2-dong, Wonju 220-962, Korea
한 돌연변이원을 처리했을 때만 영양요구성 돌연변이체를 얻게 된다[16]. 그 결과 돌연변이체의 DNA는 많은 손상 을 입기가 십상이어서 산업용 효모 균주는 우수성을 상실 하거나 건강하지 못한 균주로 변화되는 위험을 감수하게 된다.
이를 대신하여 영양요구성 돌연변이 주나 호흡결여성 돌 연변이 주에 항생제 내성 같은 우성 표지를 도입할 수 있다 면, 상대편 산업균주에 열성인 표지를 도입시키지 않아도 hybrid만을 선별할 수 있게 된다. 즉, 영양요구성 모균주가 필요로 하는 양분을 제외시킨 최소배지를 사용함으로써 이 모균주의 성장을 제어하고, 또한 그 배지가 항생제도 포함 하고 있다면 야생형 산업균주의 성장도 제어하여 hybrid만 이 콜로니를 생성하는 조건을 만족시키기 때문이다. 그동안 산업균주에 돌연변이원을 처리하지 않고 그대로 교배나 원 형질체 융합에 사용할 수 있게 하는 방안이 절실히 모색 되 어 왔는데, 우성 표지의 도입은 이 조건을 만족시키는 시도 라고 할 수 있다. 박테리아에서 흔히 쓰이는 amphicillin이나 tetracycline 같은 항생제는 효모에서는 작용하지 않으므로 우 리는 aureobasidin 내성 유전자를 주목하게 되었다.
Aureobasidium pullulans No. R106이 만들어내는 항진 균 항생물질인aureobasidin A는 8개의 L-form amino acids와 1개의 hydroxy acid가 ring 구조를 이루고 있다[10, 20]. Aureobasidin A는 Candida albicans, Saccharomyces cerevisiae, Schizosaccharomyces pombe, Aspergillus 일부에 대해서 강력한 항진균 활성을 보인다.
최근에 밝혀진 바로는 aureobasidin은 inositol phosphoryl ceramide(IPC) synthase를 방해하여, 효모의 원형질막의 주 성분 중 하나로 쓰이는 sphingolipid류의 합성을 방해하는 것 으로 알려졌다[5]. 동물세포에서 발견되는 sphingolipid들은 기본적인 ceramide 구조에 phosphocholine 등을 첨부하여 사용하는데 반해 효모는 phosphoinositol류에 ceramide가 붙 은 것들을 주로 사용한다[1]. Aureobasidin에 의해 저해되는 스텝은 phytoceramide를 IPC(inositolphosphoryl ceramide) 로 전환시키는 스텝이다. 이 효소 즉, inositolphosphoryl ceramide synthese의 저해는 IPC이외에도 mannosyl-IPC, inisitolphosphoryl MIPC들의 생성을 차단하는 것으로 보고 되었다. 효모에서 sphingolipid들은 원형질막의 중요한 구성 성분으로 쓰여질 뿐 아니라, 동물 세포의 경우와 마찬가지 로 신호전달이나 cell-cycle에도 관련된 필수적 역할을 한다 고 생각된다. 최근에는 효모에서도 endocytosis, proteolysis 뿐 아니라[5] inositol phosphorylceramide synthase가 cera- mide의 농도를 조절하여 G1에서 S로 가는 단계를 조정하는 protein kinase에게 영향을 미친다는 보고가 있었다[3, 12].
이 효소의 작용이 결핍되었을 때는 이런 역할이 저해되는 동 시에 phosphoceramide가 축적되어 세포를 해롭게 하기 때문 에 aureobasidin은 효모의 생존을 절대적으로 위협하는 항진 균물질로 작용한다는 것이다[2].
실제로 aureobasidin A을 처리하고 2~3시간 경과 시 많은 세포에서 actin assembly가 와해되어지고 정상적인 budding process가 억제되며 5~6시간이 지나면서 세포가 죽기 시작할 때 아미노산이 세포에서 이탈되고 세포막이 파괴되어 지는 것이 관찰되었다[7]. 이러한 항진균 항생물질인 aureobasidin A에 대한 내성유전자 AUR1-C[6, 8, 9]가 발견되었는데 이 내성 유전자가 만드는 변이 단백질은 정상인 단백질과 비교 했을 때 158번째 아미노산이 phe에서 tyr으로 변해 있음이 발견되었다. 정상 단백질(AUR1)은 aureobasidin에 의해 기능 이 상실되는 반면 변이 단백질은 항생제의 공격을 피할 수 있 는 것으로 보인다. AUR1-C 유전자의 발현으로 aureobasidin A에 대해서 특이적인 내성을 갖게 되고 또한 이 유전자는 우성으로 작용하기 때문에 세포내에 1 copy만 존재해도 내 성형질을 나타낸다[9].
본 연구에서는 호흡결여성 돌연변이나 영양요구성 돌연변 이와 같은 선별표지가 없는 산업용 배수체 효모 균주에 교 배나 원형질체 융합에 의해 새로운 형질을 도입하고자 할 때 우성 표지인 AUR1-C가 hybrid를 선별하기 위한 표지로서 유용하게 사용될 수 있는지를 알아보고자 하였다.
재료 및 방법 효모균주와 플라스미드
본 연구에 사용한 효모 균주는 C6, KL, K114YIp인데 그 특성과 source는 Table 1에 나타내었다. C6와 KL(발연1호) 는 배수체로서 각각 에탄올 발효능과 에탄올 생존능이 우수 한 균주이다. K114YIp는 반수체로서 glucoamylase gene (STA)를 가지고 있는 S. diastaticus의 염색체에 α-amylase 유전자를 도입시킨 형질전환체이다[13].
KL 및 K114YIp의 형질전환에 사용한 플라스미드는 Takara Shuzo Co., Ltd.(Japan)에서 구입한 pAUR112 (Code No. 3601)이다(Fig. 1). pAUR112는 S. cerevisiae로 부터 유래된 ARS1/CEN4 유전자를 보유하며 또한 pBR322 로부터 유래된 ColE1 ori 유전자를 갖는 효모 자율복제형 대 장균·효모 shuttle vector[17]이다. 또한 대장균 세포내에서 Table 1. Yeast strains used and their source.
Yeast strain Characteristics Source C6 Saccharomyces sp.
polyploid Laboratory collection KL Saccharomyces sp. Distillery
yeast, polyploid Distillery
K114YIpMS
a trp1 ade6 his2 STAa Sac- charomyces diastaticus hap- loid transformant secreting both α-amylase and glu- coamylase
Laboratory collection (Kim and Kim, 1996)
a STA, gene of glucoamylase.
의 선별표지로써 ampicillin 내성유전자가 있고 S. cerevisiae 의 선별표지로써 AUR1-C[9] 유전자와 URA3[15] 유전자를 보유하고 있다. 효모세포에 pAUR112의 도입은 Ito et al.[11] 방법을 사용하였다.
배지 및 배양
효모의 성장배지에는 1%(w/v) yeast extract(Y), 2%(w/v) peptone(P), 2%(w/v) dextrose(D)로 구성된 YPD 배지를 사 용하였다. 선별배지로써 사용되는 SG와 SD에는 영양요구주 의 성장에 필요한 탄소원과 질소원 그리고 항생제인 aureobasidin A를 넣어서 사용하였다. SG배지는 0.67%(w/v) Difco yeast nitrogen base(without amino acids), 3%(w/v) glycerol로 되어 있고, SD배지는 0.67%(w/v) Difco yeast nitrogen base(without amino acids), 2%(w/v) dextrose로 되어있다. 원형질체 융합 시 재생배지로는 삼투안정제로서 1.2 M KCl이 포함된 SG를 사용하였다. 세포의 amylase 분 비에 의한 전분 분해능을 측정하기 위한 배지는 YPD1S3(1%
dextrose + 3% soluble potato starch)에 접종한 균을 2일간 배양하였다. 또한 rare-mating에 의해 선별된 균주의 포자 형 성능을 조사하기 위한 배지인 McClary's media는 1%(w/v) potassium acetate, 0.25%(w/v) yeast extract, 0.1%(w/v) glucose로 되어있다. 모든 agar 배지의 제조 시에는 2%(w/
v) agar를 사용하였다. 효모 배양온도는 30oC이었으며, 산소 존재 하에 균체를 액체 배양할 때는 rotary shaking incu- bator에서 200 rpm 속도로 배양하였다.
Rare-mating
Rare-mating은 crossing plate method[17]를 이용하였다.
활성화된 두 균주 를 백금니로 YPD agar plate배지에 서로 교차되게 접종한 후 30oC에서 2일간 배양하였다. 서로 교차 되어지는 지점에서 자라는 균체들을 serial dilution 한 후 선 별배지에 접종하여 성장하는 균주를 선별하였다. 선별배지 는 aureobasidin A(0.5 µg/mL)가 첨가된 SD배지로서 aureobasidin A에 내성이 있고 영양요구성 돌연변이가 아닌 세포만 성장할 수 있다.
Hybrid의 특성 조사
효모의 전분분해능 측정을 위해서 효모균체를 toothpick를 사용하여 YPD1S3에 접종하고 2일간 배양한 후 다시 2일간 4oC에서 냉장시킨 후 균체 주변에 생긴 투명환의 지름(mm) 을 측정하였다.
그 외 hybrid의 특성 조사는 Kreger-van Rij(1984)[14]의 방법을 사용하였다. 탄소원 이용 여부 조사를 위해 2일간 활 성화시킨 균주를 3 mL의 멸균수에 넣고 잘 섞은 다음 현탁 액 100 µL를 3 mL 멸균수로 다시 희석한 후 100 µL를 취 하여 0.5%(w/v) raffinose를 함유한 5 mL yeast nitrogen base 배지에 배양 후 측정하였다. Bacto-yeast nitrogen base 배지는 6.7 g의 yeast nitrogen base와 5 g의 raffinose를 100 mL의 증류수에 녹여 filter(pore size 0.2 µm)로 여과한 다음 0.5 mL을 취하여 4.5 mL의 멸균수에 희석하여 제조 하였다. Sorbitol, starch, mannitol, galactose, maltose, glucose 등도 위와 같은 방법으로 준비 하였다.
Cycloheximide 내성 조사는 탄소원 이용과 같은 방법으로 시료를 준비한 후 0.67%(w/v) yeast nitrogen base와 10%
(w/v) glucose, 그리고 0.1 g의 cycloheximide가 2.5 mL acetone에 녹아있는 것을 첨가하여 100 mL stock solution을 만들고, 그중 0.5 mL를 취하여 4.5 mL의 멸균수에 희석하 여 사용하였다.
세포형태 관찰
전자현미경(SEM, Scanning electron microscopy) 및 광 학현미경 관찰을 위하여 3 mL YPD배지에 one colony를 접 종하고 30oC에서 200 rpm으로 18시간 동안 배양한 세포를 1/10, 1/100, 1/1000로 희석하였다. 1/100로 희석한 시료에서 10 µL를 취해 광학현미경(Olympus BX60, Japan)에서 1,000배로 관찰하고, 촬영모드에서 노출기(Olympus PM-20 exposure control unit)의 시간을 0.2초로 하여 사진을 찍었 다. 그리고 전자현미경(Cambridge Sci. Instruments, Ltd., Cambridge, England) 사진을 찍기 위해 동일한 시료를 전처 리하였다. 위에서 1/100으로 희석한 시료 1 mL를 polycar- bonate membrane(지름 13 mm, pore size 0.1 µm)이 부착 된 여과기로 여과하였다. Polycarbonate membrane은 여과기 내부에 부착시켰고, 윗부분에는 주사기를 끼워서 사용하였 Fig. 1. Genetic and restriction enzyme map of pAUR112 DNA.
AUR1-C, aureobasidin A resistant gene from S. cerevisiae. URA3, uracil selective marker from S. cerevisiae. ARS1, replication origin from S. cerevisiae. CEN4, centromere from S. cerevisiae. Amp, selective marker from E. coli. Ori, replication origin from E. coli.
다. 주사기에 2 mL의 1차 고정액을 넣고 polycarbonate membrane이 찢어지지 않도록 주의하면서 배출시켰다. 1차 고정액은 25%(w/v) glucoaldehyde 용액을 phosphate buffer (pH7.2)로 희석하여 4%(w/v)로 만들어서 사용하였다. 다시 1 mL의 1차 고정액을 주사기에 담고 실린더를 살며시 눌러 한 방울 정도 떨어뜨린 후 1시간 동안 반응시킨 뒤 모두 배 출했다. 그리고 잔량의 1차 고정액을 세척하기 위해 2 mL 의 증류수를 주사기에 넣어 밀어내고 다시 1 mL의 증류수 를 담아서 15분씩 두 번 방치한 후 1시간이 지나면 증류수 를 모두 배출하고, 2 mL의 2차 고정액을 넣어 1시간 동안 반응시켰다. 2차 고정액은 4%(w/w) osmium tetroxide 용액 (Sigma. O-0631)을 deionized water에 1%(w/w)가 되도록 희석하여 사용하고 갈색병에 넣어 4oC에서 보관하였다. 남 아있는 2차 고정액을 모두 배출하고 3 mL의 증류수로 세척 한 후 다시 1 mL의 증류수를 담아서 15분간 방치했다. 주 사기에 남아있는 증류수를 모두 배출하고 여과기의 polycarbonate membrane을 조심스럽게 분리해서 미리 준비 한 25%(w/v) 에탄올 1 mL가 들어있는 5 mL용량(높이 5 cm)의 cap tube에 넣었다. 그리고 50%, 75%, 90%, 100%
(w/v)의 에탄올을 각각 15분씩 차례대로 갈아주면서 반응시 켰다. 100%(w/v) 에탄올 처리가 끝나면 에탄올을 isoamyl acetate로 치환해주기 위해 에탄올 : isoamylacetate의 비율이 75 : 25, 50 : 50, 25 : 75, 0 : 100인 용액을 1 mL씩 차례대로 교체하면서 각각 15분씩 반응시켰다. 100%(w/v)의 isoamyl acetate 처리가 끝나면 isoamyl acetate를 액화 CO2로 치환 해주기 위해 polycarbonate membrane을 조심스럽게 꺼내서 critical point dryer(BIO-RAD E3000 Series) 안에 넣고 액 화 CO2를 주입하였다. Critical point dryer는 액화 CO2를 주입하기 전에 10oC 정도로 유지시켰다. 액화 CO2의 수면 을 최상부로 유지하면서 아래층의 isoamyl acetate를 완전히 배출하고 1시간 동안 반응시켰다. 치환된 액화 CO2를 기화 시키기 위해 38oC로 가온한 후 완전히 마른 polycarbonate membrane을 semicoating system(FISONS Instrument)을 이 용하여 5분 동안 gold coating을 하고 전자현미경으로 관찰 후 6,000배에서 SEM 사진을 찍었다.
결과 및 고찰
선별배지에서 aureobasidin A의 최적 선별 농도 결정 선별배지에서 aureobasidin A의 최적 선별농도를 알아보 기 위해, 0.2 µg/mL부터 0.5 µg/mL의 여러 농도의 aureo- basidin을 첨가시킨 여러 가지 종류의 배지에서 성장한 콜로 니 수를 조사하였고, 그 결과를 Table 2에 나타내었다. KL 과 C6의 내성을 비교해볼 때 C6가 KL보다 더 높은 aureobasidin 내성을 보였다. 두 균주의 야생형균주의 경우 glucose가 탄소원인 SD와 YPD 배지에서는 aureobasidin 0.2-0.3 µg/mL 농도에서 생존 콜로니 수가 급격히 감소하
였고, 0.5 µg/mL 농도에서도 약간의 수가 생존하였다. 따 라서 세포 완전 사멸농도는 0.5 µg/mL 이상으로 판단된다 . 반면에 KL 호흡결여주는 glucose가 탄소원인 SD배지에 서는 야생형균주에 비해 aureobasidin A 내성이 높아져서 0.5 µg/mL 농도에서부터 생존수가 뚜렷하게 감소하였다.
KL 균주보다 KL의 호흡 결여주는 aureobasidin에 대한 내 성이 좀 더 강하였는데, 0.3 µg/mL에서는 셀 수 없을 정 도로 많은(TMTC) 콜로니가 자랐으며 보통 다른 실험실에 서 추천하는 농도인 0.5 µg/mL에서도 1,000개 이상의 콜 로니가 자랐다. 한편 glycerol이 탄소원인 SG와 YPG에서 는 더 민감하여 KL 경우는 0.2-0.3 µg/mL에서 완전 사멸 하였고, C6 경우는 0.5 µg/mL에서 완전 사멸하였다. 따라 서 KL과 C6 야생형균주를 대상으로 하는 선별배지의 aureobasidin A의 적정농도는 SD와 YPD에서 0.5 µg/mL 이상, 그리고 SG와 YPG에서는 0.5 µg/mL인 것을 알 수 있었다.
Aureobasidin marker의 transformation
배수체 균주 C6에 전분분해능을 도입시키기 위하여 전분 분해능이 있는 반수체 영양요구주인 K114YIp균주와 rare- mating[19]을 시도하였다. 이를 위하여 K114YIp균주에 AUR1-C(aureobasidin A 항생제에 대한 내성 유전자)를 포 Table 2. Growth of yeast strains in various concentrations of aureobasidin.
Yeast strain
Aureobasidin conc.(µg/mL)
The numbera.b of yeast colony grown on different media.
SD SG YPD YPG
KL(Wild)
0.0 TMTCc TMTC Cd C
0.2 872 4 308 0
0.3 136 0 24 0
0.4 64 0 16 0
0.5 64 0 52 0
KL(RDe)
0.0 C 4 C 8
0.2 TMTC 0 TMTC 0
0.3 TMTC 0 192 0
0.4 TMTC 0 196 0
0.5 1204 0 16 0
C6
0.0 TMTC C C C
0.2 1200 640 TMTC 1700
0.3 368 252 136 88
0.4 192 64 124 0
0.5 136 0 44 0
aCell suspension of 0.1 mL containing 107 cells was spread onto various agar media and colony number was scored after 3 days of incubation.
bThe data shown are the average of the results of two experiments.
cTMTC, too many to count.
dC, confluent growth.
eRD, respiratory deficient mutant.
함하는 플라스미드 pAUR112를 도입 하였다.
Aureobasidin을 포함하는 YPD 배지에서 총 67주의 형질 전환체 K114YIp/pAUR112를 얻었다. 이들 중 rare-mating 에 사용할 균주로 전분 분해능을 나타내는 투명환 크기가 상 대적으로 큰 K114YIp/pAUR112-20을 선택하였다. 이 균주 로부터 플라스미드를 추출하여 전기영동을 실시한 결과 단 1개의 약 7 kb 정도의 밴드가 관찰이 되어 이 플라스미드가 pAUR112임을 확인할 수 있었다.
영양요구성과 aureobasidin 표지를 이용한 선별로 얻은 rare-mating hybrid
이제 영양요구주인 K114YIp에 aureobasidin 저항성을 가지는 플라스미드가 도입되었으므로, 이 균주와 산업균주 C6와의 hybrid를 얻는 시도를 하였다. Aureobasidin을 포 함하는 최소영양배지에서 성장하는 균주를 선별하면, C6 모균주는 aureobasidin 때문에 성장하지 못하고, K114YIp 모균주는 영양요구성 때문에 성장하지 못하므로, 오직 K114YIp의 pAUR112를 가지며 K114YIP의 영양요구성을 보완해주는 C6의 유전자를 소유하게 된 hybrid만이 성장 하게 된다.
이 K114YIp/pAUR112-20균주와 C6 간의 rare-mating을 하였고, 선별배지는 0.5 µg/mL의 aureobasidin을 첨가한 SG 배지를 사용하였다. Cross plating method[17]에 의한 교배 결과 많은 수의 hybrid를 얻었다. 이들 중 콜로니 성장과 투 명환 크기가 상대적으로 우수한 hybrid20-2, hybrid20-19의 두 균주를 선택하여 이들의 세포 형태와 생리적 특성을 조사 하였다. 이들 hybrid20-2과 hybrid20-19는 mannitol에서 약간 의 성장 정도 차이는 보였으나 모든 면에서 두 hybrid는 서 로 형질이 같았다. Hybrid들이 모균주 K114YIp/pAUR112- 20과는 같고 모균주 C6와는 다른 성질은 hybrid들이 전분함 유 고체배지에서 투명환을 보여 전분분해능을 가지고 있다 는 점이다. 이것은 원래 K114YIp의 전분분해능을 산업균주 C6에 도입 하려던 애초의 목적이 성취되었음을 보여주는 결
과이다. 이 밖에도 hybrid가 K114YIp/pAUR112-20와 같은 형질은 aureobasidin에 내성을 보이는 점, 그리고 mannitol 에서 성장을 보인 점들이었다. 한편 K114YIp/pAUR112-20 와는 다르고 C6와 같은 성질은 hybrid가 McClary의 배지에 서 많은 양의 포자 형성을 보인 점과 sorbitol에서는 좋은 성 장을 보인 점이다.
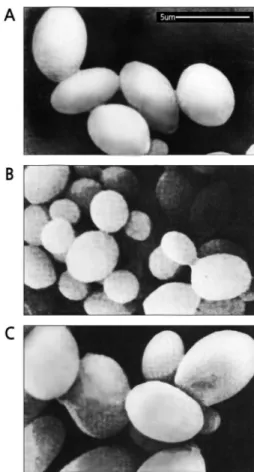
또한 두 모균주와 hybrid의 세포형태를 광학현미경으로 관
Fig 2. Scanning electron photomicrographs of C6 and K114YIp/pAUR112-20 and their hybrid. (A) C6 (B) K114YIp/
pAUR112-20 (C) Hybrid 20-2.
Table 3. Physiological characteristics of yeast C6 and K114YIp transformants harboring pAUR112 and their hybrids.
Strains Growth
in AURa
Size (mm) of halob
Cycloheximide
resistancy Spore formationc
Assimilation of carbon sourced
0.01% 0.1% Glu Mal Gal Man Sta Raf Sor
C6 -e 0.0 + + + +++ +++ +++ - ± +++ ±
K114YI p/pAUR112-20 + 3.5 + + - ++ ++ ± + ++ ++ -
Hybrid 20-2 + 2.0 + + ++ +++ ++ +++ + + +++ +
Hybrid 2-19 + 2.0 + + ++ +++ +++ +++ ± ++ +++ +
aAureobasidin A (0.5 µg/mL) were added to a SD agar plate.
bDiameter of clear zone (halo) formed around colony on YPD1S3.
cOne loopful activated cells were inoculated into a 50 mL cap tube containing 20 mL YPD broth at 30oC. After incubation for 2 days, 0.2 mL culture broth was transferred into 2 mL McClary's media and incubated at 23oC in a rotary shaking incubator for 7 days.
dGlu, glucose; Mal, maltose; Gal, galactose; Man, mannitol; Sta, starch; Raf, raffinose; Sor, sorbitol.
e+++, ++ represents good growth, + normal, ± poor, - no growth.
찰하였는데 hybrid의 세포는 모균주 K114YIp/pAUR112-20 보다 확실히 더 큰 크기였고, C6 보다는 더 동그란 형태의 약간 더 큰 세포 크기이었으며 세포 내부 형태도 다른 느낌 을 주었다(data not shown).
한편 전자현미경으로 관찰한 경우도 C6와 K114YIp/
pAUR112-20의 hybrid는 양친주 C6, K114YIp/pAUR112- 20보다 더 큰 세포 크기를 보였다(Fig. 2).
또한 에탄올 발효 후 생존능력이 좋은 배수체 산업균주 KL에게도 K114YIp의 전분분해 능력을 가지게 할 수 있는 지 시도하였다. KL과 K114YIp/pAUR112-4의 mating 후 aureobasidin 함유 SD 배지에서 hybrid를 선별하였다. 많은 hybrid들 중에서 전분분해능이 우수한 hybrid 4-13과 hybrid 4-14를 선택하여 여러 형질을 조사하였고, 그 결과를 Table 4에 나타내었다. Hybrid 들은 YPD1S3배지에서 전분분해를 나타내는 투명환을 형성하여 전분 분해 능력이 확실히 도입 되었음을 알 수 있었다. 이들 hybrid 들은 aureobasidin에 내 성, 전분분해능, cycloheximide 내성을 갖는 점과 포자를 형 성하지 않는 점에서 K114YIp/pAUR112-4에 더 가까운 형 질을 보였다. 그러나 이들 hybrid들은 SD배지에서 성장할 수 있기 때문에 이점은 영양요구주인 K114YIp/pAUR112-4 의 특성이 아니고, KL의 특성이기 때문에 두 모균주의 hybrid임을 알 수 있다. 한편 두 hybrid가 두 모균주를 닮지 않은 점은 maltose와 galactose의 동화능이 모두 감소하였다 는 점이다.
광학현미경 관찰에서 hybrid 4-13은 세포 크기가 큰 모균 주 KL과 크기가 작은 K114YIp/pAUR112-20의 중간 세포 크기였으며, KL 보다는 약간 더 동그란 형태였으며 세포 내 부는 K114YIp/pAUR112-20과 비슷한 느낌을 주었다(data not shown).
이상의 결과로 볼 때 영양요구성과 aureobasidin을 이용한 선별은 rare-mating에서 효과가 좋다는 결론을 내릴 수 있었 다. Aureobasidin 표지는 염색체 안에 있지 않고 플라스미드 상에 있었기 때문에 C6나 KL균주가 aureobasidin 표지가 있
는 플라스미드를 원형질막 안으로 받아들이는데 그치고, C6 나 KL균주의 염색체는 본연의 것을 그대로 간직 하여도 aureobasidin 저항성을 얻을 수 있었다. 그러나 hybrid의 여 러 생리적 특성과 현미경적 세포 형태로 판단하여 볼 때 일 단 mating이 된 후 C6 또는 KL 세포의 염색체와 K114YIp 염색체가 가지는 유전자들의 성격을 서로 재조합 한 것이 탄 생한 것으로 판단된다.
호흡결여성과 aureobasidin내성에 의한 선별
이번에는 호흡 결여성과 aureobasidin을 선별 표지로 사 용해 보기로 하였다. 우선 KL 균주에 aureobasidin을 포함 하는 플라스미드를 도입하여 형질전환시켰다. 이렇게 얻은 균주들은 1.6 µg/mL aureobasidin 함유배지에서도 성장하 는 aureobasidin내성을 보였다. 이 형질전환 균주들 중에서 발효력이 좋은 두 균주 KL/pAUR112(T4, T38)을 선택하 였다.
이 균주로부터 호흡 결여균주를 얻어서 융합이나 rare- mating에 사용해 보기로 하였다. Acroflavin으로 처리하여 호 흡결여 균주를 얻었다. 선별된 형질전환체의 호흡 결여주들 은 aureobasidin 0.5 µg/mL에서 잘 성장할 수 있었으므로 돌연변이원인 acroflavin이 aureobasidin 저항성 유전자에 아 무 손상을 주지 않았다는 것을 알 수 있었다. 또한 이들의 에탄올 발효 능력은 KL의 능력보다 오히려 조금 좋은 것들 이 발견 될 정도로 영향을 받지 않았다.
이렇게 얻은 KL-T4-RD12와 KL-T38-RD80을 C6균주와 원형질체 실험에 사용하였다. 산업현장 균주이며 다배체인 C6에 비해 KL은 에탄올 발효 후 생존능이 우수한데 비해 에탄올 발효능은 떨어진다. 따라서 C6 균주의 고발효능과 KL 균주의 고생존능을 겸하게 되는 균주를 제작하기위해 이 두 균주의 융합체를 만들고자 원형질체 융합을 시도하였다.
그런데 보통의 경우 1주일 안에 융합체가 자라나는데 반해 3주를 기다렸지만 융합체를 얻지 못했다. 혹시 KL의 원형질 체 형성률이 낮거나, 혹은 aureobasidin A가 세포벽 형성을
Table 4. Physiological characteristics of yeast KL and K114YIp transformants harboring pAUR112 and their hybrids.
Strains Growth
in AURa
Size (mm) of halob
Cycloheximide
resistancy Spore formationc
Assimilation of carbon sourced
0.01% 0.1% Glu Mal Gal Man Sta Raf Sor
KL -e 0.0 - - ± +++ +++ ++ - - ++ -
K114YI p/pAUR112-4 + 3.0 + + - ++ ++ + + ++ ++ -
Hybrid 4-13 + 3.5 + + - ++ + ± + ++ - -
Hybrid 4-14 + 3.0 + + - + + ± ± + ++ -
aAureobasidin A (0.5 µg/mL) were added to a SD agar plate.
bDiameter of clear zone (halo) formed around colony on a YPD1S3 agar plate.
cOne loopful activated cells were inoculated into a 50 mL cap tube containing 20 mL YPD broth at 30oC. After incubation for 2 days, 0.2 mL culture broth was transferred into a 2 mL McClary's medium and incubated at 23oC in a rotary shaking incubator for 7 days.
dGlu, glucose; Mal, maltose; Gal, galactose; Man, mannitol; Sta, starch; Raf, raffinose; Sor, sorbitol.
e+++, ++ represents good growth, + normal, ± poor, - no growth.
방해할 가능성 때문에 원형질체의 세포벽 형성이 되지 않아 융합주를 얻지 못하였는가 하고 생각하여, 원형질체를 만들 지 않고 KL/pAUR112 호흡결여주와 반수체 균주의 rare- mating도 시도하였으나 역시 실패하였다. 우리는 합당한 이 유를 찾을 수 없었다.
Aureobasidin A를 이용한 원형질체융합에서 융합주를 얻 지 못하는 이유는 확실히 알 수 없지만, Table 2에 나타난 바와 같이 야생형균주에 비해 호흡결여돌연변이주의 aureobasidin A의 내성이 증가한 것이 어떤 영향이 있지 않 나 의심하였다. 그러나 실질적으로 연관 관계가 있는지 밝 힐 수는 없었다. 어쨌든 aureobasidin A 내성을 선택표지로 사용하는 경우에는 호흡결여 표지를 counter selectable marker로 사용하는 것은 피하는 것이 좋다고 판단하였다.
요 약
교배와 원형질체 융합을 통한 배수체인 야생형 산업균주 의 개발을 위하여, hybrid의 선별표지로서 우성의 선별표지 인 aureobasidin A 내성이 사용될 수 있는 지를 알아보고자 하였다. 선별배지에서 aureobasidin A의 최적농도는 야생형 균주인 경우 SD와 YPD 배지에서는 0.5 µg/mL 이상이었고, SG와 YPG에서는 0.2-0.3 µg/mL 이었다. 한편 호흡결여돌 연변이주는 야생형 균주보다 훨씬 높은 농도의 aureobasidin A에도 내성이 있음을 나타내었다. 우리는 K114/YIP균주의 전분분해 능력이 배수체 야생형 산업 균주에 전달 될 수 있 는지를 이 방법을 통하여 관찰하였다. 반수체 영양요구성 균 주 K114/YIP에 aureobasidin에 대한 내성을 부여하는 pAUR112가 도입된 균주와 야생형 균주 KL 혹은 C6와의 rare-mating 후 aureobasidin A 함유 배지에서 성장한 hybrid 를 분리할 수 있었다. Hybrid는 전분분해 능력을 함유하고 있었을 뿐 아니라 두 양친의 특성을 동시에 지녔으며, 전자 현미경 관찰 결과에서도 hybrid는 양친주의 특성을 모두 갖 는 것으로 나타났다.
Acknowledgment
This work was supported by the New & Renewable Energy Program of the Korea Insitute of Energy Tech- nology Evaluation and Planning (KETEP) grant funded by the Ministry of Knowledge Economy, Republic of Korea (No. 20103040090060).
REFERENCES
1. Aeed, P. A., L. Y. Casey, M. M. Nagiec, and A. P.
Elhammer. 2009. Inhibition of inositol phosphorylceramide synthase by the cyclic peptide aureobasidin. Antimicrob
Agents Chemother. 53: 496-504.
2. Cerantola, V., I. Guillas, C. Roubalty, C. Vionet, D. Uldry, J. Krundsen, and A. Conzelmann. 2009. Aureobasidin A arrests growth of yeast cells through both ceramide in- toxication and deprivation of essential inisitolphosphoryl- ceramides. Molecular Microbiology 71: 1523-1537.
3. Cerbón, J., A. Falcon, A., C. Hernández-Luna, and D. Segura- Cobos. 2005. Inositol phosphoceramide synthase is a regulator of intracellular levels of diacylglycerol and ceramide during the G1 to S transition in Saccharomyces cerevisiae. Biochem J. 15; 388: 169-176.
4. Cook, J. R., S. L. Emanuel, and S. Pestka. 1993. Yeast artificial chromosome fragmentation vectors that utilize URA3 selection. Genet. Anal. Tech. Appl. 10: 109-112.
5. Cowart, L. A. and L. M. Obeid. 2007. Yeast sphingolipids:
recent developments in understanding biosynthesis, regula- tion, and function. Biochim Biophys Acta. 1771: 421-431.
6. Dujon, B., D. Alexandraki, B. Andre, and W. Ansorge.
1994. Complete DNA sequence of yeast chromosome XI.
Nature 369: 371-378.
7. Endo, M., K. Takesako, I. Kato, and H. Yamaguchi. 1997.
Fungicidal action of aureobasidin A, a cyclic depsipeptide antifungal antibiotic, against Saccharomyces cerevisiae.
Antimicrob. Agents. Chemother. 41: 672-676.
8. Hashida-Okado, T., A. Ogawa, R. Yasumoto, K. Takesako, and I. Kato. 1996. AUR1, a novel gene conferring aureo- basidin resistance on Saccharomyces cerevisiae : a study of defective morphologies in Aur1p-depleted cells. Mol. Gen.
Genet. 251: 236-244.
9. Heidler, S. A. and J. A. Radding. 1995. The AUR1 gene in Saccharomyces cerevisiae encodes dominant resistance to antifungal agent aureobasidin A (LY295337). Antimicrob.
Agents Chemother. 39: 2765-2769.
10. Ikai, K., K. Takesako, K. Shiomi, M. Moriguchi, Y. Umeda, J. Yamamoto, I. Kato, and H. Naganawa. 1991. Structure of aureobasidin A. J. Antibiot. 44: 925-933.
11. Ito, M., K. Murata, and A. Kimura. 1984. Transformation of intact yeast cells treated with alkali cations or thiol compounds. Agric. Biol. Chem. 48: 341-347.
12. Jesch, S. A., M. L. Gaspar, C. J. Stefan, M. A. Aregullin, and S. A. Henry. 2010. Interruption of inositol sphingolipid synthesis triggers Stt4p-dependent protein kinase C signal- ing. J. Biol. Chem. 285: 41947-41960.
13. Kim, T. and K. Kim. 1996. The construction of a stable starch-fermenting yeast strain using genetic engineering and rare-mating. Appli. Biochemistry and Biotechnology. 59: 39- 51.
14. Kreger-van Rij, N. J. W. 1984. The yeasts a taxonomic study, Third revised and enlarged edition, Elsevier Scince Publishers B. V.
15. Rose, M., P. Grisafi, and D. Botstein. 1984. Structure and function of the yeast URA3 gene : expression in Escherichia coli. Gene. 29: 113-124.
16. Saigal, D. 1993. Review article : Yeast strain development for ethanol production. Ind. J. Microbiol. 33: 159-168.
17. Sherman, F., G. Fink, and J. B. Hicks. 1986. Methods in Yeast Genetics, Laboratory Course Manual, Cold Spring Harbor Laboratory, Cold Spring Harbor, New York.
18. Spencer, J. F. T., C. Bizeau, N. Reynolds, and D. M. Spencer.
1985. The use of mitochondrial mutant in hybridization of industrial yeast strains. Curr. Genet. 9: 649-652.
19. Spencer, J. F. T. and D. M. Spencer. 1983. Genetic improve- ment of industrial yeasts. Ann. Rev. Microbiol. 37: 121-142.
20. Takesako, K., H. Kuroda, T. Inoue, F. Haruna, Y. Yoshikawa, I. Kato, K. Uchida, T. Hiratani, and H. Yamaguchi. 1993.
Biological properties of aureobasidin A, a cyclic depsipeptide antifungal antibiotic. J. Antibiot. 46: 1414-1427.